
Analysis of Acetylcholinesterase Activity in Cell Membrane Microarrays of Brain Areas as a Screening Tool to Identify Tissue Specific Inhibitors

 , , and
, , and
Abstract
:
1. Introduction
2. Experimental Section
2.1. Tissue Samples and Reagents
2.2. Cell Membrane Microarrays Fabrication
2.3. ChE Activity Detection
2.4. Data Normalization
2.5. Image Processing
2.6. Data Analysis and Determination of IC50 Values
3. Results and Discussion
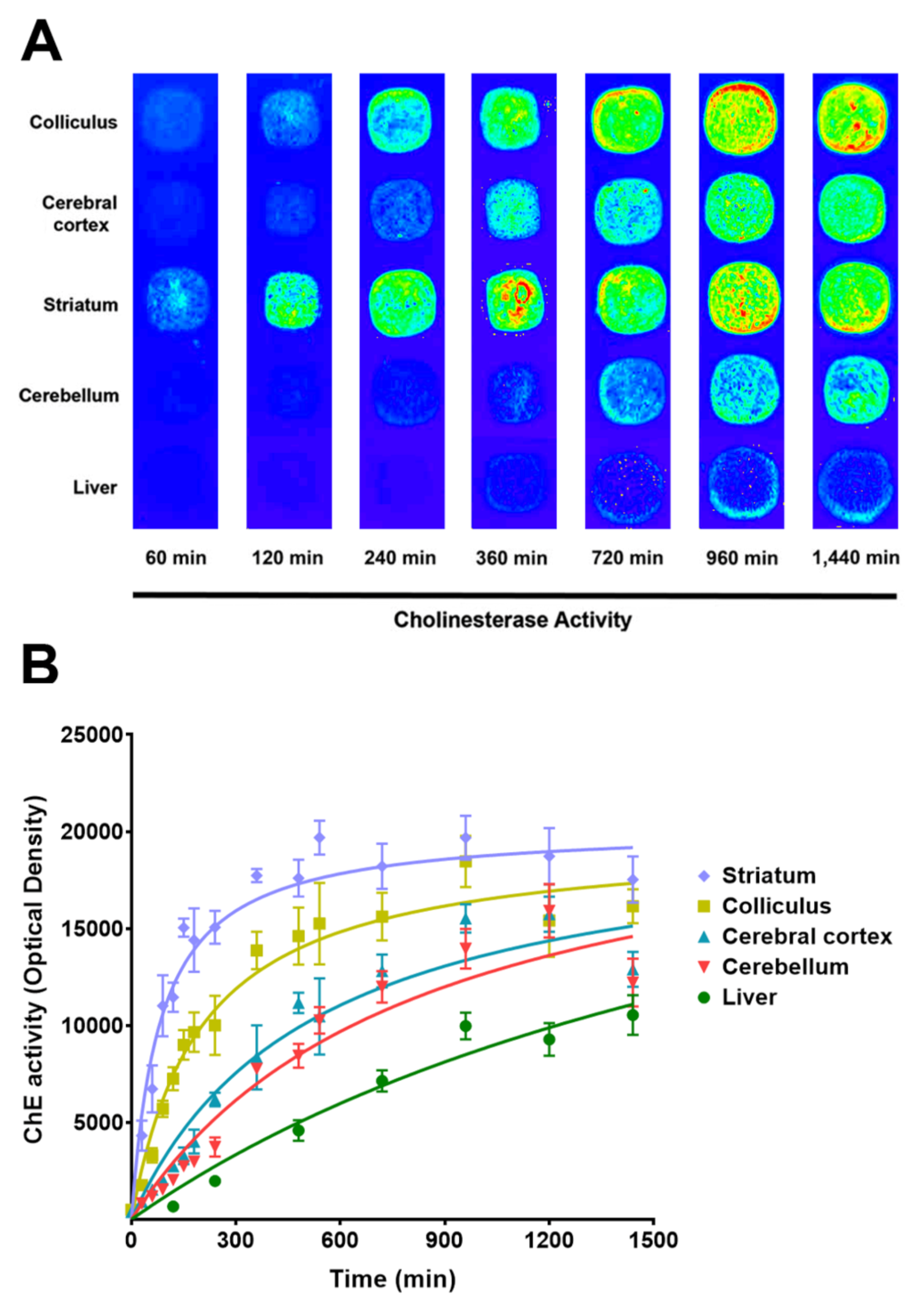
3.1. Protocol Optimization to Detect ChE and AChE Activity in Cell Membrane Microarrays
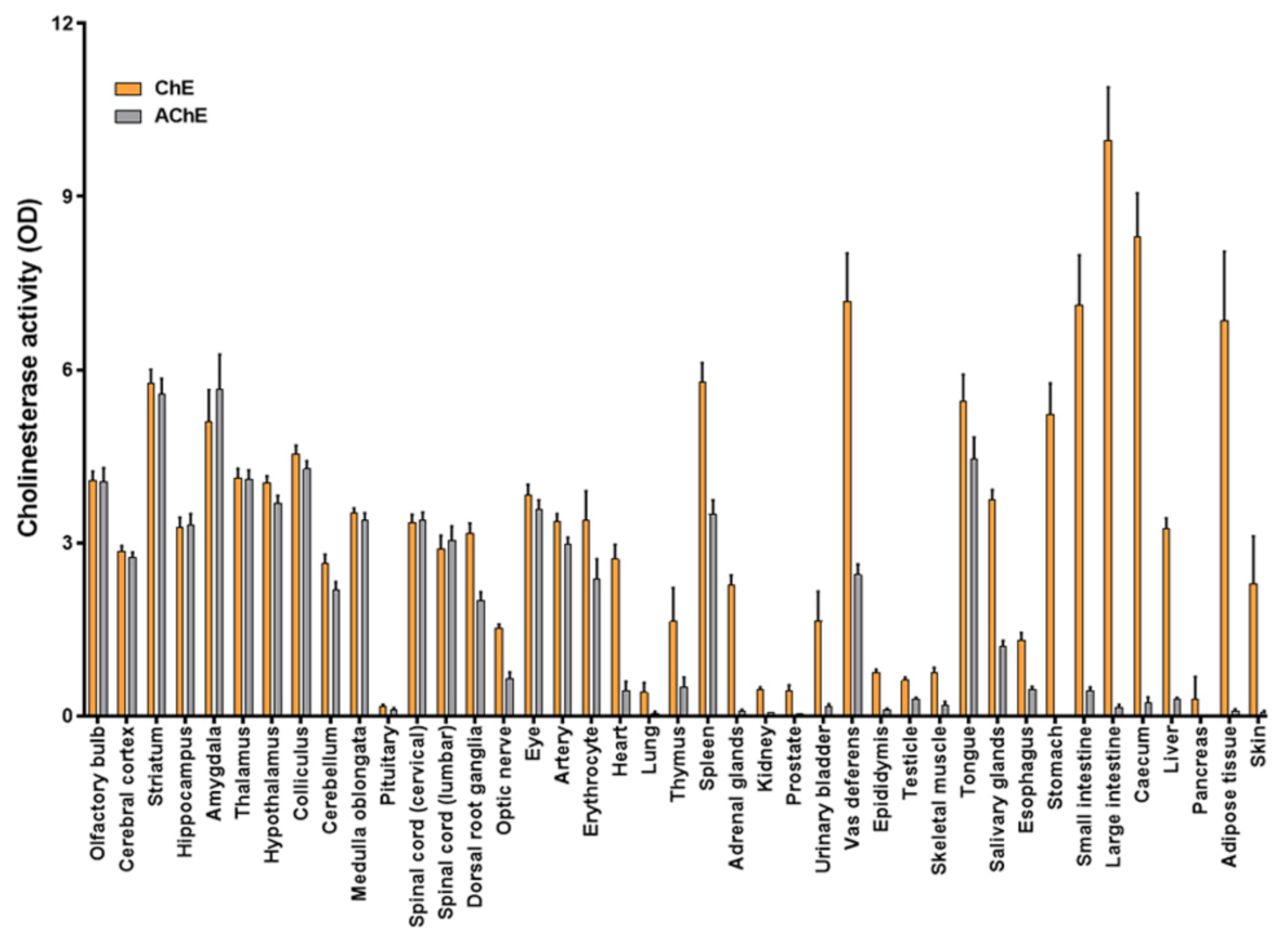
3.2. Determination of ChE and AChE Activities in a Collection of Rat Tissues Using CMMs
3.3. Effects of Donepezil on AChE Activity Using CMMs
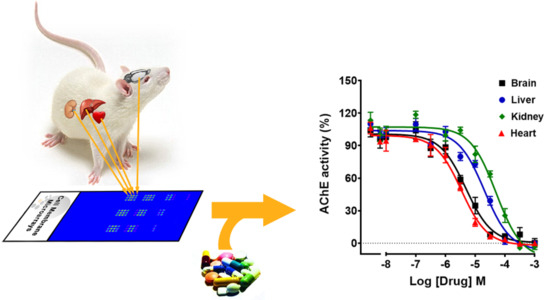
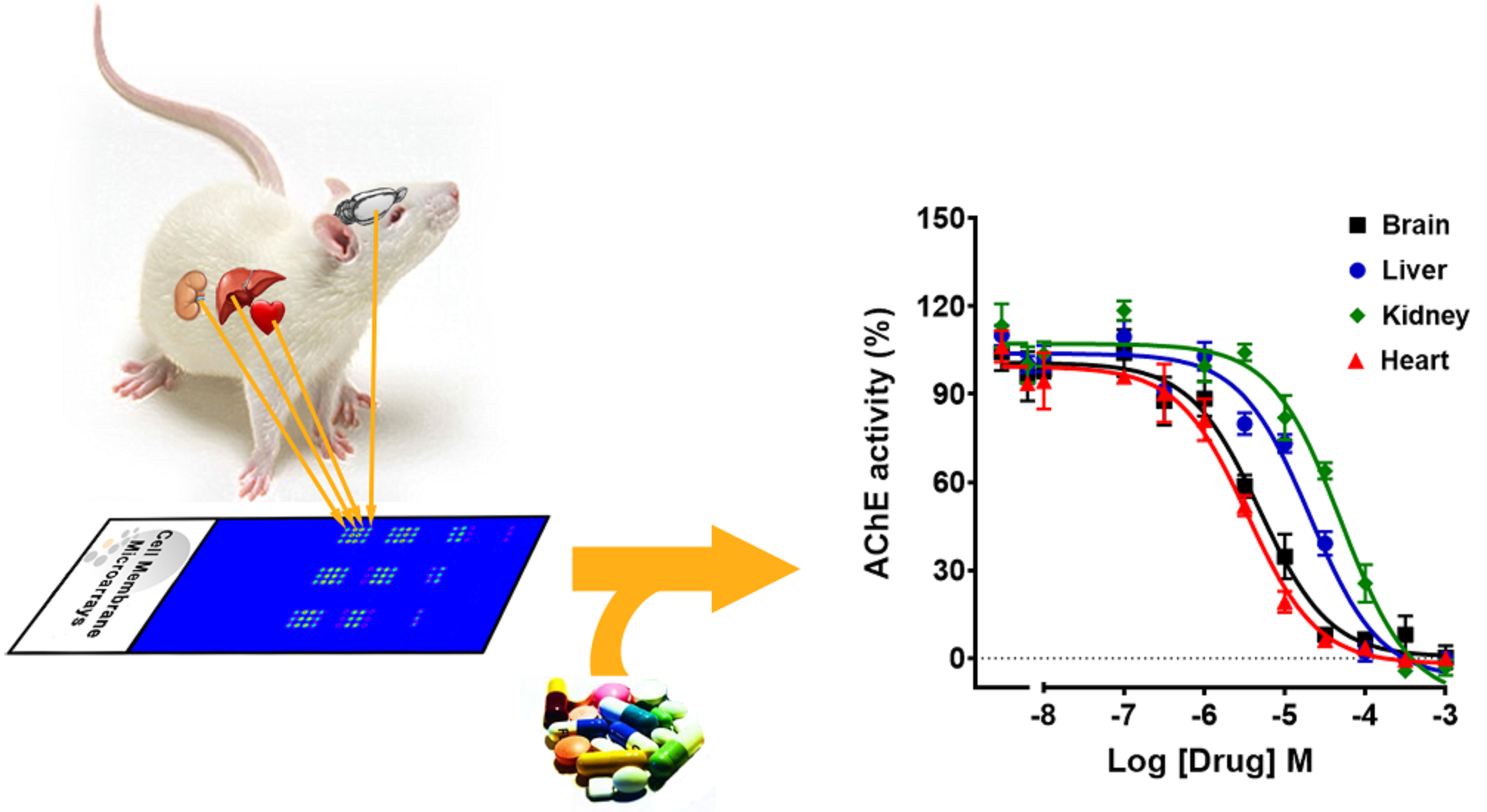
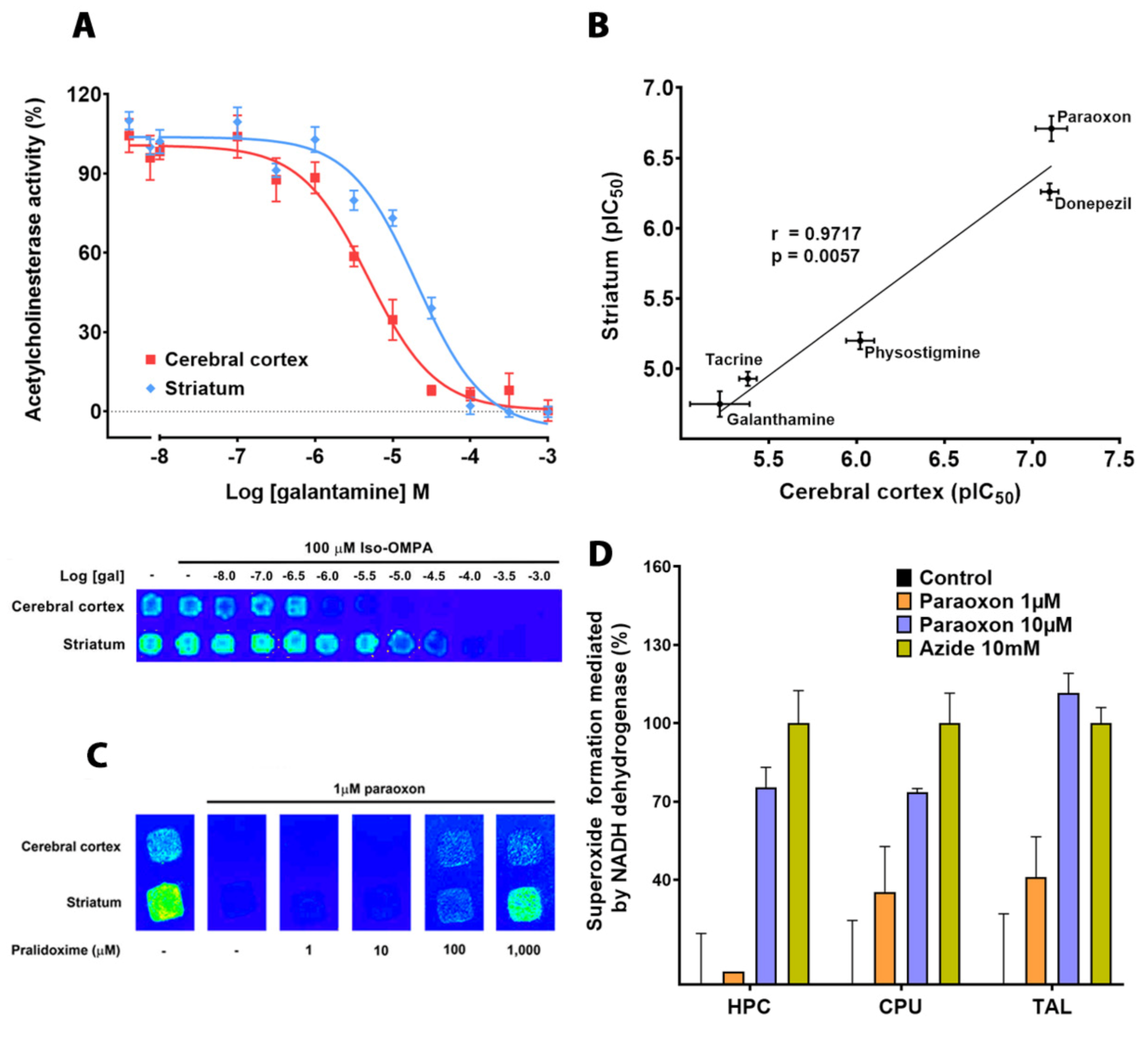
3.4. Inhibition of AChE Activity by Different Drugs Using Striatal and Cortical CMMs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bartus, R.T.; Dean, R.L., 3rd; Beer, B.; Lippa, A.S. The cholinergic hypothesis of geriatric memory dysfunction. Science 1982, 217, 408–414. [Google Scholar] [CrossRef]
- Perry, E.K.; Curtis, M.; Dick, D.J.; Candy, J.M.; Atack, J.R.; Bloxham, C.A.; Blessed, G.; Fairbairn, A.; Tomlinson, B.E.; Perry, R.H. Cholinergic correlates of cognitive impairment in Parkinson’s disease: Comparisons with Alzheimer’s disease. J. Neurol. Neurosurg. Psychiatry 1985, 48, 413–421. [Google Scholar] [CrossRef] [Green Version]
- Pohanka, M. Cholinesterases, a target of pharmacology and toxicology. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc Czech Repub. 2011, 155, 219–229. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Gu, X.; Zhang, G.; Zhang, D.; Zhu, D. Continuous colorimetric assay for acetylcholinesterase and inhibitor screening with gold nanoparticles. Langmuir 2009, 25, 2504–2507. [Google Scholar] [CrossRef]
