
Micro-RNA in Cholangiocarcinoma: Implications for Diagnosis, Prognosis, and Therapy

 and
and
Abstract
:1. Introduction
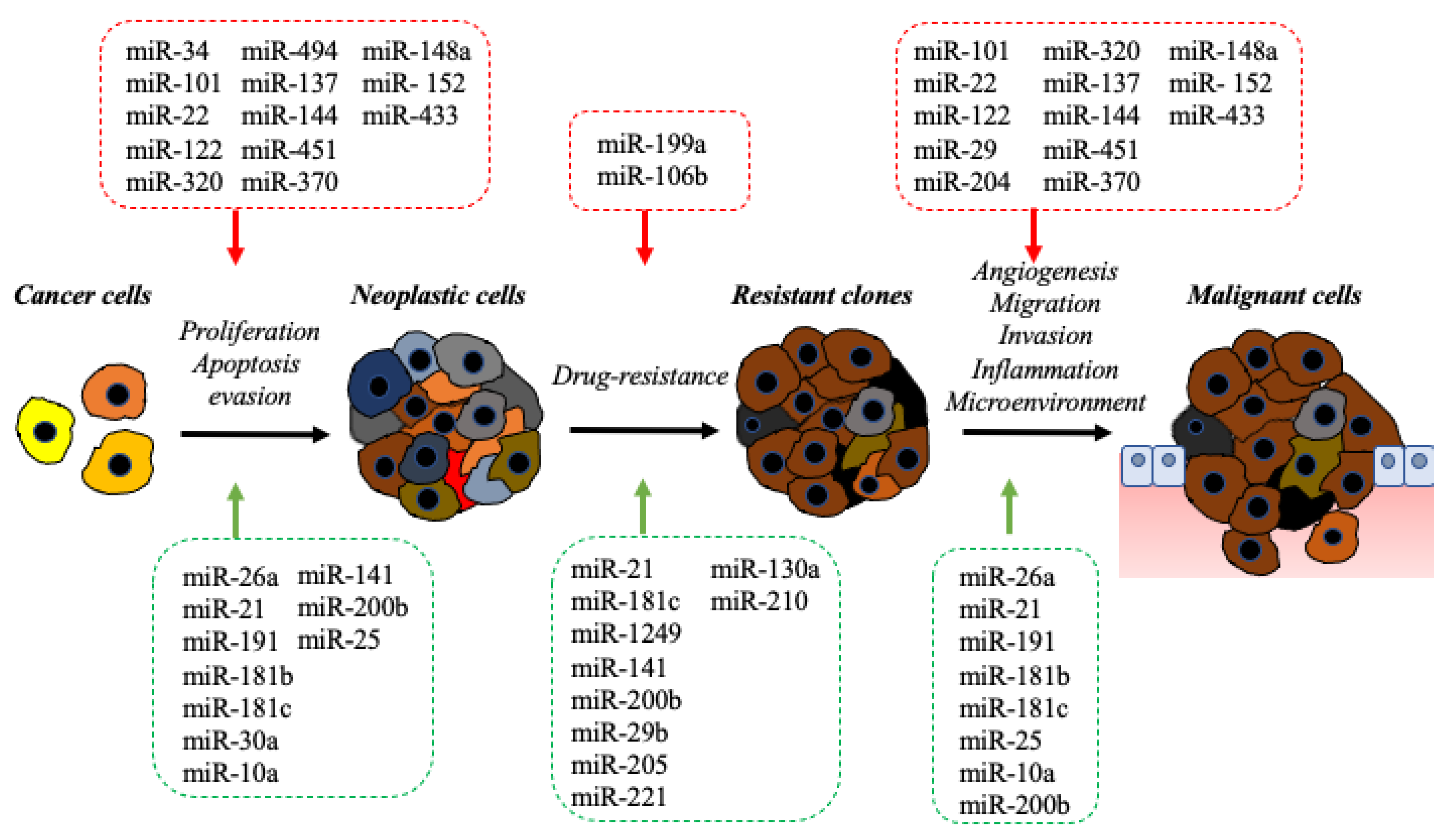
2. The Impact of the microRNA in Molecular Pathology of BDC
2.1. Onco-miRNAs
2.2. Oncosuppressor-miRNAs
3. The Impact of miRNAs in Precision Medicine of BDC
3.1. miRNAs as Biomarkers for BDC
3.2. Implication of MiRNAs in Therapy Sensitivity and Resistance of BDC
4. The Impact of miRNA-Based Therapeutic in BDC
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rizvi, S.; Khan, S.A.; Hallemeier, C.L.; Kelley, R.K.; Gores, G.J. Cholangiocarcinoma—Evolving concepts and therapeutic strate-gies. Nat. Rev. Clin. Oncol. 2018, 15, 95–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.A.; Tavolari, S.; Brandi, G. Cholangiocarcinoma: Epidemiology and risk factors. Liver Int. 2019, 39, 19–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertuccio, P.; Malvezzi, M.; Carioli, G.; Hashim, D.; Boffetta, P.; El-Serag, H.B.; Negri, E. Global trends in mortality from intrahepatic and extrahepatic cholangiocarcinoma. J. Hepatol. 2019, 71, 104–114. [Google Scholar] [CrossRef] [PubMed]
- Kelley, R.K.; Bridgewater, J.; Gores, G.J.; Zhu, A.X. Systemic therapies for intrahepatic cholangiocarcinoma. J. Hepatol. 2020, 72, 353–363. [Google Scholar] [CrossRef] [Green Version]
- Braconi, C.; Roessler, S.; Kruk, B.; Lammert, F.; Krawczyk, M.; Andersen, J.B. Molecular perturbations in cholangiocarcinoma: Is it time for precision medicine? Liver Int. 2019, 39, 32–42. [Google Scholar] [CrossRef] [Green Version]
- Adeva, J.; Sangro, B.; Salati, M.; Edeline, J.; La Casta, A.; Bittoni, A.; Berardi, R.; Bruix, J.; Valle, J.W. Medical treatment for cholangiocarcinoma. Liver Int. 2019, 39, 123–142. [Google Scholar] [CrossRef] [Green Version]
- De la Fouchardiere, C. Towards greater clarity in the treatment of cholangiocarcinoma. Lancet Oncol. 2020, 21, 738–739. [Google Scholar] [CrossRef]
- Salati, M.; Caputo, F.; Baldessari, C.; Carotenuto, P.; Messina, M.; Caramaschi, S.; Dominici, M.; Bonetti, L.R. The Evolving Role of FGFR2 Inhibitors in Intrahepatic Cholangiocarcinoma: From Molecular Biology to Clinical Targeting. Cancer Manag. Res. 2021, 13, 7747–7757. [Google Scholar] [CrossRef]
