
Solanum elaeagnifolium Invasiveness under Semi-Arid Environmental Conditions in Tunisia


Abstract
:1. Introduction
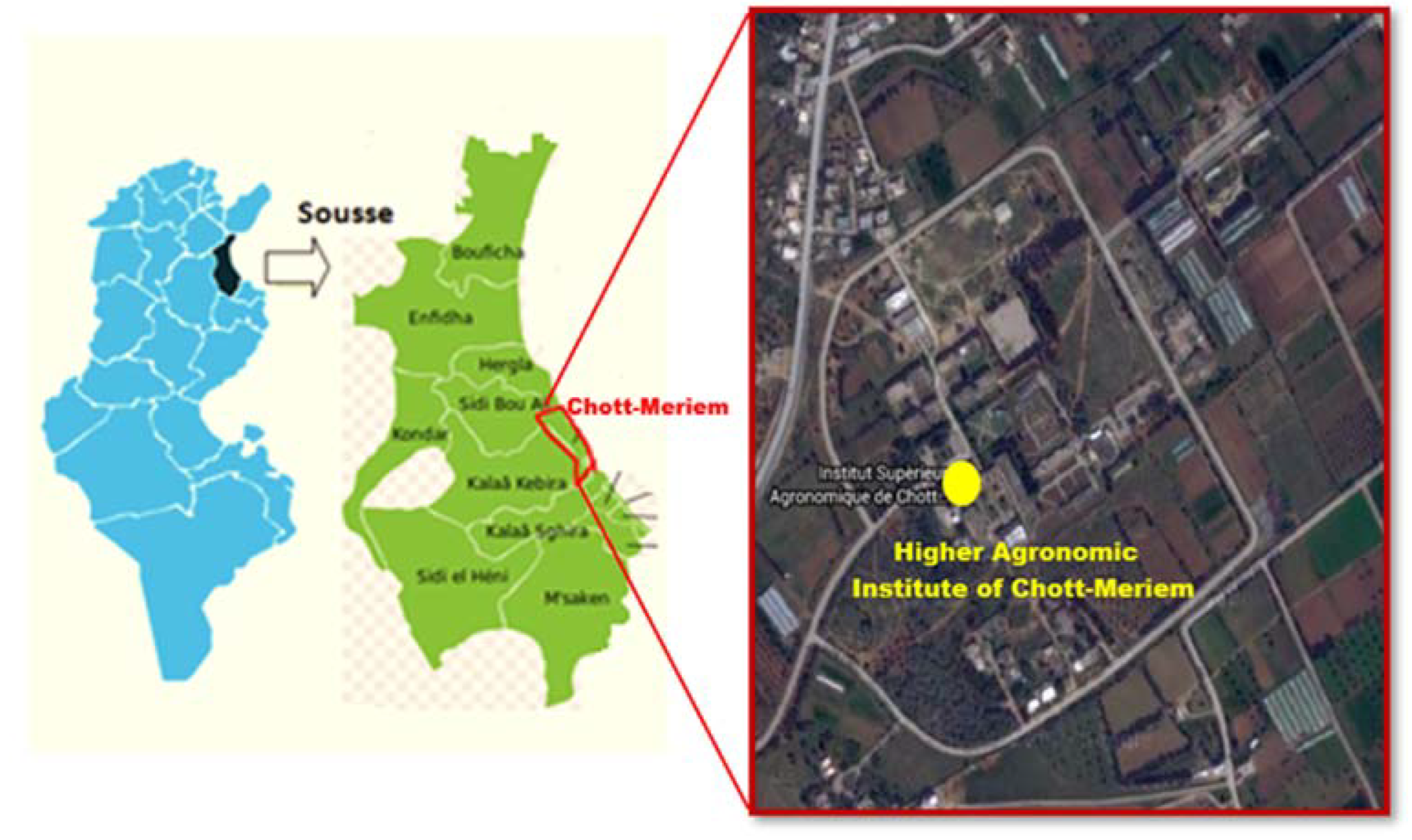
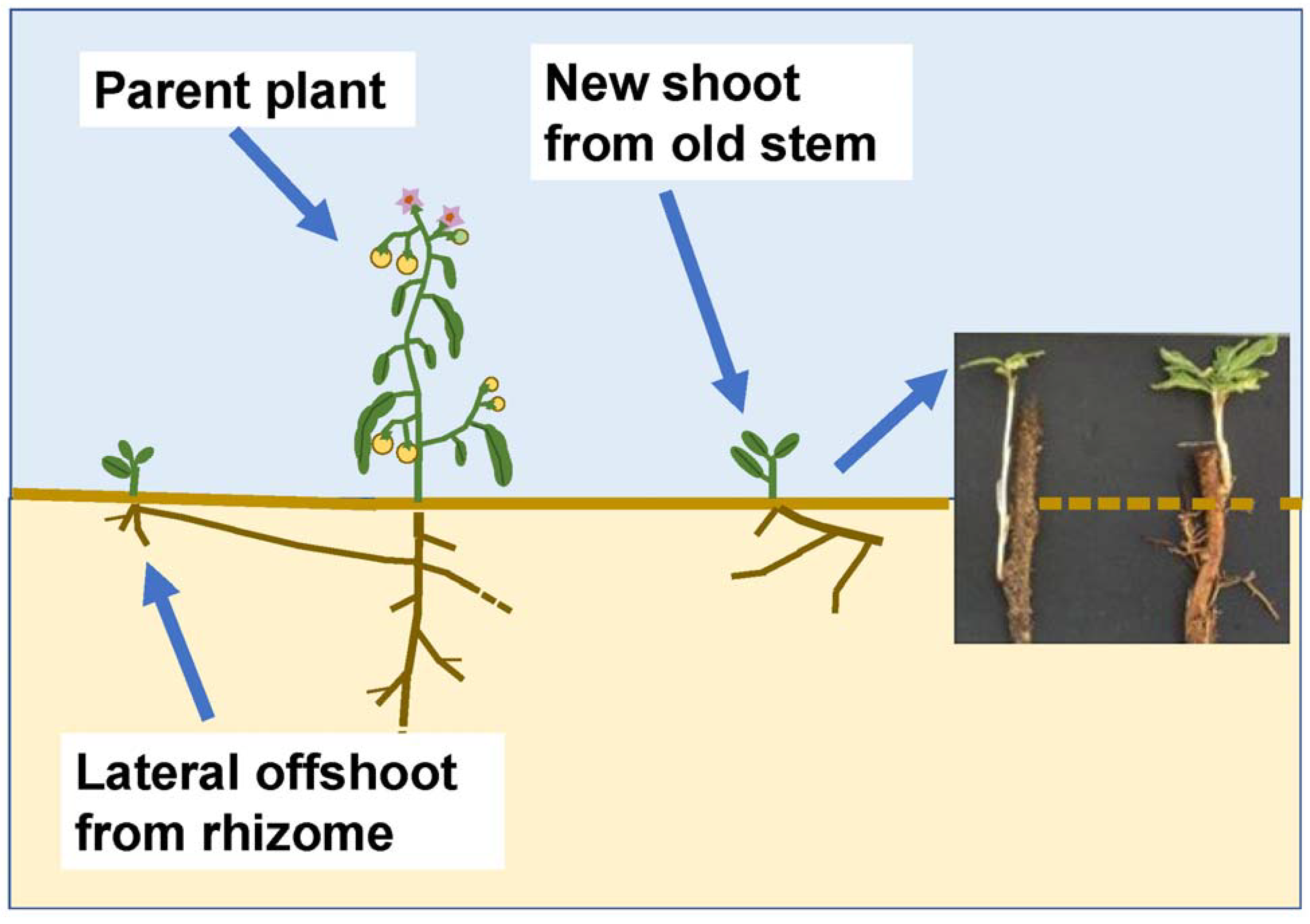
2. Materials and Methods
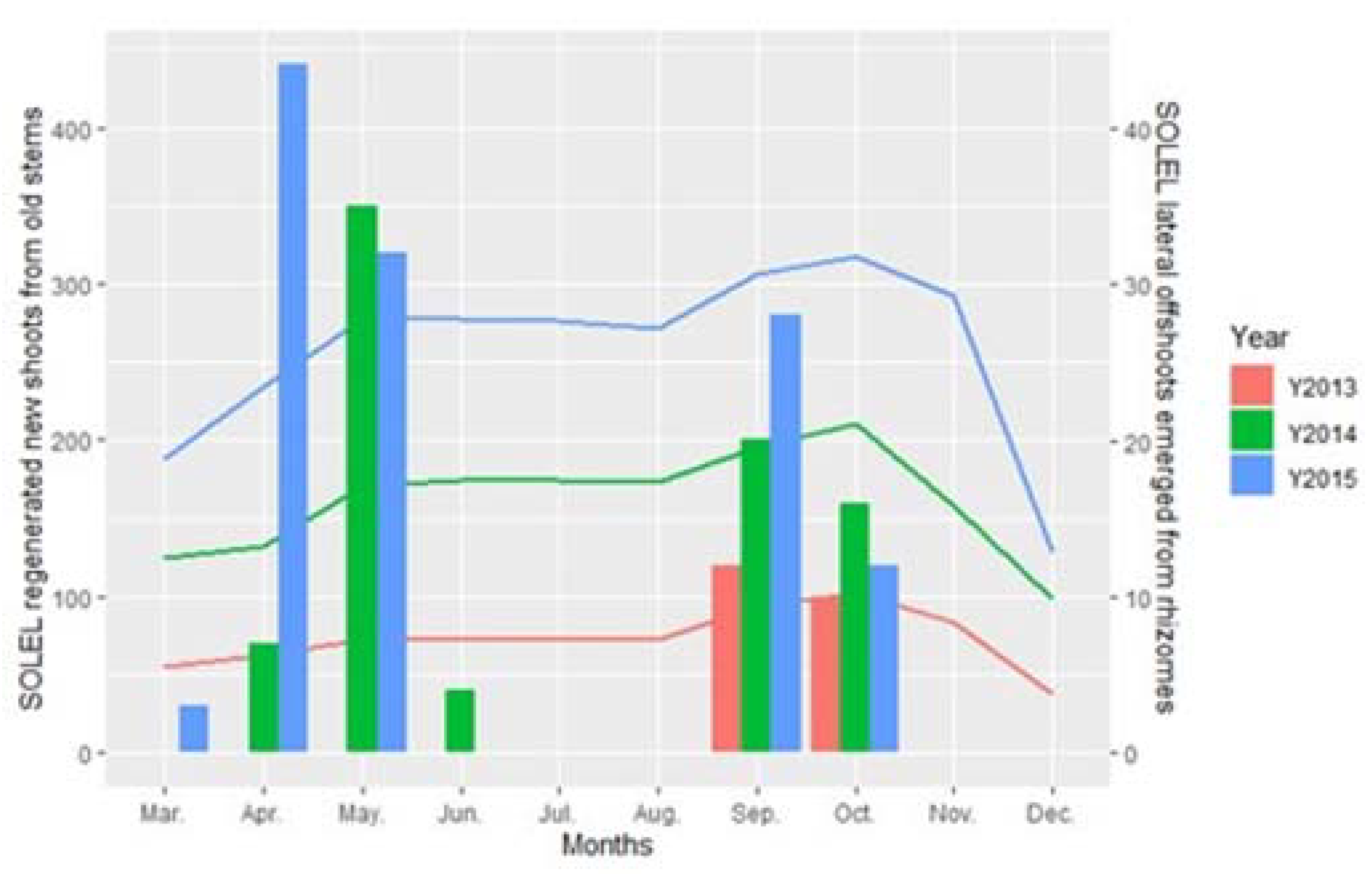
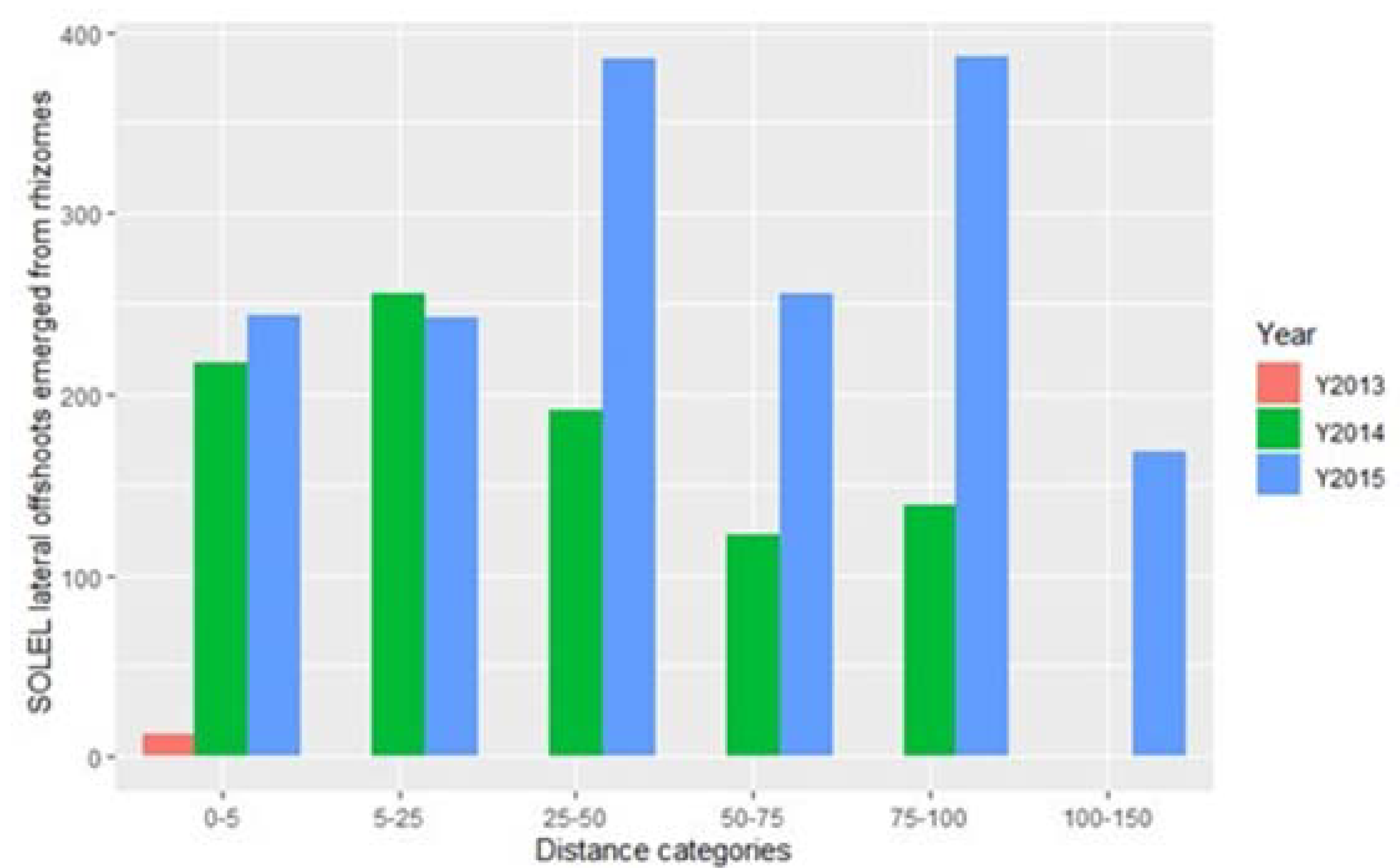
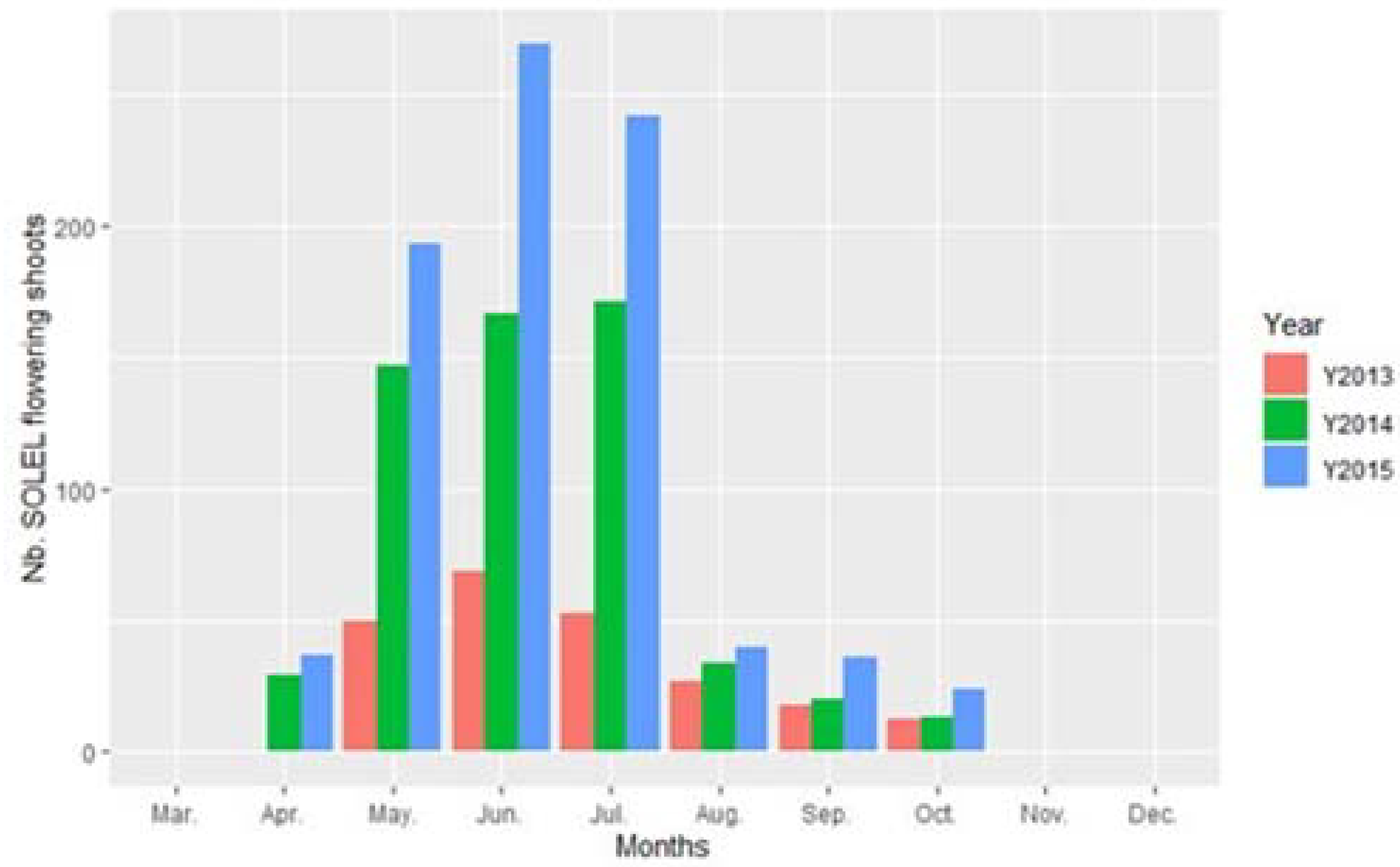
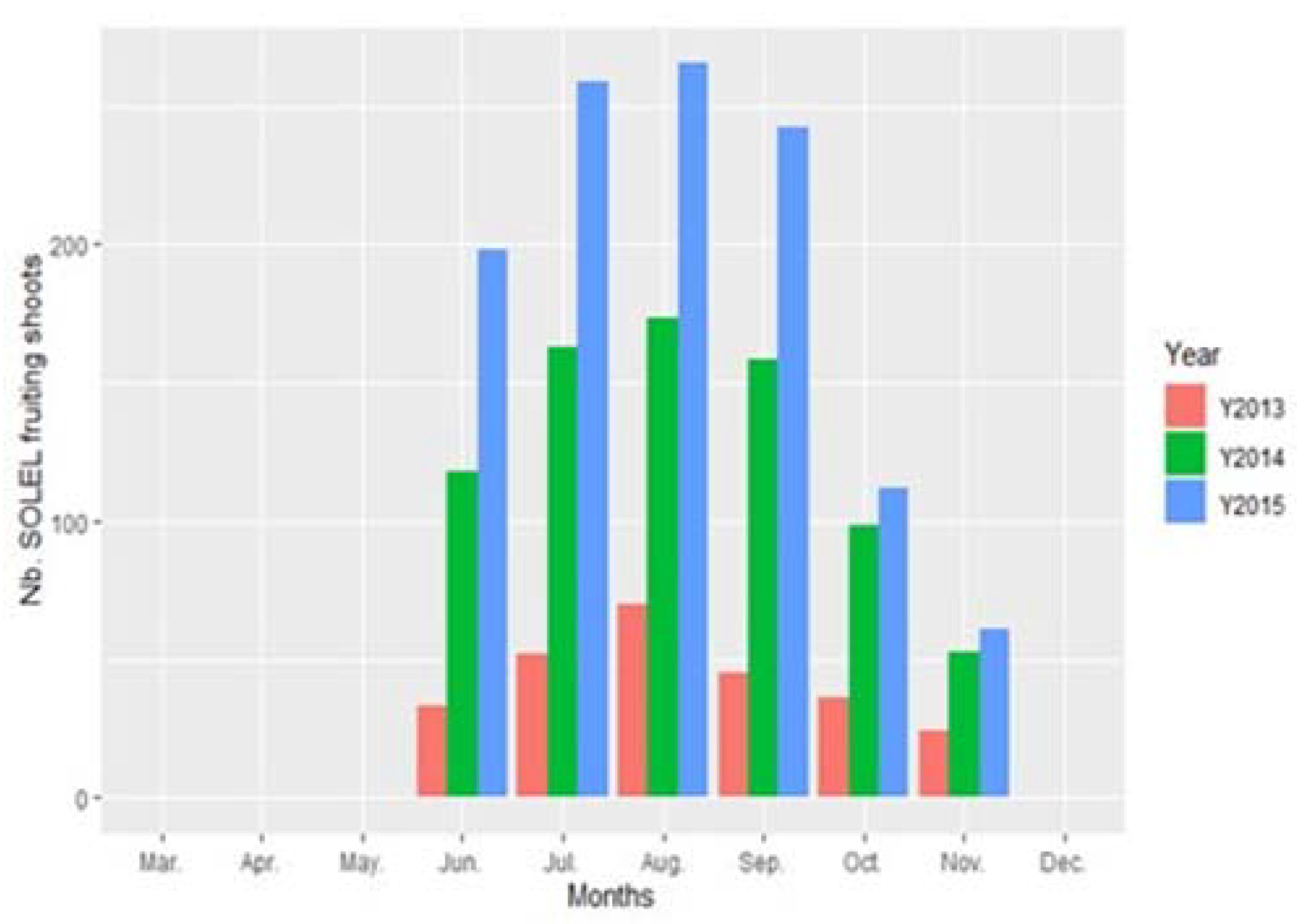
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gallardo, B.; Bacher, S.; Bradley, B.; Comín, F.A.; Gallien, L.; Jeschke, J.M.; Sorte, C.J.B.; Vilà, M. InvasiBES: Understanding and Managing the Impacts of Invasive Alien Species on Biodiversity and Ecosystem Services. NeoBiota 2019, 50, 109–122. [Google Scholar] [CrossRef] [Green Version]
- Vilà, M.; Hulme, P.E. Invading Nature–Springer Series in Invasion Ecology. In Impact of Biological Invasions on Ecosystem Services; Springer: Berlin/Heidelberg, Germany, 2017; Volume 12. [Google Scholar]
- Haubrock, P.J.; Turbelin, A.J.; Cuthbert, R.N.; Novoa, A.; Taylor, N.G.; Angulo, E.; Ballesteros-Mejia, L.; Bodey, T.W.; Capinha, C.; Diagne, C.; et al. Economic Costs of Invasive Alien Species across Europe. NeoBiota 2021, 67, 153–190. [Google Scholar] [CrossRef]
- Fried, G.; Chauvel, B.; Reynaud, P.; Sache, I. Decreases in Crop Production by Non-Native Weeds, Pests, and Pathogens. In Impact of Biological Invasions on Ecosystem Services; Springer: Berlin/Heidelberg, Germany, 2017; pp. 83–101. [Google Scholar]