- Li, Y.; Bai, H.; Li, C.; Shi, G. Colorimetric Assays for Acetylcholinesterase Activity and Inhibitor Screening Based on the Disassembly—Assembly of a Water-Soluble Polythiophene Derivative. ACS Appl. Matter Interfaces 2011, 3, 1306–1310. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Feather-Stone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Hay, D.L.; Ibrahim, G.F.; Horacek, I. Rapid acetylcholinesterase screening test for neural tube defect. Clin. Chem. 1983, 29, 1065–1069. [Google Scholar] [CrossRef]
- Guilbault, G.G.; Kramer, D.N. Fluorometric system employing immobilized cholinesterase for assaying anticholinesterase compounds. Anal. Chem. 1965, 37, 1675–1680. [Google Scholar] [CrossRef]
- Wang, M.; Gu, X.; Zhang, G.; Zhang, D.; Zhu, D. Convenient and continuous fluorometric assay method for acetylcholinesterase and inhibitor screening based on the aggregation-induced emission. Anal. Chem. 2009, 81, 4444–4449. [Google Scholar] [CrossRef]
- Shen, X.; Liang, F.; Zhang, G.; Zhang, D. A new continuous fluorometric assay for acetylcholinesterase activity and inhibitor screening with emissive core–shell silica particles containing tetraphenylethylene fluorophore. Analyst 2012, 137, 2119–2123. [Google Scholar] [CrossRef]
- Liao, D.; Chen, J.; Zhou, H.; Wang, Y.; Li, Y.; Yu, C. In situ formation of metal coordination polymer: A strategy for fluorescence turn-on assay of acetylcholinesterase activity and inhibitor screening. Anal. Chem. 2013, 85, 2667–2672. [Google Scholar] [CrossRef]
- Winteringham, F.P.; Disney, R.W. Radiometric assay of acetylcholinesterase. Nature 1962, 195, 1303. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, T.; Okamura, T.; Arai, T.; Obeta, T.; Fukushi, K.; Irie, T.; Shiraishi, T. Use of a novel radiometric method to assess the inhibitory effect of donepezil on acetylcholinesterase activity in minimally diluted tissue samples. Br. J. Pharmacol. 2010, 159, 1732–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Potter, P.E.; Meek, J.L.; Neff, N.H. Acetylcholine and choline in neuronal tissue measured by HPLC with electrochemical detection. J. Neurochem. 1983, 41, 188–194. [Google Scholar] [CrossRef]
- Durand, P.; Thomas, D. Use of immobilized enzyme coupled with an electrochemical sensor for the detection of organophosphates and carbamates pesticides. J. Environ. Pathol. Toxicol. Oncol. 1984, 5, 51–57. [Google Scholar] [PubMed]