- Franco, B.; Clarke, P.; Carotenuto, P. Pemigatinib. Fibroblast growth factor receptor inhibitor, Treatment of cholangiocarci-noma. Drugs Future 2019, 44, 923. [Google Scholar] [CrossRef]
- Guo, X.; Shen, W. Latest evidence on immunotherapy for cholangiocarcinoma (Review). Oncol. Lett. 2020, 20, 381. [Google Scholar] [CrossRef]
- Carotenuto, P.; Hedayat, S.; Fassan, M.; Cardinale, V.; Lampis, A.; Guzzardo, V.; Braconi, C. Modulation of Biliary Cancer Chemo-Resistance Through MicroRNA-Mediated Rewiring of the Expansion of CD133+ Cells. Hepatology 2020, 72, 982–996. [Google Scholar] [CrossRef] [PubMed]
- Salati, M.; Braconi, C. Noncoding RNA in Cholangiocarcinoma. Semin. Liver Dis. 2018, 39, 013–025. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anastasiadou, E.; Jacob, L.S.; Slack, F.J. Non-coding RNA networks in cancer. Nat. Rev. Cancer 2018, 18, 5–18. [Google Scholar] [CrossRef] [PubMed]
- Gebert, L.F.R.; Macrae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef] [PubMed]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef]
- Adrian, C.G.; Dan, D.C.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Croce, C.M. Frequent Deletions and Down-Regulation of Micro-RNA Genes miR15 and miR16 at 13q14 in Chronic Lymphocytic Leukemia [Internet]. Available online: https://www.pnas.org/doi/abs/10.1073/pnas.242606799 (accessed on 22 January 2022).
- Wang, P.; Lv, L. miR-26a induced the suppression of tumor growth of cholangiocarcinoma via KRT19 approach. Oncotarget 2016, 7, 81367–81376. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.-T.; Sun, J.; Yan, Y.; Cui, S.-H.; Wang, H.; Wang, C.-H.; Zhang, Q. Encapsulated microRNA by gemcitabine prodrug for cancer treatment. J. Control. Release 2019, 316, 317–330. [Google Scholar] [CrossRef]
- Zhang, J.; Han, C.; Wu, T. MicroRNA-26a Promotes Cholangiocarcinoma Growth by Activating β-catenin. Gastroenterology 2012, 143, 246–256.e8. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.-J.; He, C.-C.; Sui, X.; Cai, M.-J.; Zhou, C.-Y.; Ma, J.-L.; Zhu, Q. MiR-21 Promotes Intrahepatic Cholangiocarcinoma Proliferation and Growth In Vitro and In Vivo by Targeting PTPN14 and PTEN [Internet]. Volume 6. Available online: www.impactjournals.com/oncotarget (accessed on 22 January 2022).
