
Glycyrrhizinic Acid and Phosphatidylcholine Combination as a Preventive Therapy for Experimental Murine Non-Alcoholic Steatohepatitis
 , and
, and
Abstract
:1. Introduction
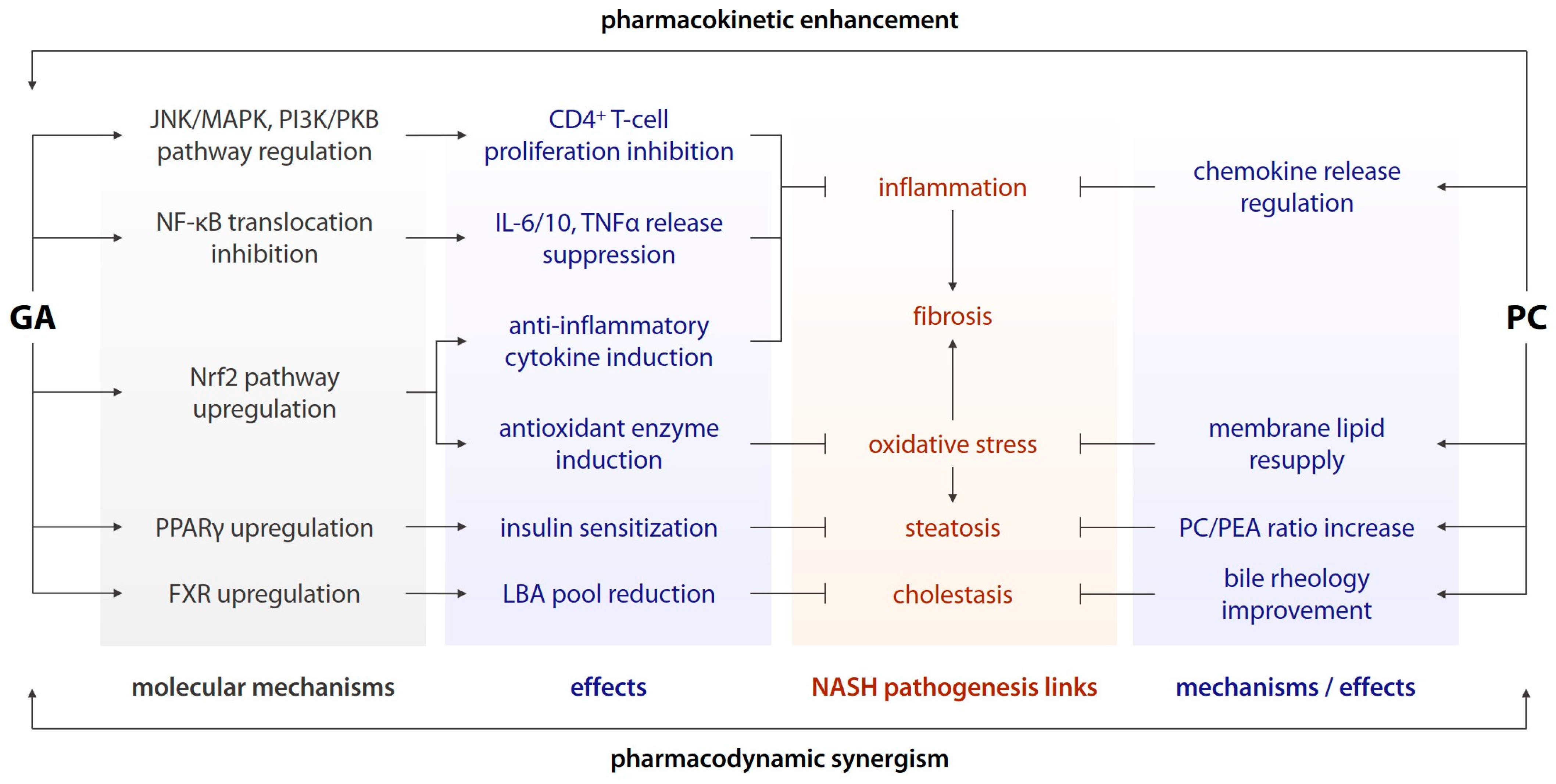
- Inhibition of nuclear factor-κB (NF-κB) translocation to the cell nucleus and the subsequent pro-inflammatory shift in gene expression [12];
- Suppression of release of interleukins (IL) 6 and 10, (TNFα), and other pro-inflammatory cytokines downstream of NF-κB [12];
- Inhibition of CD4+ T-cell proliferation via the c-Jun N-terminal kinase (JNK)/mitogen-activated protein kinase (MAPK) and phosphoinositide 3-kinase (PI3K)/protein kinase B (PKB/AKT) pathways [12];
- Upregulation of the nuclear factor erythroid 2-related factor 2 (Nrf2) signalling and its downstream effectors, including antioxidant enzymes and anti-inflammatory messengers [13];
- Upregulation of nuclear peroxisome proliferator-activated receptor γ (PPARγ) [14];
- Modulation of the sirtuin deacetylase expression and activity [15], and other effects;
2. Materials and Methods
- 1.