- Du, D.; Tao, Y.; Zhang, W.; Liu, D.; Li, H. Oxidative desorption of thiocholine assembled on core–shell Fe3O4/AuNPs magnetic nanocomposites for highly sensitive determination of acetylcholinesterase activity: An exposure biomarker of organophosphates. Biosens. Bioelectron. 2011, 26, 4231–4235. [Google Scholar] [CrossRef]
- Cuartero, M.; Ortuño, J.A.; García, M.S.; García-Cánovas, F. Assay of acetylcholinesterase activity by potentiometric monitoring of acetylcholine. Anal. Biochem. 2012, 421, 208–212. [Google Scholar] [CrossRef]
- Monton, M.R.; Lebert, J.M.; Little, J.R.; Nair, J.J.; McNulty, J.; Brennan, J.D. A Sol− Gel-Derived Acetylcholinesterase Microarray for Nanovolume Small-Molecule Screening. Anal. Chem. 2010, 82, 9365–9373. [Google Scholar] [CrossRef] [PubMed]
- Hicks, D.; John, D.; Makova, N.Z.; Henderson, Z.; Nalivaeva, N.N.; Turner, A.J. Membrane targeting, shedding and protein interactions of brain acetylcholinesterase. J. Neurochem. 2011, 116, 742–746. [Google Scholar] [CrossRef]
- Manuel, I.; Barreda-Gómez, G.; González de San Román, E.; Veloso, A.; Fernández, J.A.; Giralt, M.T.; Rodríguez-Puertas, R. Neurotransmitter receptor localization: From autoradiography to imaging mass spectrometry. ACS Chem. Neurosci. 2015, 6, 362–373. [Google Scholar] [CrossRef]
- Hebert-Chatelain, E.; Desprez, T.; Serrat, R.; Bellocchio, L.; Soria-Gomez, E.; Busquets-Garcia, A.; Pagano Zottola, A.C.; Delamarre, A.; Cannich, A.; Vincent, P.; et al. A cannabinoid link between mitochondria and memory. Nature 2016, 539, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Fernández, R.; Garate, J.; Tolentino-Cortez, T.; Herraiz, A.; Lombardero, L.; Ducrocq, F.; Rodríguez-Puertas, R.; Trifilieff, P.; Astigarraga, E.; Barreda-Gómez, G.; et al. Microarray and Mass Spectrometry-Based Methodology for Lipid Profiling of Tissues and Cell Cultures. Anal. Chem. 2019, 91, 15967–15973. [Google Scholar] [CrossRef] [PubMed]
- Pereiro, X.; Fernández, R.; Barreda-Gómez, G.; Ruzafa, N.; Acera, A.; Araiz, J.; Astigarraga, E.; Vecino, E. Comparative lipidomic analysis of mammalian retinal ganglion cells and Müller glia in situ and in vitro using High-Resolution Imaging Mass Spectrometry. Sci. Rep. 2020, 10, 20053. [Google Scholar] [CrossRef]