- Liu, Z.; Jin, Z.-Y.; Liu, C.-H.; Xie, F.; Lin, X.-S.; Huang, Q. MicroRNA-21 regulates biological behavior by inducing EMT in human cholangiocarcinoma. Int. J. Clin. Exp. Pathol. 2015, 8, 4684–4694. [Google Scholar]
- Meng, F.; Henson, R.; Lang, M.; Wehbe, H.; Maheshwari, S.; Mendell, J.T.; Jiang, J.; Schmittgen, T.D.; Patel, T. Involvement of Human Micro-RNA in Growth and Response to Chemotherapy in Human Cholangiocarcinoma Cell Lines. Gastroenterology 2006, 130, 2113–2129. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhou, Z.-Q.; Yang, Z.-R.; Tong, D.-N.; Guan, J.; Shi, B.-J.; Nie, J.; Ding, X.-T.; Li, B.; Zhou, G.-W.; et al. MicroRNA-191 acts as a tumor promoter by modulating the TET1-p53 pathway in intrahepatic cholangiocarcinoma. Hepatology 2017, 66, 136–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Indrieri, A.; Carrella, S.; Carotenuto, P.; Banfi, S.; Franco, B. The Pervasive Role of the miR-181 Family in Development, Neurodegeneration, and Cancer. Int. J. Mol. Sci. 2020, 21, 2092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, Z.L.; Zhang, F.X.; Zhan, H.L.; Yang, H.J.; Zhang, S.Y.; Liu, Z.H.; Ke, R.S. miR-181b-5p Promotes the Progression of Cholangiocarcinoma by Targeting PARK2 via PTEN/PI3K/AKT Signaling Pathway. Biochem. Genet. 2022, 60, 223–240. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Zhu, J.; Wu, B.; Chen, J.; Zhu, Z.; Cai, P.; Guo, W.; Gu, Z.; Wang, J.; Huang, S. Diagnostic and prognostic value of microRNAs in cholangiocarcinoma: A systematic review and meta-analysis. Cancer Manag. Res. 2018, 10, 2125–2139. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Xie, C.; Pan, S.; Liang, Y.; Han, J.; Lan, Y.; Liu, L. N-myc downstream-regulated gene 2 inhibits human cholangiocarci-noma progression and is regulated by leukemia inhibitory factor/MicroRNA-181c negative feedback pathway. Hepatology 2016, 64, 1606–1622. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.W.; Wang, X.; Li, G.C.; Wang, D.; Han, S.; Zhang, Y.D.; Luo, C.H.; Wang, H.W.; Jiang, W.J.; Li, C.X.; et al. MiR-30a-5p promotes cholangiocarcinoma cell proliferation through targeting SOCS3. J. Cancer 2020, 11, 3604–3614. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Ma, L.; Wang, J. Overexpression of miR-25 is associated with progression and poor prognosis of cholangiocarcinoma. Exp. Ther. Med. 2019, 18, 2687–2694. [Google Scholar] [CrossRef] [Green Version]
- Razumilava, N.; Bronk, S.F.; Smoot, R.L.; Fingas, C.D.; Werneburg, N.W.; Roberts, L.; Mott, J.L. miR-25 targets TNF-related apoptosis inducing ligand (TRAIL) death receptor-4 and promotes apoptosis resistance in cholangiocarcinoma. Hepatology 2011, 55, 465–475. [Google Scholar] [CrossRef]
- Gao, L.; Yang, X.; Zhang, H.; Yu, M.; Long, J.; Yang, T. Inhibition of miR-10a-5p suppresses cholangiocarcinoma cell growth through downregulation of Akt pathway. OncoTargets Ther. 2018, 11, 6981–6994. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Meng, F.; Venter, J.; Wu, N.; Wan, Y.; Standeford, H.; Alpini, G. miR-34a-dependent overexpression of Per1 decreases chol-angiocarcinoma growth. J. Hepatol. 2016, 64, 1295–2130. [Google Scholar] [CrossRef] [Green Version]
- Slabáková, E.; Culig, Z.; Remšík, J.; Souček, K. Alternative mechanisms of miR-34a regulation in cancer. Cell Death Dis. 2017, 8, e3100. [Google Scholar] [CrossRef] [PubMed]
- Au, S.L.-K.; Wong, C.C.L.; Lee, J.M.-F.; Fan, D.N.-Y.; Tsang, F.H.; Ng, I.O.-L.; Wong, C.-M. Enhancer of zeste homolog 2 epigenetically silences multiple tumor suppressor microRNAs to promote liver cancer metastasis. Hepatology 2012, 56, 622–631. [Google Scholar] [CrossRef]
