
Eugenol Ester Derivatives: Synthesis, Insecticidal Activity and Computational Studies †


 , , ,
, , ,  ,
,  ,
,
Abstract
:1. Introduction
2. Results and Discussion
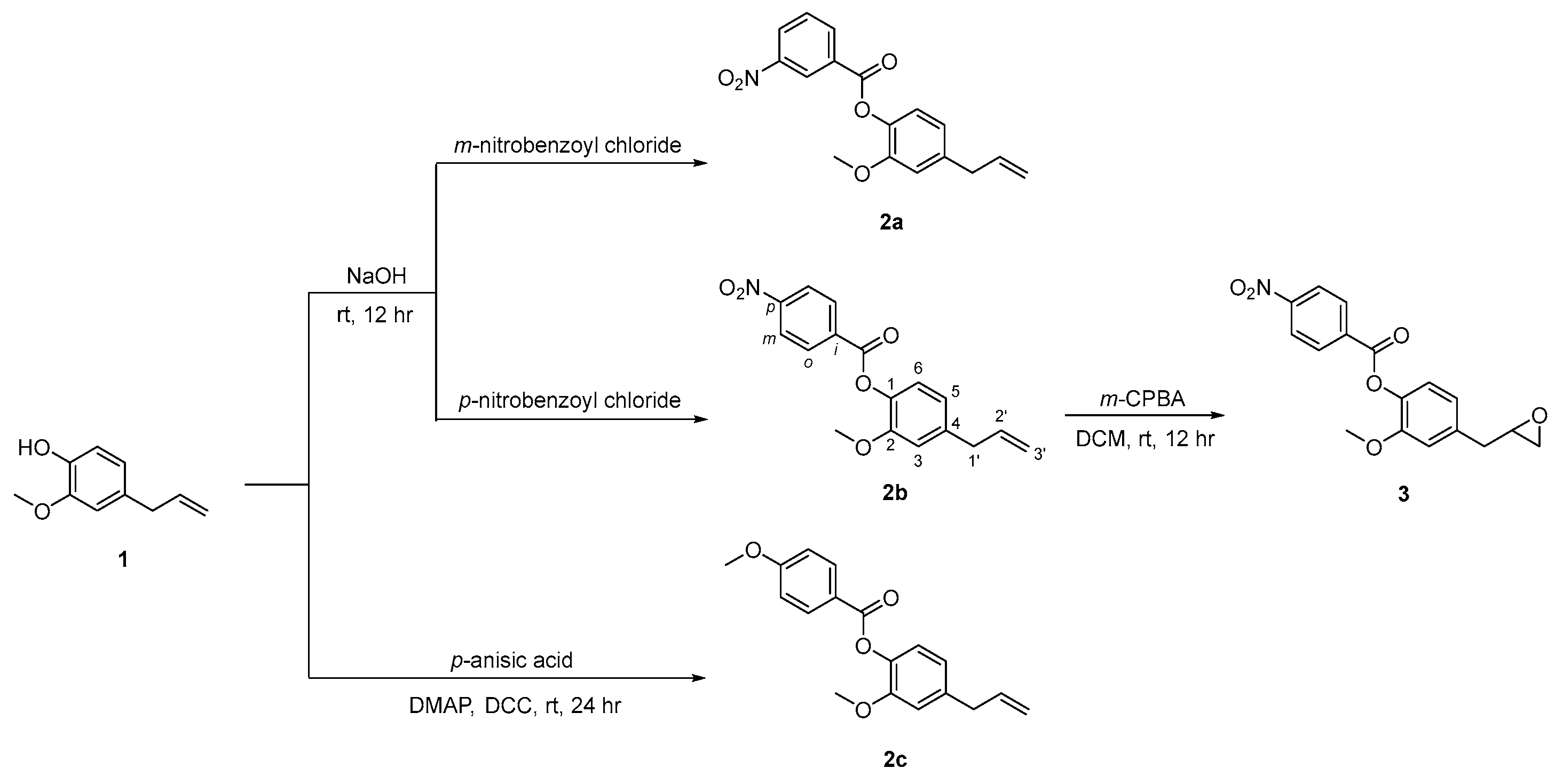
2.1. Synthesis of Eugenol Derivatives 2a–c and 3
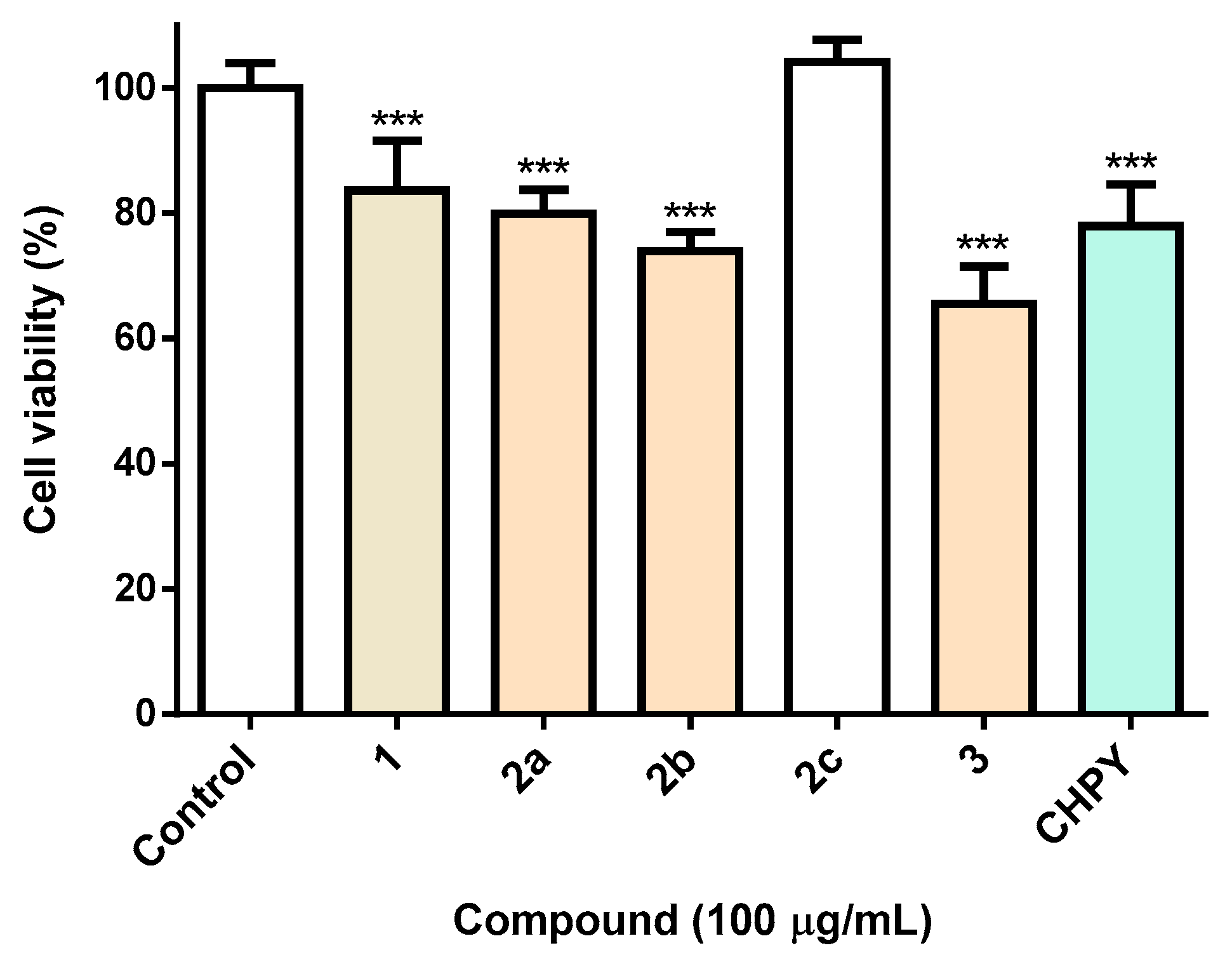
2.2. Biological Activity of Compounds 2a–c and 3 in Sf9 Insect Cells
2.3. Inverted Virtual Screening Results
2.4. Molecular Dynamics Simulations and Free Energy Calculations Results
3. Materials and Methods
3.1. Typical Procedure for the Preparation of Compounds 2a–c (Illustrated for 2b)
3.2. Synthesis of Compound 2c
3.3. Synthesis of Compound 3
3.4. Evaluation of Viability in Sf9 Cells
3.5. Inverted Virtual Screening Protocol Optimization
3.6. Molecular Dynamics Simulations and Free Energy Calculations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lengai, G.M.W.; Muthomi, J.W.; Mbega, E.R. Phytochemical activity and role of botanical pesticides in pest management for sustainable agricultural crop production. Sci. Afr. 2020, 7, e00239. [Google Scholar] [CrossRef]
- Oliveira, J.L.; Campos, E.V.R.; Fraceto, L.F. Recent developments and challenges for nanoscale formulation of botanical pesticides for use in sustainable agriculture. J. Agric. Food Chem. 2018, 66, 8898–8913. [Google Scholar] [CrossRef] [PubMed]
- Salman, M.; Abbas, R.Z.; Israr, M.; Abbas, A.; Mehmood, K.; Khan, M.K.; Sindhu, Z.D.; Hussaind, R.; Saleemie, M.K.; Shaha, S. Repellent and acaricidal activity of essential oils and their components against Rhipicephalus ticks in cattle. Vet. Parasitol. 2020, 283, 109178. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.Y. Essential oils as repellents against arthropods. Biomed. Res. Int. 2018, 2018, 6860271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, M.J.G.; Pereira, R.B.; Pereira, D.M.; Fortes, A.G.; Castanheira, E.M.S.; Gonçalves, M.S.T. New eugenol derivatives with enhanced insecticidal activity. Int. J. Mol. Sci. 2020, 21, 9257. [Google Scholar] [CrossRef]
- Ju, J.; Xie, Y.; Yu, H.; Guo, Y.; Cheng, Y.; Qian, H.; Yao, W. Analysis of the synergistic antifungal mechanism of eugenol and citral. LWT Food Sci. Technol. 2020, 123, 109128. [Google Scholar] [CrossRef]