- Karnovsky, M.J.; Roots, L. A” direct-coloring” thiocholine method for cholinesterases. J. Histochem. Cytochem. 1964, 12, 219–221. [Google Scholar] [CrossRef]
- Glutekin, F.; Ozturk, M.; Akdogan, M. The effect of organophosphate insecticide chlorpyrifos-ethyl on lipid peroxidation and antioxidant enzymes (in vitro). Arch. Toxicol. 2000, 74, 533–538. [Google Scholar] [CrossRef]
- Gupta, J.; Datta, C.; Sarkar, A.; Sengupta, D. 1992. Effect of malathion on antioxidant defense system in human fetus—An in vitro study. Indian J. Exp. Biol. 1992, 30, 352–354. [Google Scholar] [PubMed]
- Prins, J.M.; Chao, C.K.; Jacobson, S.M.; Thompson, C.M.; George, K.M. Oxidative stress resulting from exposure of a human salivary gland cells to paraoxon: An in vitro model for organophosphate oral exposure. Toxicol In Vitro 2014, 28, 715–721. [Google Scholar] [CrossRef] [Green Version]
- Barreda-Gómez, G.; Giralt, M.T.; Rodríguez-Puertas, R. Methods to measure g-protein-coupled receptor activity for the identification of inverse agonists. Methods Enzymol. 2010, 485, 261–273. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Rodríguez Puertas, R.; Barreda Gómez, G.; Giralt Rué, M.T.; Fernández Pastor, B. Method of Quantifying the G Protein-Coupled Receptor (GPCR)/G Protein Coupling Using a Cell Membrane Array. Patent WO/2007/012688, 1 February 2007. [Google Scholar]
- Rodríguez Puertas, R.; Barreda Gómez, G.; Fernández González, J.A.; Astigarraga Arribas, E. Method for the Surface Treatment of Solid Substrates. Patent WO/2008/012391, 31 January 2008. [Google Scholar]
- Tago, H.; Kimura, H.; Maeda, T.J. Visualization of detailed acetylcholinesterase fiber and neuron staining in rat brain by a sensitive histochemical procedure. Histochem. Cytochem. 1986, 34, 1431–1438. [Google Scholar] [CrossRef]
- Shih, T.M. Comparison of several oximes on reactivation of soman-inhibited blood, brain and tissue cholinesterase activity in rats. Arch. Toxicol. 1993, 67, 637–646. [Google Scholar] [CrossRef]
- Mack, A.; Robitzki, A. The key role of butyrylcholinesterase during neurogenesis and neural disorders: An antisense-5′ butyrylcholinesterase-DNA study. Prog. Neurobiol. 2000, 60, 607–628. [Google Scholar] [CrossRef]