- Intact (n = 10), standard diet + 0.1 mL almond oil intraperitoneally (i/p) once weekly (q.wk.);
- 2.
- Control (n = 30), high-fat diet (HFD) + 0.32 mg/kg CCl4 i/p q.wk;
- 3.
- Glycyrrhizinic acid (GA) (n = 30), HFD + 34.3 mg/kg/d GA p/o + 0.32 mg/kg CCl4 i/p q.wk;
- 4.
- Glycyrrhizinic acid + phosphatidylcholine (GAPC) (n = 30), HFD + 34.3 mg/kg/d GA p/o + 158.1 mg/kg PC p/o + 0.32 mg/kg CCl4 i/p q.wk.
3. Results
3.1. Tissue Morphology
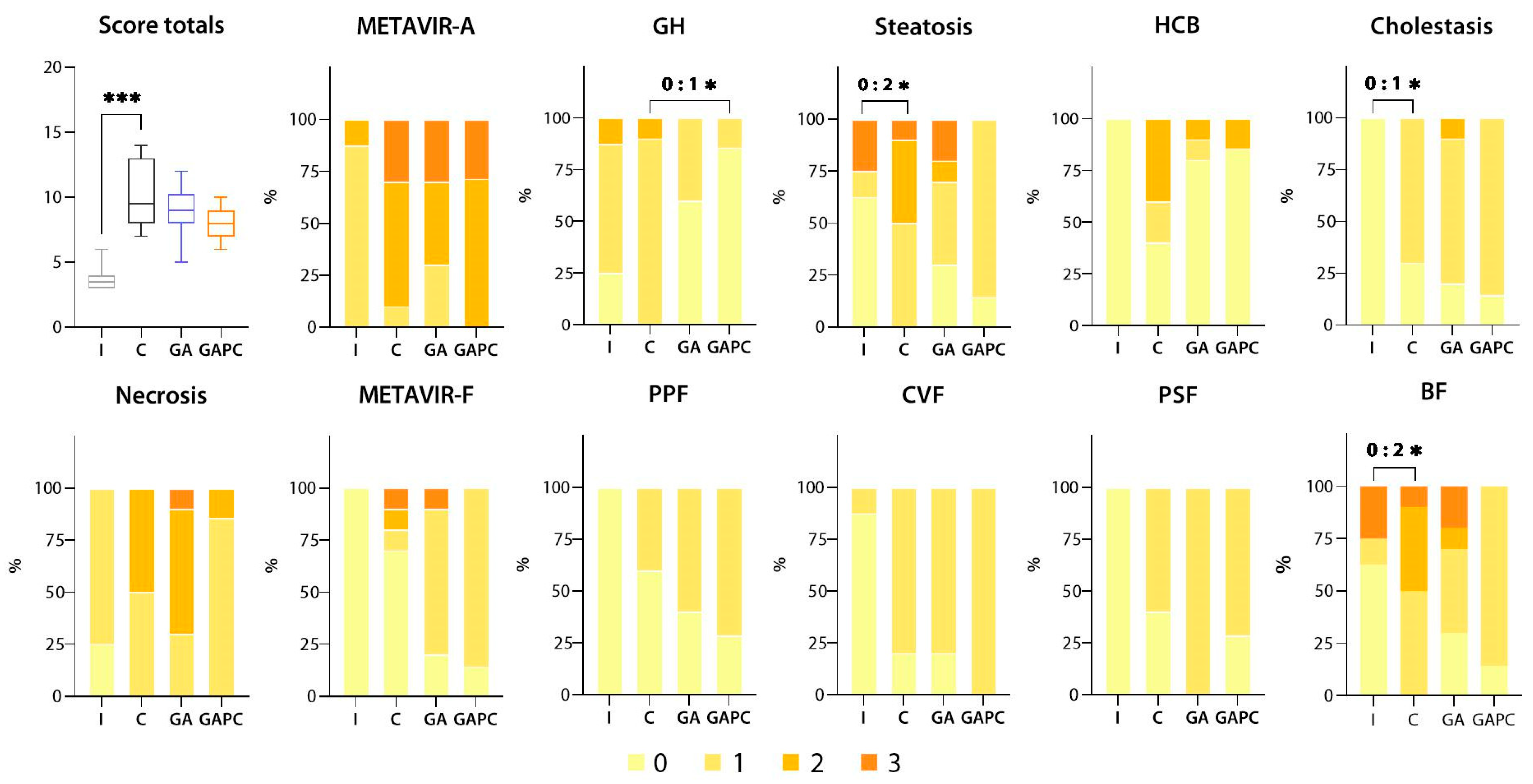
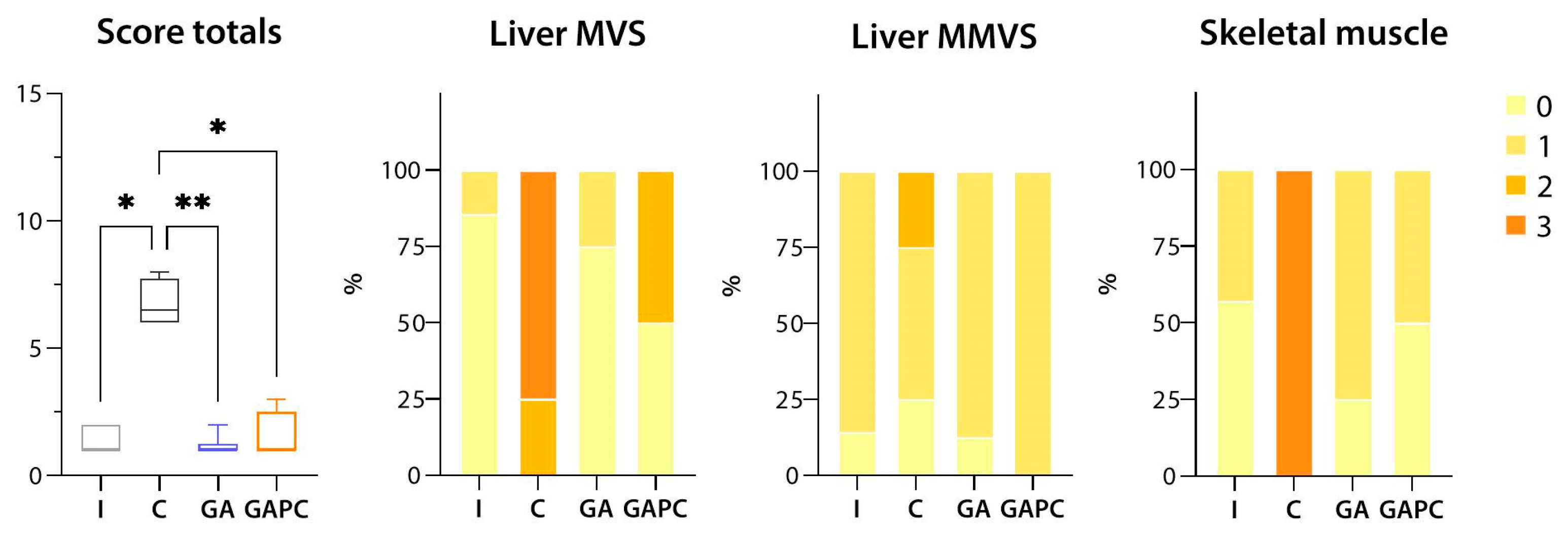