- Strillacci, A.; Griffoni, C.; Sansone, P.; Paterini, P.; Piazzi, G.; Lazzarini, G.; Tomasi, V. MiR-101 downregulation is involved in cycloox-ygenase-2 overexpression in human colon cancer cells. Exp. Cell Res. 2009, 315, 1439–1447. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Yang, J.-R.; Xu, T.; Huang, J.; Xu, L.; Yuan, Y.; Zhuang, S.M. MicroRNA-101, down-regulated in hepatocellular carcinoma, pro-motes apoptosis and suppresses tumorigenicity. Cancer Res. 2009, 69, 1135–1142. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Han, C.; Zhu, H.; Song, K.; Wu, T. miR-101 Inhibits Cholangiocarcinoma Angiogenesis through Targeting Vascular Endothelial Growth Factor (VEGF). Am. J. Pathol. 2013, 182, 1629–1639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Li, Y.; Ding, M.; Zhang, H.; Xu, X.-M.; Tang, J. Molecular mechanisms and clinical applications of miR-22 in regulating malignant progression in human cancer (Review). Int. J. Oncol. 2016, 50, 345–355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansini, A.P.; Pisarello, M.J.L.; Thelen, K.M.; Cruz-Reyes, M.; Peixoto, E.; Jin, S.; Howard, B.N.; Trussoni, C.E.; Gajdos, G.B.; LaRusso, N.F.; et al. MicroRNA (miR)-433 and miR-22 dysregulations induce histone-deacetylase-6 overexpression and ciliary loss in cholangiocarcinoma. Hepatology 2018, 68, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Calin, G.A. miR-122 and hepatocellular carcinoma: From molecular biology to therapeutics. EBioMedicine 2018, 37, 17–18. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Liu, G.; Zhang, M.; Zhang, Z.; Jia, Y.; Peng, L.; Zhu, Y.; Hu, J.; Huang, R.; Sun, X. miR-122-5p Inhibits the Proliferation, Invasion and Growth of Bile Duct Carcinoma Cells by Targeting ALDOA. Cell. Physiol. Biochem. 2018, 48, 2596–2606. [Google Scholar] [CrossRef]
- Kong, L.; Wu, Q.; Zhao, L.; Ye, J.; Li, N.; Yang, H. Upregulated lncRNA-UCA1 contributes to metastasis of bile duct carcinoma through regulation of miR-122/CLIC1 and activation of the ERK/MAPK signaling pathway. Cell Cycle 2019, 18, 1212–1228. [Google Scholar] [CrossRef]
- Hozaka, Y.; Seki, N.; Tanaka, T.; Asai, S.; Moriya, S.; Idichi, T.; Wada, M.; Tanoue, K.; Kawasaki, Y.; Mataki, Y.; et al. Molecular Pathogenesis and Regulation of the miR-29-3p-Family: Involvement of ITGA6 and ITGB1 in Intra-Hepatic Cholangiocarcinoma. Cancers 2021, 13, 2804. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Wang, X.; Chen, P. MiR-204 down regulates SIRT1 and reverts SIRT1-induced epithelial-mesenchymal transition, anoikis resistance and invasion in gastric cancer cells. BMC Cancer 2013, 13, 290–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, T.; Lau, T.; Cheung, T.; Yim, S.; Lo, K.; Siu, N.; Chan, L.; Yu, M.; Kwong, J.; Doran, G.; et al. Dysregulation of microRNA-204 mediates migration and invasion of endometrial cancer by regulating FOXC1. Int. J. Cancer 2011, 130, 1036–1045. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Li, S.; Tang, L. MicroRNA 320, an Anti-Oncogene Target miRNA for Cancer Therapy. Biomedicines 2021, 9, 591. [Google Scholar] [CrossRef]
- Zhu, H.; Jiang, X.; Zhou, X.; Dong, X.; Xie, K.; Yang, C.; Jiang, H.; Sun, X.; Lu, J. Neuropilin-1 regulated by miR-320 contributes to the growth and metastasis of cholangiocarcinoma cells. Liver Int. 2017, 38, 125–135. [Google Scholar] [CrossRef]
- Olaru, A.V.; Ghiaur, G.; Yamanaka, S.; Luvsanjav, D.; An, F.; Popescu, I.; Selaru, F.M. MicroRNA down-regulated in human cholangio-carcinoma control cell cycle through multiple targets involved in the G1/S checkpoint. Hepatology 2011, 54, 2089–2098. [Google Scholar] [CrossRef] [Green Version]