- Lassiter, T.L.; Marshall, R.S.; Jackson, L.C.; Hunter, D.L.; Vu, J.T.; Padilla, S. Automated measurement of acetylcholinesterase activity in rat peripheral tissues. Toxicology 2003, 186, 241–253. [Google Scholar] [CrossRef]
- Ord, M.G.; Thompson, R.H. Pseudo-cholinesterase activity in the central nervous system. Biochem. J. 1952, 51, 245–251. [Google Scholar] [CrossRef] [Green Version]
- Milatovic, D.; Dettbarn, W.D. Modification of acetylcholinesterase during adaptation to chronic, subacute paraoxon application in rat. Toxicol. Appl. Pharmacol. 1996, 136, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Goh, C.W.; Aw, C.C.; Lee, J.H.; Chen, C.P.; Browne, E.R. Pharmacokinetic and pharmacodynamic properties of cholinesterase inhibitors donepezil, tacrine, and galantamine in aged and young Lister hooded rats. Drug Metab. Dispos. 2011, 39, 402–411. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards, J.A.; Brimijoin, S. Divergent regulation of acetylcholinesterase and butyrylcholinesterase in tissues of the rat. J. Neurochem. 1982, 38, 1393–1403. [Google Scholar] [CrossRef]
- Li, B.; Stribley, J.A.; Ticu, A.; Xie, W.; Schopfer, L.M.; Hammond, P.; Brimijoin, S.; Hinrichs, S.H.; Lockridge, O. Abundant tissue butyrylcholinesterase and its possible function in the acetylcholinesterase knockout mouse. J. Neurochem. 2000, 75, 1320–1331. [Google Scholar] [CrossRef]
- Das, A.; Dikshit, M.; Nath, C. Profile of acetylcholinesterase in brain areas of male and female rats of adult and old age. Life Sci. 2001, 68, 1545–1555. [Google Scholar] [CrossRef]
- Tecles, F.; Cerón, J.J. Determination of whole blood cholinesterase in different animal species using specific substrates. Res. Vet. Sci. 2001, 70, 233–238. [Google Scholar] [CrossRef]
- Grigoryan, H.; Halebyan, G.; Lefebvre, B.; Brasme, B.; Masson, P. Mechanism of hydrolysis of dicholine esters with long polymethylene chain by human butyrylcholinesterase. Biochim. Biophys. Acta 2008, 1784, 1818–1824. [Google Scholar] [CrossRef]