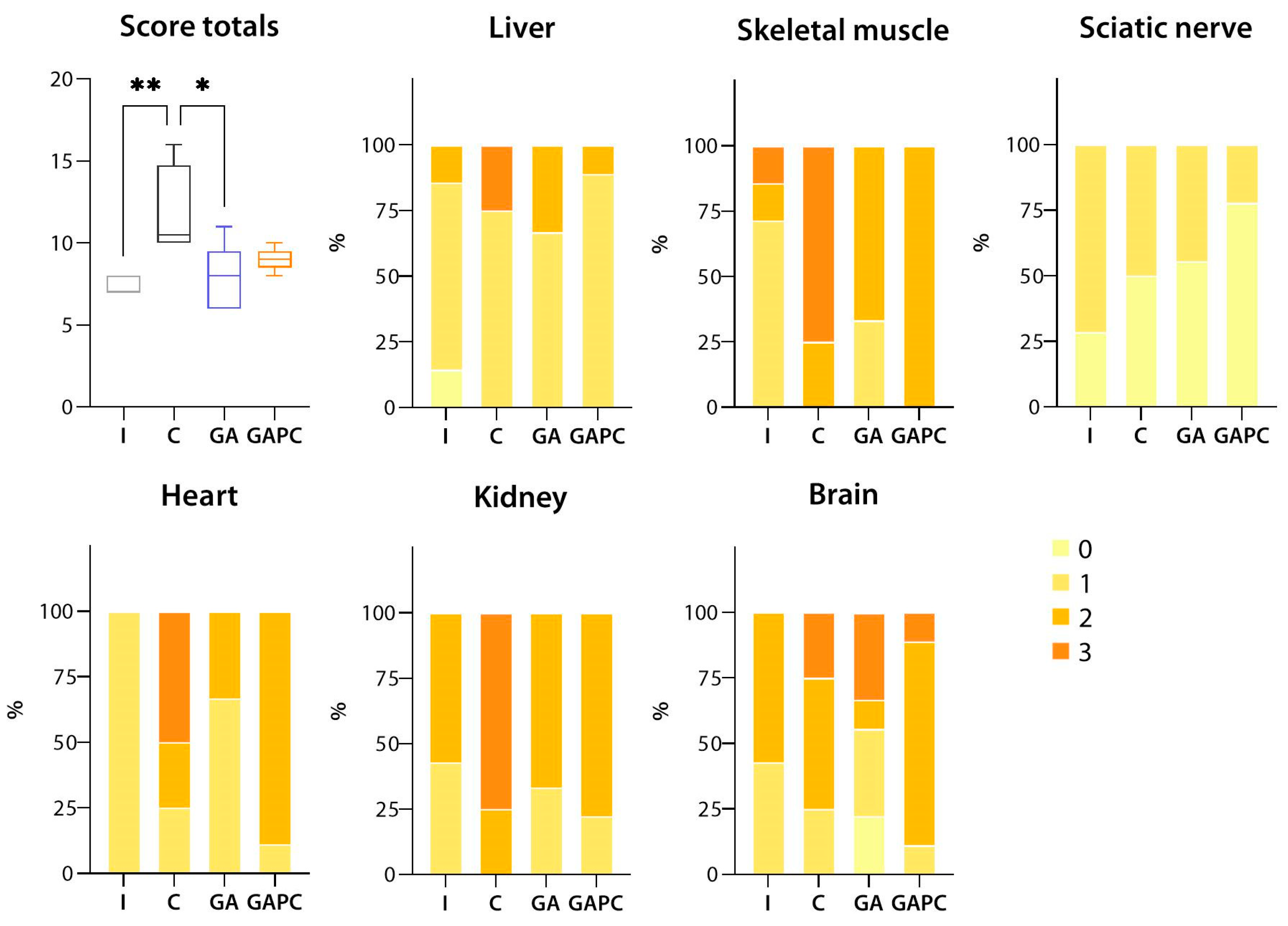
3.1.1. Liver
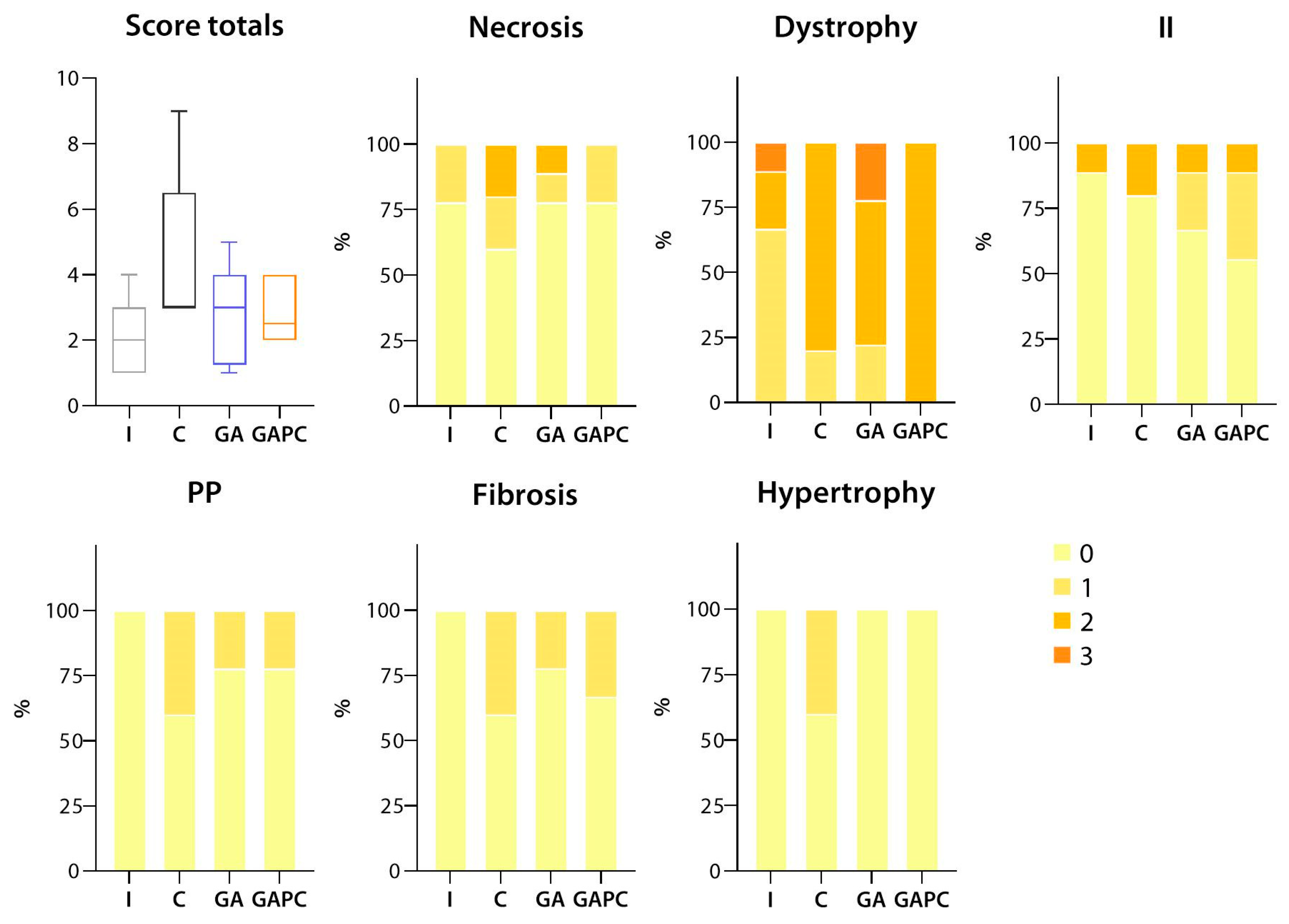
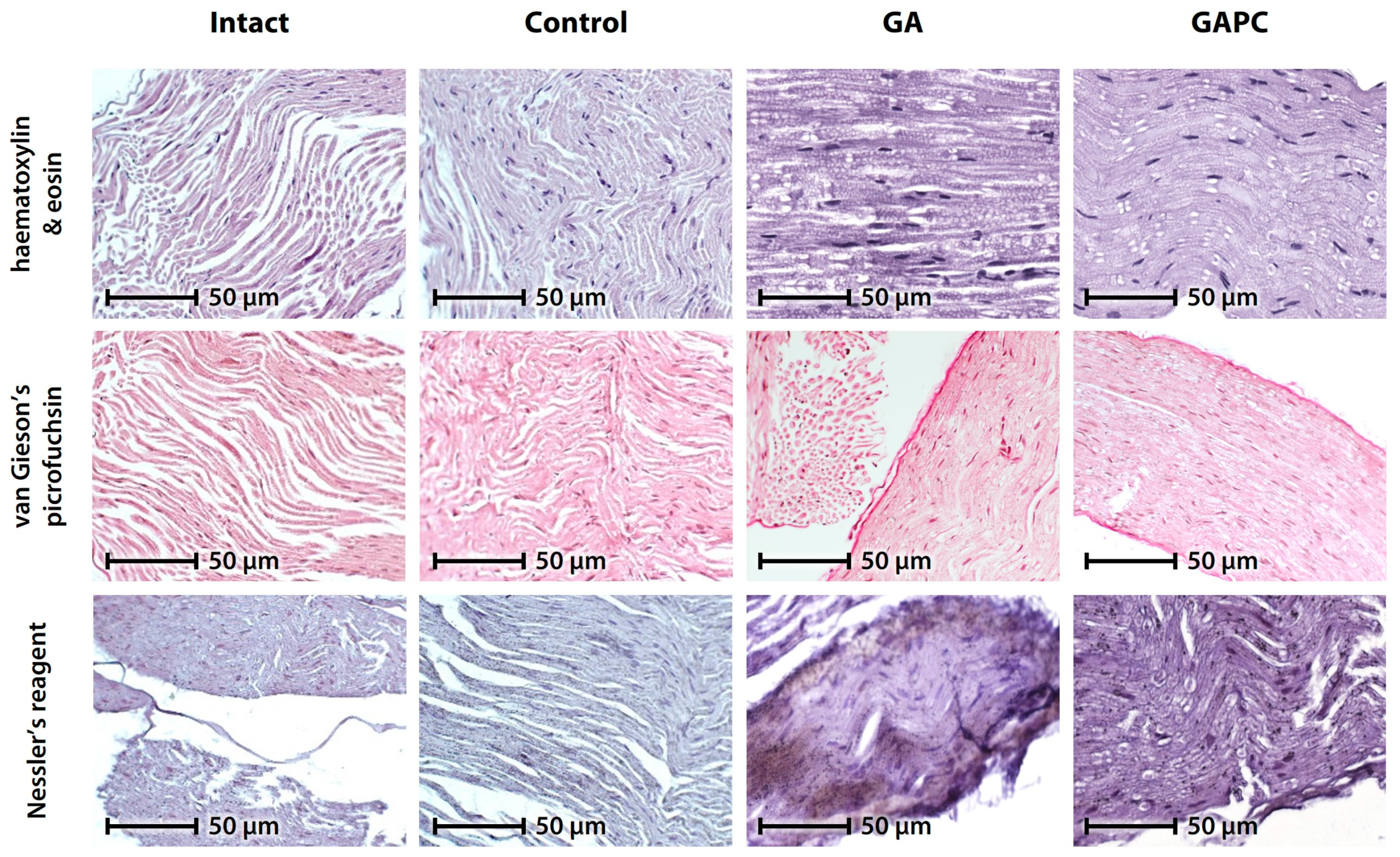