- Chen, T.; Lei, S.; Zeng, Z.; Pan, S.; Zhang, J.; Xue, Y.; Sun, Y.; Lan, J.; Xu, S.; Mao, D.; et al. MicroRNA-137 suppresses the proliferation, migration and invasion of cholangiocarcinoma cells by targeting WNT2B. Int. J. Mol. Med. 2020, 45, 886–896. [Google Scholar] [CrossRef]
- Fu, W.; Yu, G.; Liang, J.; Fan, P.; Dong, K.; Zhang, B.; Chen, X.; Zhu, H.; Chu, L. miR-144-5p and miR-451a Inhibit the Growth of Cholangiocarcinoma Cells Through Decreasing the Expression of ST8SIA4. Front. Oncol. 2021, 10, 3104. [Google Scholar] [CrossRef]
- Meng, F.; Wehbe-Janek, H.; Henson, R.; Smith, H.; Patel, T. Epigenetic regulation of microRNA-370 by interleukin-6 in malig-nant human cholangiocytes. Oncogene 2008, 27, 378–386. [Google Scholar] [CrossRef] [Green Version]
- An, F.; Yamanaka, S.; Allen, S.; Roberts, L.R.; Gores, G.J.; Pawlik, T.M.; Xie, Q.; Ishida, M.; Mezey, E.; Ferguson-Smith, A.C.; et al. Silencing of miR-370 in Human Cholangiocarcinoma by Allelic Loss and Interleukin-6 Induced Maternal to Paternal Epigenotype Switch. PLoS ONE 2012, 7, e45606. [Google Scholar] [CrossRef] [Green Version]
- Omar, H.A.; El-Serafi, A.T.; Hersi, F.; Arafa, E.A.; Zaher, D.M.; Madkour, M.; Arab, H.H.; Tolba, M.F. Immunomodulatory MicroRNAs in cancer: Targeting immune checkpoints and the tumor microenvironment. FEBS J. 2019, 286, 3540–3557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, T.; Morishita, A.; Kobara, H.; Masaki, T. The Role of microRNAs in Cholangiocarcinoma. Int. J. Mol. Sci. 2021, 22, 7627. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Yan, H.-X.; Yang, W.; Hu, L.; Yu, L.-X.; Liu, Q.; Li, L.; Huang, D.-D.; Ding, J.; Shen, F. The role of microRNA expression pattern in human intrahepatic cholangiocarcinoma. J. Hepatol. 2009, 50, 358–369. [Google Scholar] [CrossRef] [PubMed]
- Peng, F.; Jiang, J.; Yu, Y.; Tian, R.; Guo, X.; Li, X.; Qin, R. Direct targeting of SUZ12/ROCK2 by miR-200b/c inhibits cholangiocarcinoma tumourigenesis and metastasis. Br. J. Cancer 2013, 109, 3092–3104. [Google Scholar] [CrossRef] [Green Version]
- Park, S.-M.; Gaur, A.B.; Lengyel, E.; Peter, M.E. The miR-200 family determines the epithelial phenotype of cancer cells by target-ing the E-cadherin repressors ZEB1 and ZEB2. Genes Dev. 2008, 22, 894–907. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Wang, Y.-N.; Song, D.-J.; Tan, J.-P.; Cao, Y.; Fan, J.; Zhou, J. A High-Accuracy Model Based on Plasma miRNAs Diagnoses In-trahepatic Cholangiocarcinoma: A Single Center with 1001 Samples. Diagnostics 2021, 11, 610. [Google Scholar] [CrossRef]
- Macias, R.I.; Banales, J.; Sangro, B.; Muntané, J.; Avila, M.; Lozano, E.; Perugorria, M.J.; Padillo, F.J.; Bujanda, L.; Marin, J. The search for novel diagnostic and prognostic biomarkers in cholangiocarcinoma. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2018, 1864, 1468–1477. [Google Scholar] [CrossRef]
- Lawrie, C.H.; Gal, S.; Dunlop, H.M.; Pushkaran, B.; Liggins, A.P.; Pulford, K.; Banham, A.H.; Pezzella, F.; Boultwood, J.; Wainscoat, J.S.; et al. Detection of elevated levels of tumour-associated microRNAs in serum of patients with diffuse large B-cell lymphoma. Br. J. Haematol. 2008, 141, 672–675. [Google Scholar] [CrossRef]
- Mitchell, P.S.; Parkin, R.K.; Kroh, E.M.; Fritz, B.R.; Wyman, S.K.; Pogosova-Agadjanyan, E.L.; Tewari, M. Circulating microRNAs as stable blood-based markers for cancer detection. Proc. Nat. Acad. Sci. USA 2008, 105, 10513–10518. [Google Scholar] [CrossRef] [Green Version]