- Kita, T.; Hata, T.; Higashiguchi, T.; Itoh, E.; Kawabata, A. Changes of total acetylcholine content and the activity of related enzymes in SART (repeated cold)-stressed rat brain and duodenum. Jpn. J. Pharmacol. 1986, 40, 174–177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paxinos, G.; Watson, C.R.; Emson, P.C. AChE-stained horizontal sections of the rat brain in stereotaxic coordinates. J. Neurosci. Methods 1980, 3, 129–149. [Google Scholar] [CrossRef]
- Rakonczay, Z. Potencies and selectivities of inhibitors of acetylcholinesterase and its molecular forms in normal and Alzheimer’s disease brain. Acta Biol. Hung. 2003, 54, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Chen, V.P.; Luk, W.K.; Chan, W.K.; Leung, K.W.; Guo, A.J.; Chan, G.K.; Xu, S.L.; Choi, R.C.; Tsim, K.W. Molecular assembly and biosynthesis of acetylcholinesterase in brain and muscle: The roles of t-peptide, FHB domain, and N-linked glycosylation. Front. Mol. Neurosci. 2011, 4, 36. [Google Scholar] [CrossRef] [Green Version]
- Snape, M.F.; Misra, A.; Murray, T.K.; De Souza, R.J.; Williams, J.L.; Cross, A.J.; Green, A.R. A comparative study in rats of the in vitro and in vivo pharmacology of the acetylcholinesterase inhibitors tacrine, donepezil and NXX-066. Neuropharmacology 1999, 38, 181–193. [Google Scholar] [CrossRef]
- Ogura, H.; Kosasa, T.; Kuriya, Y.; Yamanishi, Y. Comparison of inhibitory activities of donepezil and other cholinesterase inhibitors on acetylcholinesterase and butyrylcholinesterase in vitro. Methods Find. Exp. Clin. Pharmacol. 2000, 8, 609–613. [Google Scholar] [CrossRef]
- Thomsen, T.; Kaden, B.; Fischer, J.P.; Bickel, U.; Barz, H.; Gusztony, G.; Cervos-Navarro, J.; Kewitz, H. Inhibition of acetylcholinesterase activity in human brain tissue and erythrocytes by galanthamine, physostigmine and tacrine. Eur. J. Clin. Chem. Clin. Biochem. 1991, 29, 487–492. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Q.; Tang, X.C. Effects of huperzine A on acetylcholinesterase isoforms in vitro: Comparison with tacrine, donepezil, rivastigmine and physostigmine. Eur. J. Pharmacol. 2002, 455, 101–107. [Google Scholar] [CrossRef]