- Lososová, Z.; Chytrý, M.; Tichý, L.; Danihelka, J.; Fajmon, K.; Hájek, O.; Kintrová, K.; Láníková, D.; Otýpková, Z.; Řehořek, V. Biotic Homogenization of Central European Urban Floras Depends on Residence Time of Alien Species and Habitat Types. Biol. Conserv. 2012, 145, 179–184. [Google Scholar] [CrossRef]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological Impacts of Invasive Alien Plants: A Meta-Analysis of Their Effects on Species, Communities and Ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef]
- Bartz, R.; Kowarik, I. Assessing the Environmental Impacts of Invasive Alien Plants: A Review of Assessment Approaches. NeoBiota 2019, 43, 69–99. [Google Scholar] [CrossRef] [Green Version]
- Knapp, S.; Sagona, E.; Carbonell, A.K.Z.; Chiarini, F. A Revision of the Solanum elaeagnifolium Clade (Elaeagnifolium Clade; Subgenus leptostemonum, Solanaceae). PhytoKeys 2017, 84, 1–104. [Google Scholar] [CrossRef] [Green Version]
- Scaldaferro, M. Geographical Pattern and Ploidy Levels of the Weed Solanum elaeagnifolium (Solanaceae) from Argentina. Genet. Resour. Crop Evol. 2012, 59, 1833–1847. [Google Scholar] [CrossRef]
- Boyd, J.W.; Murray, D.S.; Tyrl, R.J. Silverleaf nightshade, Solarium elaeagnifolium, Origin, Distribution, and Relation to Man. Econ. Bot. 1984, 38, 210–217. [Google Scholar] [CrossRef]