- Spinelli, S.; Lagarde, A.; Iovinella, I.; Legrand, P.; Tegoni, M.; Pelosi, P.; Cambillau, C. Crystal structure of Apis mellifera OBP14, a C-minus odorant-binding protein, and its complexes with odorant molecules. Insect Biochem. Mol. Biol. 2012, 42, 41–50. [Google Scholar] [CrossRef]
- Ramos, R.S.; Costa, J.S.; Silva, R.C.; Costa, G.V.; Rodrigues, A.B.L.; Rabelo, E.M.; Souto, R.N.P.; Taft, C.A.; Silva, C.H.T.P.; Rosa, J.M.C.; et al. Identification of potential inhibitors from Pyriproxyfen with insecticidal activity by virtual screening. Pharmaceuticals 2019, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Riva, C.; Suzanne, P.; Charpentier, G.; Dulin, F.; Halm-Lemeille, M.-P.; Santos, J.S.-O. In silico chemical library screening and experimental validation of novel compounds with potential varroacide activities. Pestic. Biochem. Physiol. 2019, 160, 11–19. [Google Scholar] [CrossRef]
- Correy, G.J.; Zaidman, D.; Harmelin, A.; Carvalho, S.; Mabbitta, P.D.; Calaora, V.; Peter, J.; James, P.J.; Kotzeg, A.C.; Jackson, C.J.; et al. Overcoming insecticide resistance through computational inhibitor design. Proc. Natl. Acad. Sci. USA 2019, 116, 21012–21021. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Liu, M.; Yao, Y.; Wang, J.; Li, Y.; Li, G.; Wang, Y. Identification of novel potential β-N-Acetyl-D-Hexosaminidase inhibitors by virtual screening, molecular dynamics simulation and MM-PBSA calculations. Int. J. Mol. Sci. 2012, 13, 4545–4563. [Google Scholar] [CrossRef] [PubMed]
- Dong, L.; Shen, S.; Xu, Y.; Wang, L.; Yang, Q.; Zhang, J.; Lu, H. Identification of novel insect β-N-acetylhexosaminidase OfHex1 inhibitors based on virtual screening, biological evaluation, and molecular dynamics simulation. J. Biomol. Struct. Dyn. 2021, 39, 1735–1743. [Google Scholar] [CrossRef]
- Dong, Y.; Jiang, X.; Liu, T.; Ling, Y.; Yang, Q.; Zhang, L.; He, X. Structure-based virtual screening, compound synthesis, and bioassay for the design of chitinase inhibitors. J. Agric. Food Chem. 2018, 66, 3351–3357. [Google Scholar] [CrossRef] [PubMed]