- Geerts, H.; Guillaumat, P.O.; Grantham, C.; Bode, W.; Anciaux, K.; Sachak, S. Brain levels and acetylcholinesterase inhibition with galantamine and donepezil in rats, mice, and rabbits. Brain. Res. 2005, 1033, 186–193. [Google Scholar] [CrossRef]
- Albuquerque, E.X.; Pereira, E.F.; Aracava, Y.; Fawcett, W.P.; Oliviera, M.; Randall, W.R.; Hamilton, T.A.; Kan, R.K.; Romano, J.A., Jr.; Adler, M. Effective countermeasure against poisoning by organophosphorus insecticides and nerve agents. Proc. Natl. Acad. Sci. USA 2006, 103, 13220–13225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuca, K.; Hrabinova, M.; Soukup, O.; Tobin, G.; Karasova, J.; Pohanka, M. Pralidoxime—The gold standard of acetylcholinesterase reactivators—Reactivation in vitro efficacy. Bratisl. Lek Listy. 2010, 111, 502–504. [Google Scholar]
- Ferrari, F.; Villa, R.F. The Neurobiology of Depression: An Integrated Overview from Biological Theories to Clinical Evidence. Mol. Neurobiol. 2017, 54, 4847–4865. [Google Scholar] [CrossRef] [PubMed]
- Umeno, A.; Biju, V.; Yoshida, Y. In vivo ROS production and use of oxidative stress-derived biomarkers to detect the onset of diseases such as Alzheimer’s disease, Parkinson’s disease, and diabetes. Free Radic Res. 2017, 51, 413–427. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AChE Activity | AChE/ChE (%) | Log IC50 Donepezil | |
|---|---|---|---|
| Olfactory bulb | 4.06 ± 0.24 | 99 ± 7 | −6.76 ± 0.05 |
| Cerebral cortex | 2.75 ± 0.09 | 96 ± 4 | −7.10 ± 0.05 |
| Striatum | 5.57 ± 0.28 | 97 ± 6 | −6.26 ± 0.06 |
| Hippocampus | 3.32 ± 0.18 | 102 ± 8 | −6.87 ± 0.07 |
| Amygdala | 5.66 ± 0.61 | 111 ± 17 | −5.90 ± 0.03 |
| Thalamus | 4.10 ± 0.16 | 99 ± 5 | −6.56 ± 0.03 |
| Hypothalamus | 3.69 ± 0.13 | 91 ± 4 | −6.93 ± 0.03 |
| Colliculus | 4.28 ± 0.14 | 94 ± 4 | −6.87 ± 0.03 |
| Cerebellum | 2.19 ± 0.14 | 82 ± 7 | −7.20 ± 0.05 |
| Medulla oblongata | 3.40 ± 0.12 | 97 ± 4 | −6.85 ± 0.05 |
| Pituitary | 0.11 ± 0.03 | 69 ± 26 | ND |
| Spinal cord (cervical) | 3.05 ± 0.24 | 105 ± 12 | −6.72 ± 0.05 |
| Spinal cord (lumbar) | 3.39 ± 0.14 | 101 ± 6 | −6.91 ± 0.02 |
| Dorsal root ganglia | 2.00 ± 0.15 | 63 ± 6 | −7.28 ± 0.07 |
| Optic nerve | 0.65 ± 0.11 | 43 ± 7 | −7.25 ± 0.08 |
| Eye | 3.59 ± 0.15 | 93 ± 6 | −6.20 ± 0.06 |