- Toiyama, Y.; Takahashi, M.; Hur, K.; Nagasaka, T.; Tanaka, K.; Inoue, Y.; Goel, A. Serum miR-21 as a diagnostic and prognostic biomarker in colorectal cancer. J. Natl. Cancer Inst. 2013, 105, 849–859. [Google Scholar] [CrossRef] [Green Version]
- Selaru, F.M.; Olaru, A.V.; Kan, T.; David, S.; Cheng, Y.; Mori, Y.; Meltzer, S.J. MicroRNA-21 is overexpressed in human cholangiocarci-noma and regulates programmed cell death 4 and tissue inhibitor of metalloproteinase 3. Hepatology 2009, 49, 1595–1601. [Google Scholar] [CrossRef]
- Silakit, R.; Loilome, W.; Yongvanit, P.; Thongchot, S.; Sithithaworn, P.; Boonmars, T.; Namwat, N. Urinary microRNA-192 and mi-croRNA-21 as potential indicators for liver fluke-associated cholangiocarcinoma risk group. Parasitol. Int. 2017, 66, 479–485. [Google Scholar] [CrossRef] [PubMed]
- Kishimoto, T.; Eguchi, H.; Nagano, H.; Kobayashi, S.; Akita, H.; Hama, N.; Wada, H.; Kawamoto, K.; Tomokuni, A.; Tomimaru, Y.; et al. Plasma miR-21 is a novel diagnostic biomarker for biliary tract cancer. Cancer Sci. 2013, 104, 1626–1631. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.-Y.; Zhang, Q.; Wang, D.-D.; Yan, W.; Sha, H.-H.; Zhao, J.-H.; Yang, S.-J.; Zhang, H.-D.; Hou, J.-C.; Xu, H.-Z.; et al. MiR-29a: A potential therapeutic target and promising biomarker in tumors. Biosci. Rep. 2018, 38, BSR20171265. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Chen, Y. Increased Expression of miR-29a and Its Prognostic Significance in Patients with Cholangiocarcinoma. Oncol. Res. Treat. 2017, 40, 128–132. [Google Scholar] [CrossRef]
- Wang, F.; Ren, X.; Zhang, X. Role of microRNA-150 in solid tumors. Oncol. Lett. 2015, 10, 11–16. [Google Scholar] [CrossRef] [Green Version]
- Salem, P.E.S.; Ghazala, R.A.; El Gendi, A.M.; Emara, D.M.; Ahmed, N.M. The association between circulating MicroRNA-150 level and cholangiocarcinoma. J. Clin. Lab. Anal. 2020, 34, e23397. [Google Scholar] [CrossRef]
- Cheng, Q.; Feng, F.; Zhu, Z.; Zheng, Y.; Luo, X.; Liu, C.; Jiang, X. Circulating miR-106a is a Novel Prognostic and Lymph Node Me-tastasis Indicator for Cholangiocarcinoma. Sci. Rep. 2015, 5, 16103. Available online: www.nature.com/scientificreports (accessed on 22 January 2022). [CrossRef]
- Zhang, R.-X.; Zheng, Z.; Li, K.; Wu, X.-H.; Zhu, L. Both plasma and tumor tissue miR-146a high expression correlates with prolonged overall survival of surgical patients with intrahepatic cholangiocarcinoma. Medicine 2017, 96, e8267. [Google Scholar] [CrossRef]
- Li, J.; Gao, B.; Huang, Z.; Duan, T.; Li, D.; Zhang, S.; Cheng, K. Prognostic significance of microRNA-203 in cholangiocarcinoma [Internet]. Int. J. Clin. Exp. Pathol. 2015, 8, 9512. Available online: www.ijcep.com/ (accessed on 22 January 2022).
- Chen, Q.; Wang, C.; Zhang, H.; Li, Y.; Cao, Y.; Zhang, Y.; Han, X. Expression levels of serum miRNA-195 in different types of pa-tients with cholangiocarcinoma and its value to determine the prognosis thereof. Oncol. Let. 2018, 15, 5947–5951. [Google Scholar]
- Meijer, L.L.; Puik, J.R.; le Large, T.Y.S.; Heger, M.; Dijk, F.; Funel, N. Unravelling the Diagnostic Dilemma: A MicroRNA Panel of Circulating MiR-16 and MiR-877 as A Diagnostic Classifier for Distal Bile Duct Tumors. Cancers. 2019, 11, 1181. Available online: www.mdpi.com/journal/cancers (accessed on 22 January 2022). [CrossRef] [PubMed] [Green Version]
- McNally, M.E.; Collins, A.; Wojcik, S.E.; Liu, J.; Henry, J.C.; Jiang, J.; Bloomston, M. Concomitant dysregulation of microRNAs miR-151-3p and miR-126 correlates with improved survival in resected cholangiocarcinoma. HPB 2013, 15, 260–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, J.; Sun, L.; Li, J.; Zhou, C.; Cheng, L.; Chen, K.; Duan, W. A novel three miRNA signature predicts survival in cholangiocarcino-ma based on RNA Seq data. Oncol. Rep. 2018, 40, 1422–1434. [Google Scholar]