- Krigas, N.; Tsiafouli, M.A.; Katsoulis, G.; Votsi, N.-E.; van Kleunen, M. Investigating the Invasion Pattern of the Alien Plant Solanum elaeagnifolium Cav. (Silverleaf nightshade): Environmental and Human-Induced Drivers. Plants 2021, 10, 805. [Google Scholar] [CrossRef] [PubMed]
- EPPO. Solanum elaeagnifolium. EPPO Bull. 2007, 37, 236–245. [Google Scholar] [CrossRef]
- Brunel, S. Pest Risk Analysis for Solanum elaeagnifolium and International Management Measures Proposed. EPPO Bull. 2011, 41, 232–242. [Google Scholar] [CrossRef]
- Formozis, G.; Tsakaldimi, M.; Ganatsas, P. Are Mediterranean Forest Ecosystems under the Threat of Invasive Species Solanum elaeagnifolium? iForest Biogeosci. For. 2021, 14, 236–241. [Google Scholar] [CrossRef]
- Zhu, X.C.; Wu, H.W.; Stanton, R.; Burrows, G.E.; Lemerle, D.; Raman, H. Morphological Variation of Solanum elaeagnifolium in South-Eastern Australia. Weed Res. 2013, 53, 344–354. [Google Scholar] [CrossRef]
- Uludag, A.; Gbehounou, G.; Kashefi, J.; Bouhache, M.; Bon, M.-C.; Bell, C.; Lagopodi, A.L. Review of the Current Situation for Solanum elaeagnifolium in the Mediterranean Basin. EPPO Bull. 2016, 46, 139–147. [Google Scholar] [CrossRef]
- Qasem, J.R.; Al Abdallat, A.M.; Hasan, S.M. Genetic Diversity of Solanum elaeagnifolium, an Invasive Problematic Weed in Jordan. Weed Res. 2019, 59, 222–234. [Google Scholar] [CrossRef]
- Tataridas, A.; Oliveira, R.S.; Frazão, L.; Moreira, M.; Travlos, I.; Freitas, H. Solanum elaeagnifolium Cav. (Solanales: Solanaceae) Presence Confirmed in Portugal. EPPO Bull. 2022, 52, 499–504. [Google Scholar] [CrossRef]