- Hu, X.; Yin, B.; Cappelle, K.; Swevers, L.; Smagghe, G.; Yang, X.; Zhang, L. Identification of novel agonists and antagonists of the ecdysone receptor by virtual screening. J. Mol. Graph Model. 2018, 81, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Nakagawa, Y.; Ogura, T.; Yamada, Y.; Ohe, T.; Miyagawa, H. Virtual screening for ligands of the insect molting hormone receptor. J. Chem. Inf. Model. 2011, 51, 296–305. [Google Scholar] [CrossRef]
- Min, J.; Lin, D.; Zhang, Q.; Zhang, J.; Yu, Z. Structure-based virtual screening of novel inhibitors of the uridyltransferase activity of Xanthomonas oryzae pv. oryzae GlmU. Eur. J. Med. Chem. 2012, 53, 150–158. [Google Scholar] [CrossRef]
- Offermann, L.R.; Chan, S.L.; Osinski, T.; Tan, Y.W.; Chew, F.T.; Sivaraman, J.; Mok, Y.-K.; Minor, W.; Chruszcz, M. The major cockroach allergen Bla g 4 binds tyramine and octopamine. Mol. Immunol. 2014, 60, 86–94. [Google Scholar] [CrossRef] [Green Version]
- Laughlin, J.D.; Ha, T.S.; Jones, D.N.M.; Smith, D.P. Activation of pheromone-sensitive neurons is mediated by conformational activation of pheromone-binding protein. Cell 2008, 133, 1255–1265. [Google Scholar] [CrossRef] [Green Version]
- Oliferenko, P.V.; Oliferenko, A.A.; Poda, G.I.; Osolodkin, D.I.; Pillai, G.G.; Bernier, U.R.; Tsikolia, M.; Agramonte, N.M.; Clark, G.G.; Linthicum, K.J.; et al. Promising aedes aegypti repellent chemotypes identified through integrated QSAR, virtual ccreening, synthesis, and bioassay. PLoS ONE 2013, 8, e64547. [Google Scholar] [CrossRef]
- Joshi, T.; Joshi, T.; Sharma, P.; Chandra, S.; Pande, V. Molecular docking and molecular dynamics simulation approach to screen natural compounds for inhibition of Xanthomonas oryzae pv. Oryzae by targeting peptide deformylase. J. Biomol. Struct. Dyn. 2021, 39, 823–840. [Google Scholar] [CrossRef]
- Fu, Y.; Liu, Y.-X.; Kang, T.; Sun, Y.-N.; Li, J.-Z.; Ye, F. Identification of novel inhibitors of p-hydroxyphenylpyruvate dioxygenase using receptor-based virtual screening. J. Taiwan Inst. Chem. Eng. 2019, 103, 33–43. [Google Scholar] [CrossRef]
