
Directed Evolution of a Genetically Encoded Bioluminescent Ca2+ Sensor †


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Methods
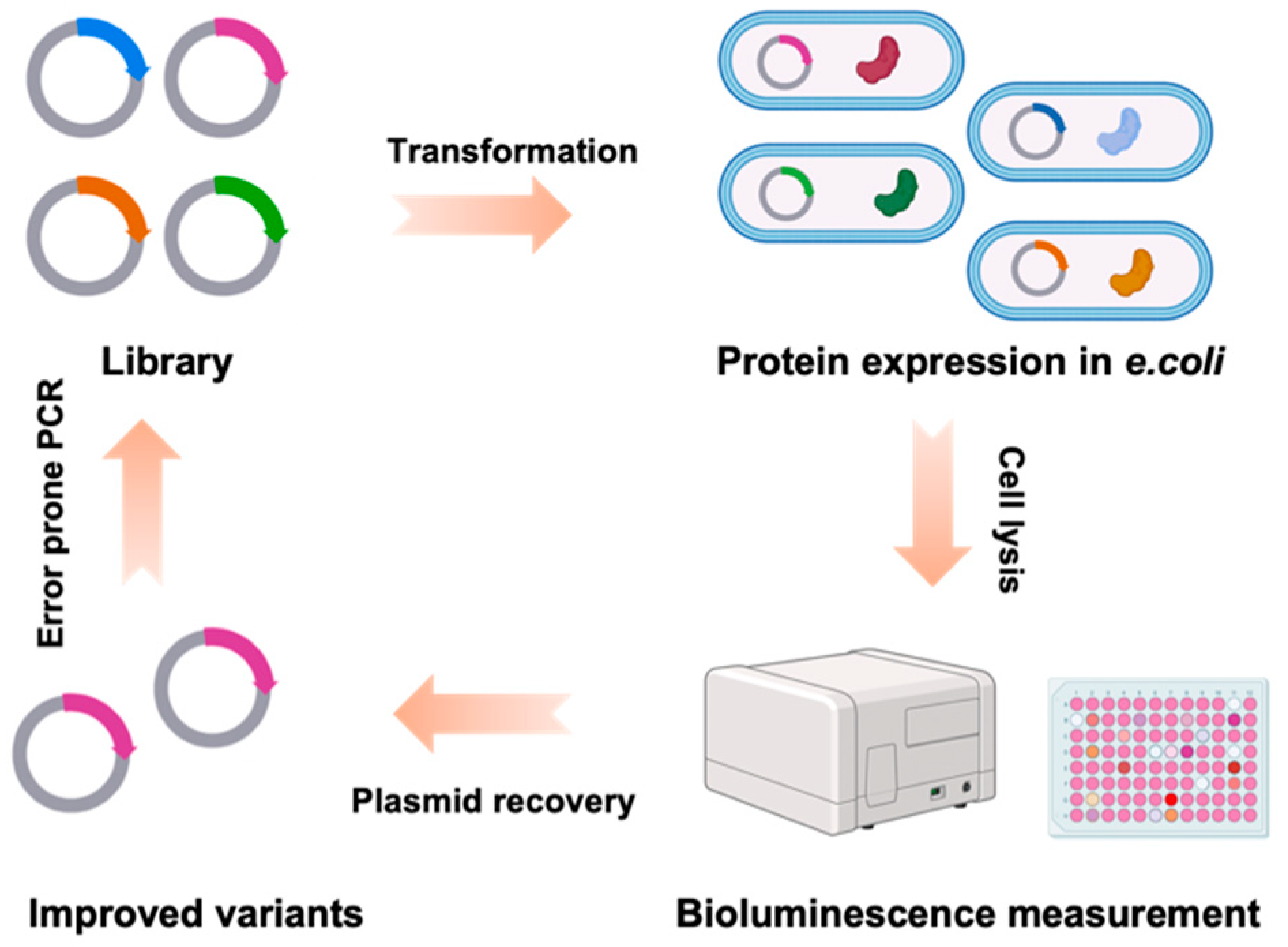
2.1. Directed Evolution of CaMBI
2.2. In Vitro Characterization
2.3. HeLa Cell Culture and Imaging
2.4. Culture and Imaging of Dissociated Neurons
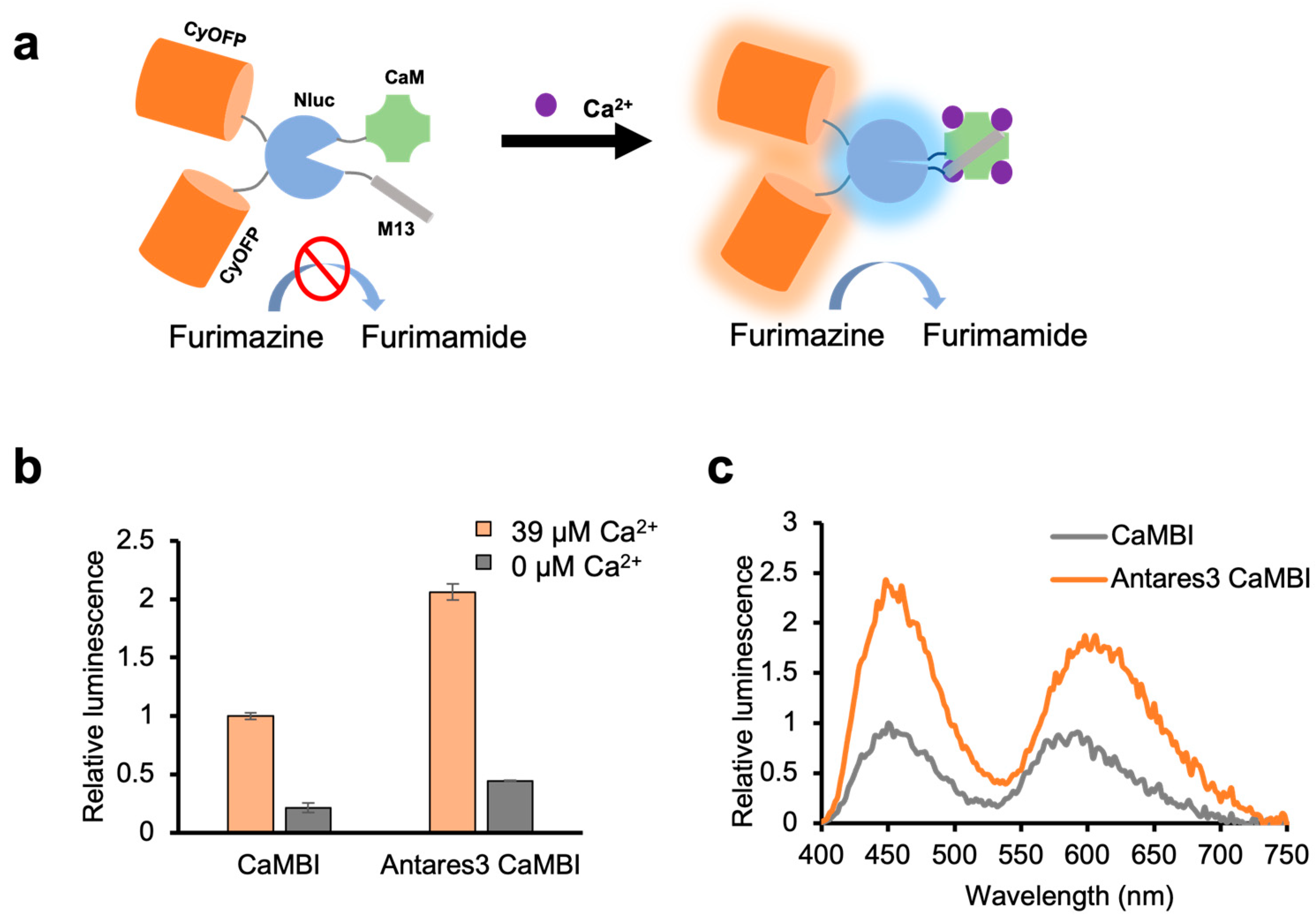
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, T.W.; Wardill, T.J.; Sun, Y.; Pulver, S.R.; Renninger, S.L.; Baohan, A.; Schreiter, E.R.; Kerr, R.A.; Orger, M.B.; Jayaraman, V.; et al. Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature 2013, 499, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Dana, H.; Sun, Y.; Mohar, B.; Hulse, B.K.; Kerlin, A.M.; Hasseman, J.P.; Tsegaye, G.; Tsang, A.; Wong, A.; Patel, R.; et al. High-performance calcium sensors for imaging activity in neuronal populations and microcompartments. Nat. Methods 2019, 16, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Oh, Y.; Park, Y.; Cho, J.; Wu, H.; Paulk, N.K.; Liu, L.X.; Kim, N.; Kay, M.A.; Wu, J.C.; Lin, M.Z. An orange calcium-modulated bioluminescent indicator for non-invasive activity imaging. Nat. Chem. Biol. 2019, 15, 433–436. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Zhang, Y.; Li, X.; Xiong, Y.; Wu, T.; Ai, H.W. A luciferase prosubstrate and a red bioluminescent calcium indicator for imaging neuronal activity in mice. Nat. Commun. 2022, 13, 3967. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, K.; Kimura, T.; Shinoda, H.; Bai, G.; Daniels, M.J.; Arai, Y.; Nakano, M.; Nagai, T. Five colour variants of bright luminescent protein for real-time multicolour bioimaging. Nat. Commun. 2016, 7, 13718. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Cumberbatch, D.; Centanni, S.; Shi, S.-Q.; Winder, D.; Webb, D.; Johnson, C.H. Coupling optogenetic stimulation with NanoLuc-based luminescence (BRET) Ca++ sensing. Nat. Commun. 2016, 7, 13268. [Google Scholar] [CrossRef] [PubMed]
- Farhana, I.; Hossain, M.N.; Suzuki, K.; Matsuda, T.; Nagai, T. Genetically encoded fluorescence/bioluminescence bimodal indicators for Ca2+ imaging. ACS Sens. 2019, 4, 1825–1834. [Google Scholar] [CrossRef] [PubMed]
- Su, Y.; Walker, J.R.; Hall, M.P.; Klein, M.A.; Wu, X.; Encell, L.P.; Casey, K.M.; Liu, L.X.; Hong, G.; Lin, M.Z.; et al. An optimized bioluminescent substrate for non-invasive imaging in the brain. Nat. Chem. Biol. 2023, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Villette, V.; Chavarha, M.; Dimov, I.K.; Bradley, J.; Pradhan, L.; Mathieu, B.; Evans, S.W.; Chamberland, S.; Shi, D.; Yang, R.; et al. Ultrafast Two-Photon Imaging of a High-Gain Voltage Indicator in Awake Behaving Mice. Cell 2019, 179, 1590–1608.e23. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, Y.; Lee, S.; Campbell, R.E.; Lin, M.Z. Directed Evolution of a Genetically Encoded Bioluminescent Ca2+ Sensor. Eng. Proc. 2023, 35, 20. https://doi.org/10.3390/IECB2023-14563
Zhao Y, Lee S, Campbell RE, Lin MZ. Directed Evolution of a Genetically Encoded Bioluminescent Ca2+ Sensor. Engineering Proceedings. 2023; 35(1):20. https://doi.org/10.3390/IECB2023-14563
Chicago/Turabian StyleZhao, Yufeng, Sungmoo Lee, Robert E. Campbell, and Michael Z. Lin. 2023. "Directed Evolution of a Genetically Encoded Bioluminescent Ca2+ Sensor" Engineering Proceedings 35, no. 1: 20. https://doi.org/10.3390/IECB2023-14563