- Gmira, N.; Douira, A.; Bouhache, M. Ecological Grouping of Solanum elaeagnifolium: A Principal Weed in the Irrigated Tadla Plain (Central Morocco). Weed Res. 1998, 38, 87–94. [Google Scholar] [CrossRef]
- Stanton, R.; Heap, J.; Carter, R.; Wu, H. Solanum elaeagnifolium Cav. In The Biology of Australian Weeds. Volume 3; R.G. and F.J. Richardson: Melbourne, Australia, 2009; pp. 1–35. [Google Scholar]
- Vladimirov, V.; Bancheva, S.; Delcheva, M. Solanum elaeagnifolium (Solanaceae), a New Alien Species for the Bulgarian Flora. Fl. Medit 2015, 25, 121–125. [Google Scholar]
- Tscheulin, T.; Petanidou, T.; Potts, S.G.; Settele, J. The Impact of Solanum elaeagnifolium, an Invasive Plant in the Mediterranean, on the Flower Visitation and Seed Set of the Native Co-Flowering Species Glaucium Flavum. Plant Ecol. 2009, 205, 77–85. [Google Scholar] [CrossRef]
- Karmezi, M.; Krigas, N.; Argyropoulou, M.D. The Invasion and Long Naturalization of Solanum elaeagnifolium Affects the Soil Nematode Community: Evidence from a Comparative Study. Agronomy 2022, 12, 2346. [Google Scholar] [CrossRef]
- Adjim, Z.; Tani, C.K. L’infestation Par Solanum elaeagnifolium Menace l’Algérie. Rev. D’ecologie Terre Vie 2018, 73, 569–581. [Google Scholar]
- Chalghaf, E.; Aissa, M.; Mellassi, H.; Mekki, M. Maîtrise de La Propagation de La Morelle Jaune (Solanum elaeagnifolium Cav.) Dans Le Gouvernorat de Kairouan (Tunisie)*. EPPO Bull. 2007, 37, 132–136. [Google Scholar] [CrossRef]