- Fattouch, S.; Raboudi-Fattouch, F.; Ponce, J.V.G.; Forment, J.V.; Lukovic, D.; Marzouki, N.; Vidal, D.R. Concentration dependent effects of commonly used pesticides on activation versus inhibition of the quince (Cydonia Oblonga) polyphenol oxidase. Food Chem. Toxicol. 2010, 48, 957–963. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Du, X.; Wang, C.; Lin, J.; Du, X. Identification of Potential Helicoverpa armigera (Lepidoptera: Noctuidae) Sterol Carrier Protein-2 Inhibitors through High-Throughput Virtual Screening. J. Econ. Entomol. 2017, 110, 1779–1784. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Li, Z.; Jiang, Y.; Pan, X.; Wu, J.; Cristofori-Armstrong, B.; Smith, J.J.; Chin, Y.K.Y.; Lei, J.; Zhou, Q.; et al. Structural basis for the modulation of voltage-gated sodium channels by animal toxins. Science 2018, 362, eaau2596. [Google Scholar] [CrossRef] [Green Version]
- Berman, H.M.; Westbrook, J.; Feng, Z.; Gilliland, G.; Bhat, T.N.; Weissig, H.; Shindyalov, I.N.; Bourne, P.E. The protein data bank. Nucleic Acids Res. 2000, 28, 235–242. [Google Scholar] [CrossRef] [Green Version]
- Seeliger, D.; de Groot, B.L. Ligand docking and binding site analysis with PyMOL and Autodock/Vina. J. Comput. Aided Mol. Des. 2010, 24, 417–422. [Google Scholar] [CrossRef] [Green Version]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2009, 31, 455–461. [Google Scholar] [CrossRef] [Green Version]
- Jones, G.; Willett, P.; Glen, R.C.; Leach, A.R.; Taylor, R. Development and validation of a genetic algorithm for flexible docking Edited by F. E. Cohen. J. Mol. Biol. 1997, 267, 727–748. [Google Scholar] [CrossRef] [Green Version]
- Sander, T.; Freyss, J.; von Korff, M.; Rufener, C. DataWarrior: An open-source program for chemistry aware data visualization and analysis. J. Chem. Inf. Model. 2015, 55, 460–473. [Google Scholar] [CrossRef]
- O’Boyle, N.M.; Banck, M.; James, C.A.; Morley, C.; Vandermeersch, T.; Hutchison, G.R. Open babel: An open chemical toolbox. J. Cheminform. 2011, 3, 33. [Google Scholar] [CrossRef] [Green Version]
- Case, D.A.; Cheatham, T.E., III; Darden, T.; Gohlke, H.; Luo, R.; Merz, K.M., Jr.; Onufriev, A.; Simmerling, C.; Wang, B.; Woods, R.J. The Amber biomolecular simulation programs. J. Comput. Chem. 2005, 26, 1668–1688. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and testing of a general amber force field. J. Comp. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef] [PubMed]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. ff14SB: Improving the accuracy of protein side chain and backbone parameters from ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [Green Version]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian16; Gaussian, Inc.: Wallingford, CT, USA, 2016. [Google Scholar]