3.1.2. Skeletal Muscle
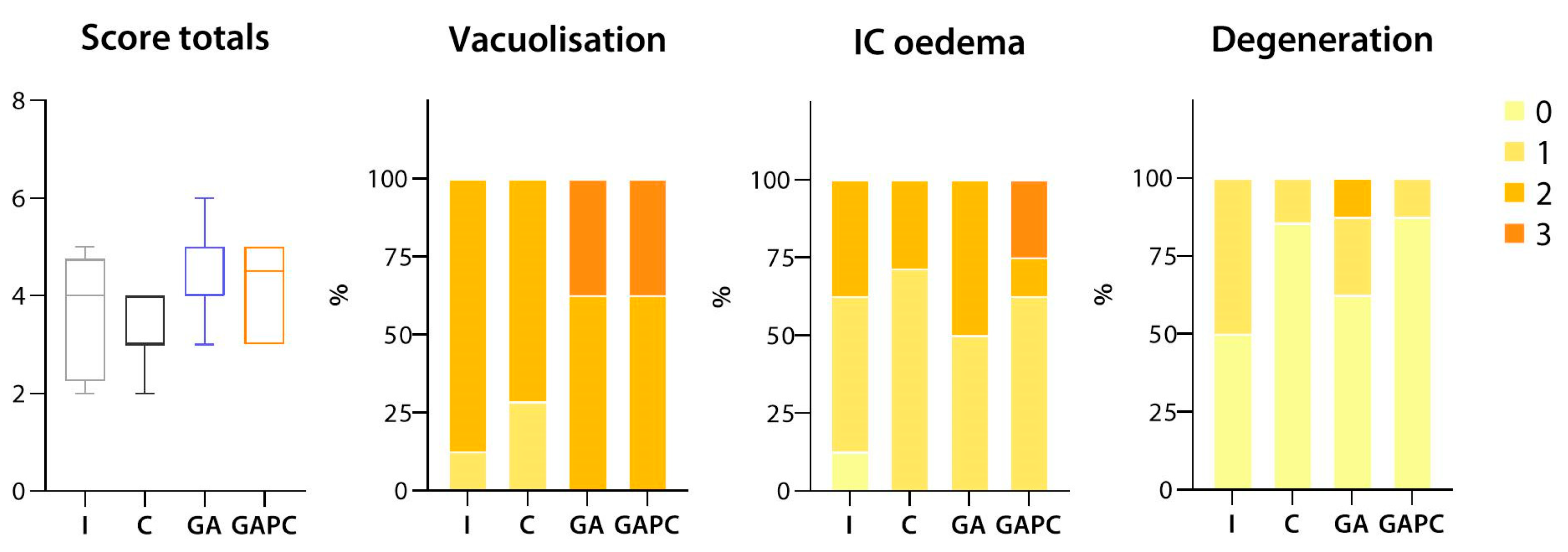
3.1.3. Sciatic Nerve
3.1.4. Tissue Lipid Content
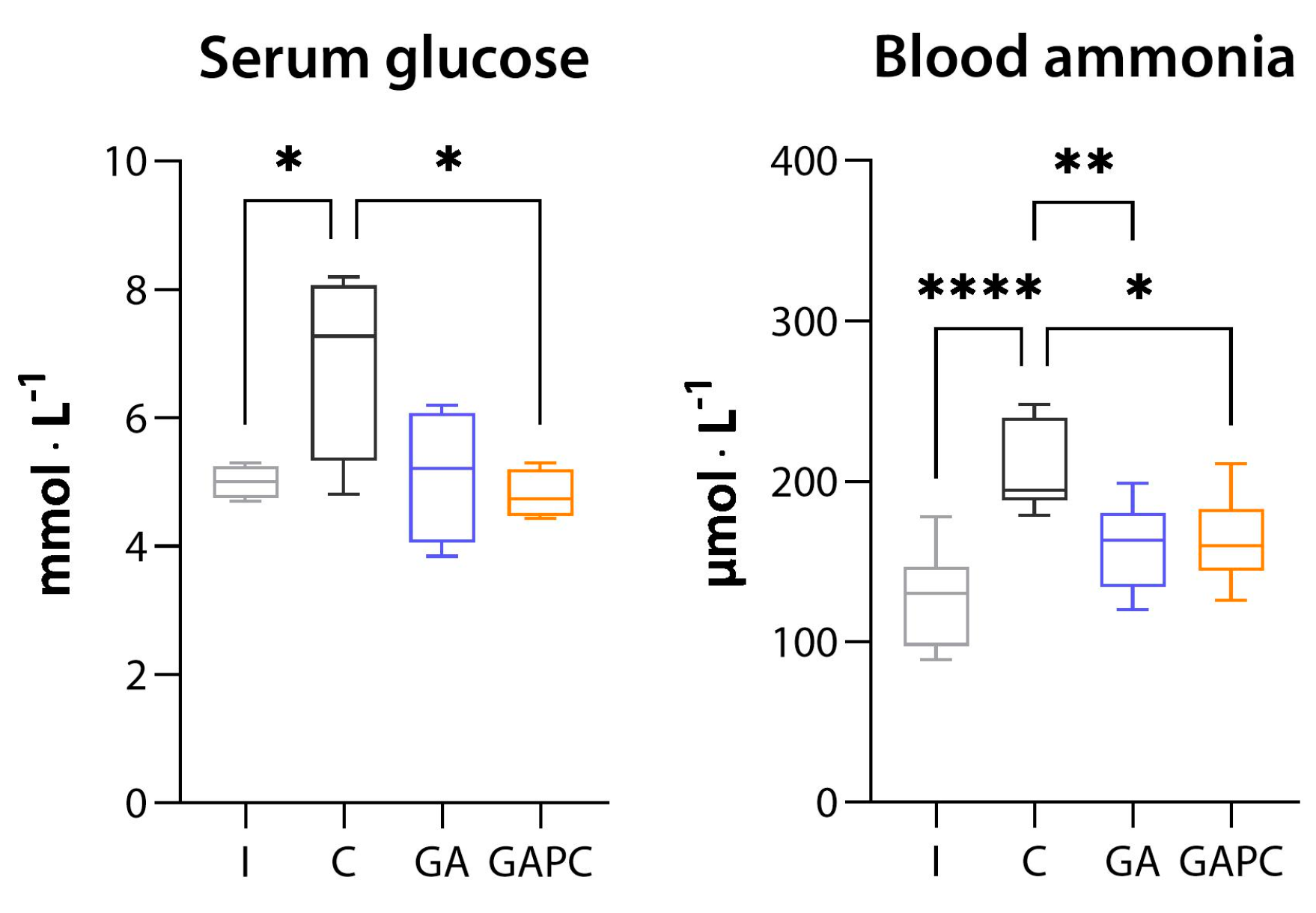