- Mekki, M. Potential Threat of Solanum elaeagnifolium Cav. To the Tunisian Fields. In Invasive Plants in Mediterranean Type Regions of the World. Mèze, France, 2005; Brunel, S., Ed.; Council of Europe Publishing: Strasbourg, France, 2006; pp. 170–175. [Google Scholar]
- Mekki, M. Biology, Distribution and Impacts of Silverleaf nightshade (Solanum elaeagnifolium Cav.)*. EPPO Bull. 2007, 37, 114–118. [Google Scholar] [CrossRef]
- Sayari, N.; Brundu, G.; Mekki, M. Mapping and Monitoring an Invasive Alien Plant in Tunisia: Silverleaf nightshade (Solanum elaeagnifolium) a Noxious Weed of Agricultural Areas. Tunis. J. Plant Prot. 2016, 11, 219–227. [Google Scholar]
- Wassermann, V.D.; Zimmermann, H.G.; Neser, S. The Weed Silverleaf Bitter Apple (“Satansbos”) (Solanum elaeagnifolium Cav.) with Special Reference to Its Status in South Africa. South Afr. Dep. Agric. Water Supply Tech. Commun. 1988. Available online: https://eurekamag.com/research/007/945/007945510.php (accessed on 1 July 2022).
- Kidston, J.; Thompson, R.; Johnson, A. Silverleaf nightshade. New South Wales: Primefact 237. 2007. Available online: http://www.dpi.nsw.gov.au/__data/assets/pdf_file/0008/90890/silverleaf-nightshade.pdf (accessed on 1 July 2022).
- Douh, B.; Mguidiche, A.; Jar Allah al-Marri, M.; Moussa, M.; Rjeb, H. Assessment of Deficit Irrigation Impact on Agronomic Parameters and Water Use Efficiency of Six Chickpea (Cicer arietinum L.) Cultivars under Mediterranean Semi-Arid Climate. Ital. J. Agrometeorol. 