- Wang, J.; Wang, W.; Kollman, P.A.; Case, D.A. Automatic atom type and bond type perception in molecular mechanical calculations. J. Mol. Graph Model. 2006, 25, 247–260. [Google Scholar] [CrossRef] [PubMed]
- Roe, D.R.; Cheatham, T.E. PTRAJ and CPPTRAJ: Software for processing and analysis of molecular dynamics trajectory data. J. Chem. Theory Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Wang, E.; Sun, H.; Wang, J.; Wang, Z.; Liu, H.; Zhang, J.Z.H.; Hou, T. End-point binding free energy calculation with MM/PBSA and MM/GBSA: Strategies and applications in drug design. Chem. Rev. 2019, 119, 9478–9508. [Google Scholar] [CrossRef]
- Miller, B.R.; McGee, T.D.; Swails, J.M.; Homeyer, N.; Gohlke, H.; Roitberg, A.E. MMPBSA.py: An efficient program for end-state free energy calculations. J. Chem. Theory Comput. 2012, 8, 3314–3321. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Target | PDB | PLP | ASP | ChemScore | GoldScore | Vina | Overall Ranking |
|---|---|---|---|---|---|---|---|
| Acetylcholinesterase | 1QON | 76.71 | 56.76 | 38.14 | 61.43 | −9.63 | 2 |
| 4EY6 | 76.16 | 51.57 | 38.88 | 57.98 | −9.00 | ||
| 1DX4 | 76.32 | 50.62 | 35.14 | 63.56 | −9.33 | ||
| alpha-Esterase-7 (αE7) | 5TYJ | 62.07 | 36.72 | 28.51 | 55.37 | −7.33 | 7 |
| 5TYP | 63.49 | 40.84 | 31.42 | 55.85 | −7.15 | ||
| beta-N-Acetyl-D-hexosaminidase OfHex1 | 3NSN | 75.83 | 54.25 | 32.29 | 58.32 | −7.63 | 4 |
| 3OZP | 70.09 | 50.77 | 30.95 | 61.58 | −8.55 | ||
| Chitinase | 3WL1 | 74.64 | 48.90 | 33.49 | 61.49 | −8.28 | 3 |
| 3WQV | 74.20 | 47.85 | 33.40 | 64.67 | −8.55 | ||
| Ecdysone receptor | 1R20 | 71.10 | 32.79 | 32.22 | 56.77 | −8.03 | 5 |
| 1R1K | 69.64 | 35.64 | 34.15 | 60.76 | −8.78 | ||
| N-Acetylglucosamine-1-phosphate uridyltransferase (GlmU) | 2V0K | 54.77 | 25.78 | 23.49 | 53.69 | −7.20 | 12 |
| 2VD4 | 47.41 | 26.34 | 22.46 | 42.93 | −5.98 | ||