- Li, L.; Masica, D.; Ishida, M.; Tomuleasa, C.; Umegaki, S.; Kalloo, A.N.; Georgiades, C.; Singh, V.K.; Khashab, M.; Amateau, S. Human Bile Contains MicroRNA-Laden Extracellu-lar Vesicles That Can Be Used for Cholangiocarcinoma Diagnosis. Hepatology 2014, 60, 896–907. [Google Scholar] [CrossRef]
- Salati, M.; Rizzo, A.; Merz, V.; Messina, C.; Francesco, C.; Gelsomino, F.; Dominici, M. Third-line chemotherapy in advanced biliary cancers (ABC): Pattern of care, treatment outcome and prognostic factors from a multicenter study. Expert Rev. Gastroenterol. Hepatol. 2022, 16, 73–79. [Google Scholar] [CrossRef]
- Rizvi, S.; Gores, G.J. Emerging molecular therapeutic targets for cholangiocarcinoma. J. Hepatol. 2017, 67, 632–644. [Google Scholar] [CrossRef]
- Personeni, N.; Lleo, A.; Pressiani, T.; Colapietro, F.; Openshaw, M.R.; Stavraka, C.; Pouptsis, A.; Pinato, D.J.; Rimassa, L. Biliary Tract Cancers: Molecular Heterogeneity and New Treatment Options. Cancers 2020, 12, 3370. [Google Scholar] [CrossRef]
- Ghidini, M.; Cascione, L.; Carotenuto, P.; Lampis, A.; Trevisani, F.; Previdi, M.C.; Hahne, J.C.; Said-Huntingford, I.; Raj, M.; Zerbi, A.; et al. Characterisation of the immune-related transcriptome in resected biliary tract cancers. Eur. J. Cancer 2017, 86, 158–165. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Dong, C.; Ji, C. MicroRNA and drug resistance. Cancer Gene Ther. 2010, 17, 523–531. Available online: www.nature.com/cgt (accessed on 22 January 2022). [CrossRef] [Green Version]
- Okamoto, K.; Miyoshi, K.; Murawaki, Y. miR-29b, miR-205 and miR-221 Enhance Chemosensitivity to Gemcitabine in HuH28 Human Cholangiocarcinoma Cells. PLoS ONE 2013, 8, e77623. [Google Scholar] [CrossRef] [PubMed]
- Silakit, R.; Kitirat, Y.; Thongchot, S.; Loilome, W.; Techasen, A.; Ungarreevittaya, P.; Khuntikeo, N.; Yongvanit, P.; Yang, J.H.; Kim, N.H.; et al. Potential role of HIF-1-responsive microRNA210/HIF3 axis on gemcitabine resistance in cholangiocarcinoma cells. PLoS ONE 2018, 13, e0199827. [Google Scholar] [CrossRef] [PubMed]
- Asukai, K.; Kawamoto, K.; Eguchi, H.; Konno, M.; Asai, A.; Iwagami, Y.; Yamada, D.; Asaoka, T.; Noda, T.; Wada, H.; et al. Micro-RNA-130a-3p Regulates Gemcitabine Resistance via PPARG in Cholangiocarcinoma. Ann. Surg. Oncol. 2017, 24, 2344–2352. [Google Scholar] [CrossRef]
- Li, Q.; Xia, X.; Ji, J.; Ma, J.; Tao, L.; Mo, L.; Chen, W. Oncotarget 33621 MiR-199a-3p enhances cisplatin sensitivity of cholangiocarcinoma cells by inhibiting mTOR signaling pathway and expression of MDR1. Oncotarget 2017, 8, 33621. [Google Scholar] [CrossRef] [Green Version]
- Jiao, D.; Yan, Y.; Shui, S.; Wu, G.; Ren, J.; Wang, Y.; Han, X. miR-106b regulates the 5-fluorouracil resistance by targeting Zbtb7a in cholangiocarcinoma. Oncotarget 2017, 8, 52913. [Google Scholar] [CrossRef] [Green Version]
- Lampis, A.; Carotenuto, P.; Vlachogiannis, G.; Cascione, L.; Hedayat, S.; Burke, R.; Clarke, P.A.; Bosma, E.; Simbolo, M.; Scarpa, A.; et al. MIR21 Drives Resistance to Heat Shock Protein 90 Inhibition in Cholangiocarcinoma. Gastroenterology 2018, 154, 1066–1079.e5. [Google Scholar] [CrossRef] [Green Version]
- Boilève, A.; Hilmi, M.; Smolenschi, C.; Ducreux, M.; Hollebecque, A.; Malka, D. Immunotherapy in Advanced Biliary Tract Cancers. Cancers 2021, 13, 1569. [Google Scholar] [CrossRef]
- Clinical Trial Database. Available online: www.clinicaltrials.gov (accessed on 22 January 2022).