2021, 2, 29–42. [Google Scholar] [CrossRef]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S-PLUS; Springer Science & Business Media: Berlin, Germany, 2013. [Google Scholar]
- Wickham, H.; Chang, W.; Wickham, M.H. Package ‘Ggplot2.’ Create elegant data visualisations using the grammar of graphics. Version 2016, 2, 1–189. [Google Scholar]
- Stanton, R.; Wu, H.; Lemerle, D. Root Regenerative Ability of Silverleaf nightshade (‘Solanum elaeagnifolium’ Cav.) in the Glasshouse. Plant Protection Quarterly 2011, 26, 54–56. [Google Scholar]
- Fernández, O.; Brevedan, R. Regeneracion de Solanum elaeagnifolium Cav. a Partir de Fragmentos de Sus Raices. Darwiniana 1972, 433–442. [Google Scholar]
- Wang, Y.-J.; Müller-Schärer, H.; van Kleunen, M.; Cai, A.-M.; Zhang, P.; Yan, R.; Dong, B.-C.; Yu, F.-H. Invasive Alien Plants Benefit More from Clonal Integration in Heterogeneous Environments than Natives. New Phytol. 2017, 216, 1072–1078. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Dong, M.; Miao, S.L.; Li, Z.Y.; Song, M.H.; Wang, R.Q. Invasive Alien Plants in China: Role of Clonality and Geographical Origin. Biol. Invasions 2006, 8, 1461–1470. [Google Scholar] [CrossRef]
- Caron, V.; Ede, F.J.; Sunnucks, P. Unravelling the Paradox of Loss of Genetic Variation during Invasion: Superclones May Explain the Success of a Clonal Invader. PLoS ONE 2014, 9, e97744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heap, J.; Carter, R. The Biology of Australian Weeds: 35. Solanum elaeagnifolium Cav. Plant Prot. Q. 1999, 14, 2–12. [Google Scholar]