| Octopamine receptor | 4N7C | 47.39 | 33.06 | 27.26 | 47.53 | −5.90 | 13 |
| Odorant Binding Protein | 5V13 | 84.80 | 52.54 | 40.54 | 65.30 | −9.13 | 1 |
| 2GTE | 65.44 | 37.13 | 36.20 | 61.29 | −8.15 | ||
| 3N7H | 80.79 | 44.86 | 30.52 | 69.46 | −7.45 | ||
| 3K1E | 85.76 | 46.38 | 35.88 | 71.78 | −7.20 | ||
| Peptide deformylase | 5CY8 | 69.29 | 32.36 | 24.45 | 61.43 | −7.93 | 8 |
| p-Hydroxyphenylpyruvate dioxygenase | 6ISD | 63.44 | 38.44 | 28.09 | 52.91 | −8.10 | 9 |
| Polyphenol oxidase | 3HSS | 54.54 | 29.27 | 24.58 | 64.34 | −6.75 | 10 |
| Sterol carrier protein-2 (HaSCP-2) | 4UEI | 65.99 | 34.95 | 31.54 | 52.99 | −8.25 | 6 |
| Voltage-gated sodium channel | 6A95 | 61.46 | 25.01 | 23.35 | 58.99 | −7.33 | 11 |
| Average RMSD of the Complex (Å) | Average RMSD of the Ligand (Å) | Average SASA (Å2) | Percentage of Potential Ligand SASA Buried (%) | Average Number of Hbonds | ΔGbind (kcal/mol) | Main Contributors (kcal/mol) | ||
|---|---|---|---|---|---|---|---|---|
| OBP | 2a | 2.0 ± 0.2 | 0.8 ± 0.2 | 69.2 ± 15.2 | 87 | 0.01 ± 0.05 | −37.7 ± 0.1 | Trp105 (−3.2 ± 0.6); Leu67 (−2.2 ± 0.4); Met82 (−1.5 ± 0.4) |
| 2b | 2.2 ± 0.2 | 0.8 ± 0.2 | 70.2 ± 12.9 | 87 | 0.01 ± 0.1 | −38.6 ± 0.1 | Trp105 (−3.1 ± 0.5); Leu67 (−1.7 ± 0.5); Ile78 (−1.7 ± 0.6) | |
| 2c | 2.2 ± 0.2 | 0.6 ± 0.1 | 59.6 ± 13.5 | 89 | 0.01 ± 0.1 | −37.2 ± 0.1 | Trp105 (−2.6 ± 0.5); Leu67 (−1.9 ± 0.4); Ile78 (−1.6 ± 0.6) | |
| 3 | 2.0 ± 0.1 | 0.8 ± 0.3 | 59.6 ± 13.5 | 89 | 0.01 ± 0.1 | −39.7 ± 0.1 | Trp105 (−3.2 ± 0.5); Leu67 (−1.9 ± 0.4); Ile78 (−1.8 ± 0.5) | |
| AChE | 2a | 2.9 ± 0.2 | 0.9 ± 0.2 | 39.1 ± 9.9 | 93 | 0.01 ± 0.3 | −25.4 ± 0.1 | Tyr69 (−1.4 ±0.6); Gly148 (−1.2 ± 0.5); Tyr322 (−1.0 ± 0.5) |
| 2b | 2.9 ± 0.4 | 1.3 ± 0.2 | 68.2 ± 21.2 | 88 | 0.3 ± 0.5 | −29.2 ± 0.2 | Tyr372 (−3.0 ± 0.8); Trp81 (−2.0 ± 0.9); Tyr69 (−1.6 ± 0.5) | |
| 2c | 2.9 ± 0.4 | 0.9 ± 0.2 | 51.1 ± 12.8 | 90 | 0.7 ± 0.9 | −27.3 ± 0.2 | Trp81 (−2.3 ± 0.5); Tyr69 (−1.7 ± 1.0); Tyr368 (−1.6 ± 1.1) | |
| 3 | 3.6 ± 0.3 | 1.0 ± 0.3 | 39.3 ± 13.1 | 93 | 0.1 ± 0.3 | −31.7 ± 0.2 | Trp81 (−2.7 ± 0.5): Gly148 (−1.2 ± 0.5); Tyr372 (−1.2 ± 0.4) |
| Target | Organism | PDB Target | Resolution (Å) | Ref. |
|---|---|---|---|---|
| Acetylcholinesterase | Aedes aegypti | 1QON | 2.72 | [8] |