| Artery | 2.98 ± 0.12 | 88 ± 5 | −7.12 ± 0.05 |
| Erythrocyte | 2.38 ± 0.34 | 70 ± 14 | −6.77 ± 0.10 |
| Heart | 0.45 ± 0.15 | 16 ± 6 | ND |
| Lung | 0.04 ± 0.04 | 10 ± 10 | ND |
| Thymus | 0.50 ± 0.17 | 31 ± 15 | ND |
| Spleen | 3.50 ± 0.24 | 61 ± 5 | −7.46 ± 0.06 |
| Adrenal glands | 0.08 ± 0.03 | 3 ± 1 | ND |
| Kidney | 0.06 ± 0.00 | 12 ± 1 | ND |
| Prostate | 0.05 ± 0.00 | 11 ± 2 | ND |
| Urinary bladder | 0.16 ± 0.05 | 9 ± 4 | ND |
| Vas deferens | 2.45 ± 0.18 | 34 ± 5 | −7.47 ± 0.09 |
| Epididymis | 0.11 ± 0.02 | 14 ± 3 | ND |
| Testicle | 0.30 ± 0.02 | 49 ± 5 | ND |
| Skeletal muscle | 0.20 ± 0.05 | 26 ± 8 | ND |
| Tongue | 4.46 ± 0.37 | 82 ± 10 | −7.42 ± 0.06 |
| Salivary glands | 1.20 ± 0.11 | 32 ± 3 | −7.49 ± 0.10 |
| Esophagus | 0.46 ± 0.05 | 35 ± 5 | ND |
| Stomach | ND | ND | ND |
| Small intestine | 0.44 ± 0.06 | 6 ± 1 | ND |
| Large intestine | 0.15 ± 0.05 | 2 ± 1 | ND |
| Caecum | 0.24 ± 0.09 | 3 ± 1 | ND |
| Liver | 0.30 ± 0.02 | 9 ± 1 | ND |
| Pancreas | ND | ND | ND |
| Adipose tissue | 0.08 ± 0.04 | 1 ± 1 | ND |
| Skin | 0.06 ± 0.03 | 3 ± 2 | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rienda, B.; Elexpe, A.; Tolentino-Cortez, T.; Gulak, M.; Bruzos-Cidón, C.; Torrecilla, M.; Astigarraga, E.; Barreda-Gómez, G. Analysis of Acetylcholinesterase Activity in Cell Membrane Microarrays of Brain Areas as a Screening Tool to Identify Tissue Specific Inhibitors. Analytica 2021, 2, 25-36. https://doi.org/10.3390/analytica2010003
Rienda B, Elexpe A, Tolentino-Cortez T, Gulak M, Bruzos-Cidón C, Torrecilla M, Astigarraga E, Barreda-Gómez G. Analysis of Acetylcholinesterase Activity in Cell Membrane Microarrays of Brain Areas as a Screening Tool to Identify Tissue Specific Inhibitors. Analytica. 2021; 2(1):25-36. https://doi.org/10.3390/analytica2010003
Chicago/Turabian StyleRienda, Bárbara, Ane Elexpe, Tarson Tolentino-Cortez, Marina Gulak, Cristina Bruzos-Cidón, María Torrecilla, Egoitz Astigarraga, and Gabriel Barreda-Gómez. 2021. "Analysis of Acetylcholinesterase Activity in Cell Membrane Microarrays of Brain Areas as a Screening Tool to Identify Tissue Specific Inhibitors" Analytica 2, no. 1: 25-36. https://doi.org/10.3390/analytica2010003