- Ghabrit, S.B.; Bouhache, M.; Birouk, A.; BON, C. Variation Intraspécifique Du Potentiel Invasif de La Morelle Jaune (Solanum elaeagnifolium Cav.) Au Maroc: Influence Des Conditions Écologiques. Rev. Maroc. Prot. Plantes 2020, 14, 1–18. [Google Scholar]
- Baye, Y.; Ameur, A.; Bouhache, M.; Taleb, A. Strategie de Lutte Chimique Contre La Morelle Jaune (Solanum elaeagnifolium Cav.) Au Maroc*. EPPO Bull. 2007, 37, 145–152. [Google Scholar] [CrossRef]
- Baye, Y. Influence de Certains Facteurs Sur l’efficacité Du Glyphosate et de l’aminotriazole Contre Solanum elaeagnifolium Cav.*. EPPO Bull. 2007, 37, 153–155. [Google Scholar] [CrossRef]
- Wu, H.; Stanton, R.; Lemerle, D. Herbicidal Control of Solanum elaeagnifolium Cav. in Australia. Crop Prot. 2016, 88, 58–64. [Google Scholar] [CrossRef]
- Bouhache, M.; Boulet, C.; El Karakhi, F. Evolution Des Hydrates de Carbone Non Structuraux Chez La Morelle Jaune (Solatium elaeagnifolium Cav.). Weed Res. 1993, 33, 291–298. [Google Scholar] [CrossRef]
- Chavana, J.; Singh, S.; Vazquez, A.; Christoffersen, B.; Racelis, A.; Kariyat, R.R. Local Adaptation to Continuous Mowing Makes the Noxious Weed Solanum elaeagnifolium a Superweed Candidate by Improving Fitness and Defense Traits. Sci. Rep. 2021, 11, 6634. [Google Scholar] [CrossRef]