- Li, L.; Piontek, K.; Ishida, M.; Fausther, M.; Dranoff, J.A.; Fu, R.; Selaru, F.M. Extracellular vesicles carry micro RNA -195 to intrahepatic cholangiocarcinoma and improve survival in a rat model. Hepatology 2017, 65, 501–514. [Google Scholar] [CrossRef]
- Hong, D.S.; Kang, Y.-K.; Brenner, A.J.; Sachdev, J.C.; Ejadi, S.; Borad, M.J.; Kim, T.-Y.; Lim, H.Y.; Park, K.; Becerra, C.; et al. MRX34, a liposomal miR-34 mimic, in patients with advanced solid tumors: Final dose-escalation results from a first-in-human phase I trial of microRNA therapy. J. Clin. Oncol. 2016, 34, 2508. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, Y.; Li, J.; Hang, Y.; Jaramillo, L.; Wehrkamp, C.J.; Phillippi, M.A.; Mohr, A.M.; Chen, Y.; Talmon, G.A.; et al. Cholangiocarcinoma therapy with nanoparticles that combine downregulation of MicroRNA-210 with inhibition of cancer cell invasiveness. Theranostics 2018, 8, 4305–4320. [Google Scholar] [CrossRef]


{kind=link}
| miRNA | Target Gene | Mechanism | References |
|---|---|---|---|
| miR-26a | GSK-3β; KRT19 | Proliferation, migration, and invasion | [17,18,19] |
| miR-21 | PTEN; PDCD4; TIMP3; PTPN14; 15-PGDH/HPGD | Proliferation, apoptosis, EMT, inflammation | [20,21,22] |
| miR-191 | TET | Proliferation, invasion, and migration | [23] |
| miR-181b-5p | PARK2 | Proliferation, migration, and invasion | [24,25,26] |
| miR-181c | NDRG2 | Proliferation, drug-resistance, and metastasis | [27] |
| MiR-30a-5p | SOCS3 | Proliferation | [28] |
| miR-25 | DR4 | Proliferation, invasion, and apoptosis | [29,30] |
| miR-10a-5p | PTEN | Proliferation | [31] |
| miRNA | Target Gene | Mechanism | References |
|---|---|---|---|
| miR-34 | MYC, MET, CDK4/6, BCL2, CD44, NOTCH1, NOTCH2, JAGGED1 | Proliferation, apoptosis | [32,33] |
| miR-101 | EZH2, COX-2, APP, MCL-1, VEGF | Proliferation, apoptosis; angiogenesis, inflammation; transcriptional repression | [34,35,36,37] |
| miR-22 | SIRT1, CDK6, SP1, HDAC6 | Proliferation, senescence, invasion, metastasis; ciliogenesis, histone modifications | [38,39] |
| miR-122 | ALDOA, CLIC1 | Proliferation and invasion | [40,41,42] |
| miR-29-3p | ITGA6, ITGB1 | Cell migration and invasion | [43] |
| miR-204 | SLUG | Cell migration, invasion, EMT | [44,45] |
| miR-320 | VEGF, NRP-1 | Proliferation, invasion, EMT, tumor migration, and metastasis | [46,47] |
| miR-494 | CCNB1, CDK2, CDK4, CDK6, CCND1, CCNE2, HDAC1, RB1, PLK1, PTTG1, TOP2A | Proliferation, cell cycle | [48] |
| miR-137 | WNT2B | Proliferation, migration, and invasion | [49] |
| miR-144-5p/miR-451 | ST8SIA4 | Proliferation, migration, and invasion | [50] |
| miR-370 | MAP3K8 | Proliferation, inflammation, tumor microenvironment | [51,52] |
| miR-148a | RASSF1 | Proliferation, inflammation, tumor microenvironment | [53,54] |
| miR-152 | CDKN2A | Proliferation, inflammation, tumor microenvironment | [53,54] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barbato, A.; Piscopo, F.; Salati, M.; Reggiani-Bonetti, L.; Franco, B.; Carotenuto, P. Micro-RNA in Cholangiocarcinoma: Implications for Diagnosis, Prognosis, and Therapy. J. Mol. Pathol. 2022, 3, 88-103. https://doi.org/10.3390/jmp3020009
Barbato A, Piscopo F, Salati M, Reggiani-Bonetti L, Franco B, Carotenuto P. Micro-RNA in Cholangiocarcinoma: Implications for Diagnosis, Prognosis, and Therapy. Journal of Molecular Pathology. 2022; 3(2):88-103. https://doi.org/10.3390/jmp3020009
Chicago/Turabian StyleBarbato, Anna, Fabiola Piscopo, Massimiliano Salati, Luca Reggiani-Bonetti, Brunella Franco, and Pietro Carotenuto. 2022. "Micro-RNA in Cholangiocarcinoma: Implications for Diagnosis, Prognosis, and Therapy" Journal of Molecular Pathology 3, no. 2: 88-103. https://doi.org/10.3390/jmp3020009