| 4EY6 | 2.40 | |||
| Drosophila melanogaster | 1DX4 | 2.70 | [9] | |
| alpha-Esterase-7 (αE7) | Lucilia cuprina | 5TYJ | 1.75 | [10] |
| 5TYP | 1.88 | |||
| beta-N-Acetyl-D-hexosaminidase OfHex1 | Ostrinia furnacalis | 3NSN | 2.10 | [11] |
| 3OZP | 2.00 | [12] | ||
| Chitinase | Ostrinia furnacalis | 3WL1 | 1.77 | [13] |
| 3WQV | 2.04 | |||
| Ecdysone receptor | Heliothis virescens | 1R20 | 3 | [14] |
| 1R1K | 2.9 | [15] | ||
| N-Acetylglucosamine-1-phosphate uridyltransferase (GlmU) | Xanthomonas oryzae | 2V0K | 2.3 | [16] |
| 2VD4 | 1.9 | |||
| Octopamine receptor | Blattella germanica | 4N7C | 1.75 | [17] |
| Odorant Binding Protein | Aedes aegypti | 5V13 | 1.84 | [8] |
| Drosophila melanogaster | 2GTE | 1.4 | [18] | |
| Anopheles gambiae | 3N7H | 1.6 | [19] | |
| Aedes aegypti | 3K1E | 1.85 | ||
| Peptide deformylase | Xanthomonas oryzae | 5CY8 | 2.38 | [20] |
| p-Hydroxyphenylpyruvate dioxygenase | Arabidopsis thaliana | 6ISD | 2.4 | [21] |
| Polyphenol oxidase | Manduca sexta | 3HSS | 2.7 | [22] |
| Sterol carrier protein-2 (HaSCP-2) | Helicoverpa armigera | 4UEI | Solution NMR | [23] |
| Voltage-gated sodium channel | Periplaneta americana | 6A95 | 2.6 | [24] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Coelho, J.R.A.; Vieira, T.F.; Pereira, R.B.; Pereira, D.M.; Castanheira, E.M.S.; Fortes, A.G.; Sousa, S.F.; Fernandes, M.J.G.; Gonçalves, M.S.T. Eugenol Ester Derivatives: Synthesis, Insecticidal Activity and Computational Studies. Chem. Proc. 2022, 8, 83. https://doi.org/10.3390/ecsoc-25-11787
Coelho JRA, Vieira TF, Pereira RB, Pereira DM, Castanheira EMS, Fortes AG, Sousa SF, Fernandes MJG, Gonçalves MST. Eugenol Ester Derivatives: Synthesis, Insecticidal Activity and Computational Studies. Chemistry Proceedings. 2022; 8(1):83. https://doi.org/10.3390/ecsoc-25-11787
Chicago/Turabian StyleCoelho, José R. A., Tatiana F. Vieira, Renato B. Pereira, David M. Pereira, Elisabete M. S. Castanheira, A. Gil Fortes, Sérgio F. Sousa, Maria José G. Fernandes, and M. Sameiro T. Gonçalves. 2022. "Eugenol Ester Derivatives: Synthesis, Insecticidal Activity and Computational Studies" Chemistry Proceedings 8, no. 1: 83. https://doi.org/10.3390/ecsoc-25-11787