- Hulme, P.E.; Liu, W. Species Prevalence and Plant Traits Discriminate between Herbicide Resistant and Susceptible Weeds. Pest Manag. Sci. 2022, 78, 313–320. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Jan | Feb | Mar | Apr | May | Jun | Jul | Ago | Sep | Oct | Nov | Dec | Tot |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 2011 | 32.5 | 35.7 | 40.5 | 32.4 | 60.0 | 31.5 | 0.0 | 0.0 | 0.0 | 135.5 | 67.0 | 20.5 | 455.6 |
| 2012 | 32.0 | 55.2 | 80.1 | 15.0 | 7.2 | 0.0 | 0.0 | 6.5 | 81.5 | 8.0 | 0.0 | 0.0 | 285.5 |
| 2013 | 14.0 | 19.0 | 44.5 | 8.0 | 0.0 | 0.0 | 0.0 | 8.0 | 23.5 | 29.5 | 27.0 | 94.7 | 268.2 |
| 2014 | 6.3 | 5.4 | 9.7 | 2.7 | 2.7 | 3.9 | 0.0 | 0.1 | 18.7 | 3.5 | 13.4 | 41.7 | 108.1 |
| 2015 | 0.0 | 40.0 | 39.5 | 0.0 | 0.0 | 0.0 | 3.0 | 79.0 | 33.0 | 39.0 | 40.0 | 22.3 | 295.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sayari, N.; Brundu, G.; Soilhi, Z.; Mekki, M. Solanum elaeagnifolium Invasiveness under Semi-Arid Environmental Conditions in Tunisia. Earth 2022, 3, 1076-1086. https://doi.org/10.3390/earth3040062
Sayari N, Brundu G, Soilhi Z, Mekki M. Solanum elaeagnifolium Invasiveness under Semi-Arid Environmental Conditions in Tunisia. Earth. 2022; 3(4):1076-1086. https://doi.org/10.3390/earth3040062
Chicago/Turabian StyleSayari, Najla, Giuseppe Brundu, Zayneb Soilhi, and Mounir Mekki. 2022. "Solanum elaeagnifolium Invasiveness under Semi-Arid Environmental Conditions in Tunisia" Earth 3, no. 4: 1076-1086. https://doi.org/10.3390/earth3040062