3.1.5. Tissue Ammonia Content
3.2. Blood Biochemistry
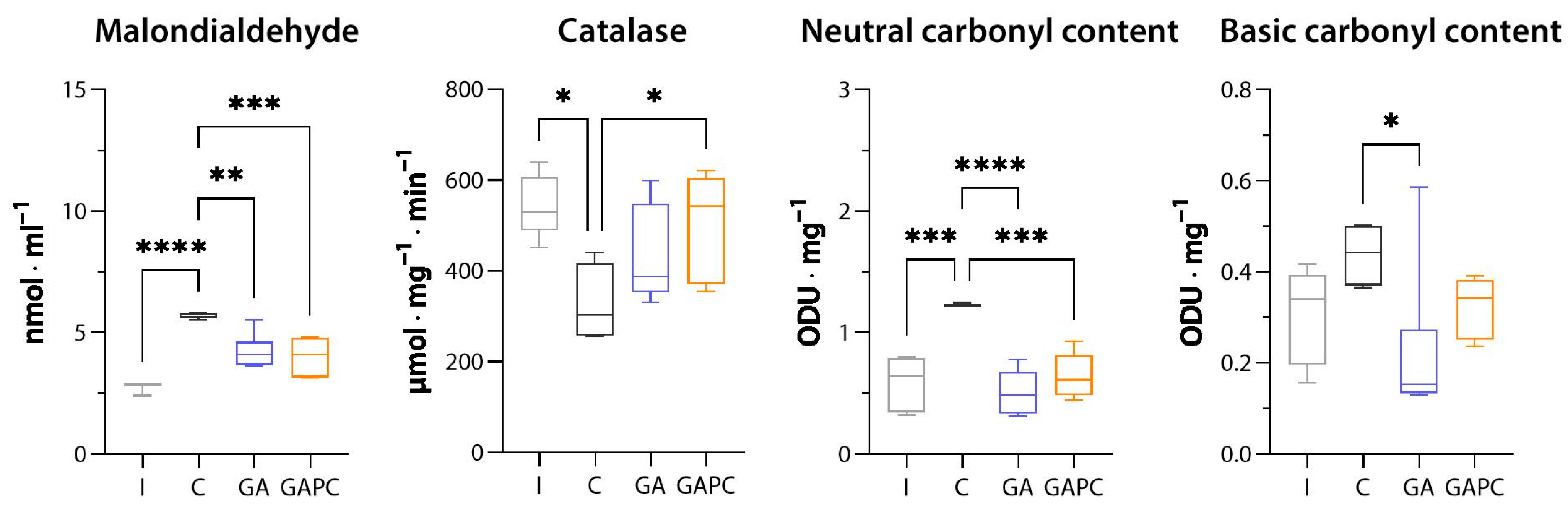
3.3. Oxidative Stress
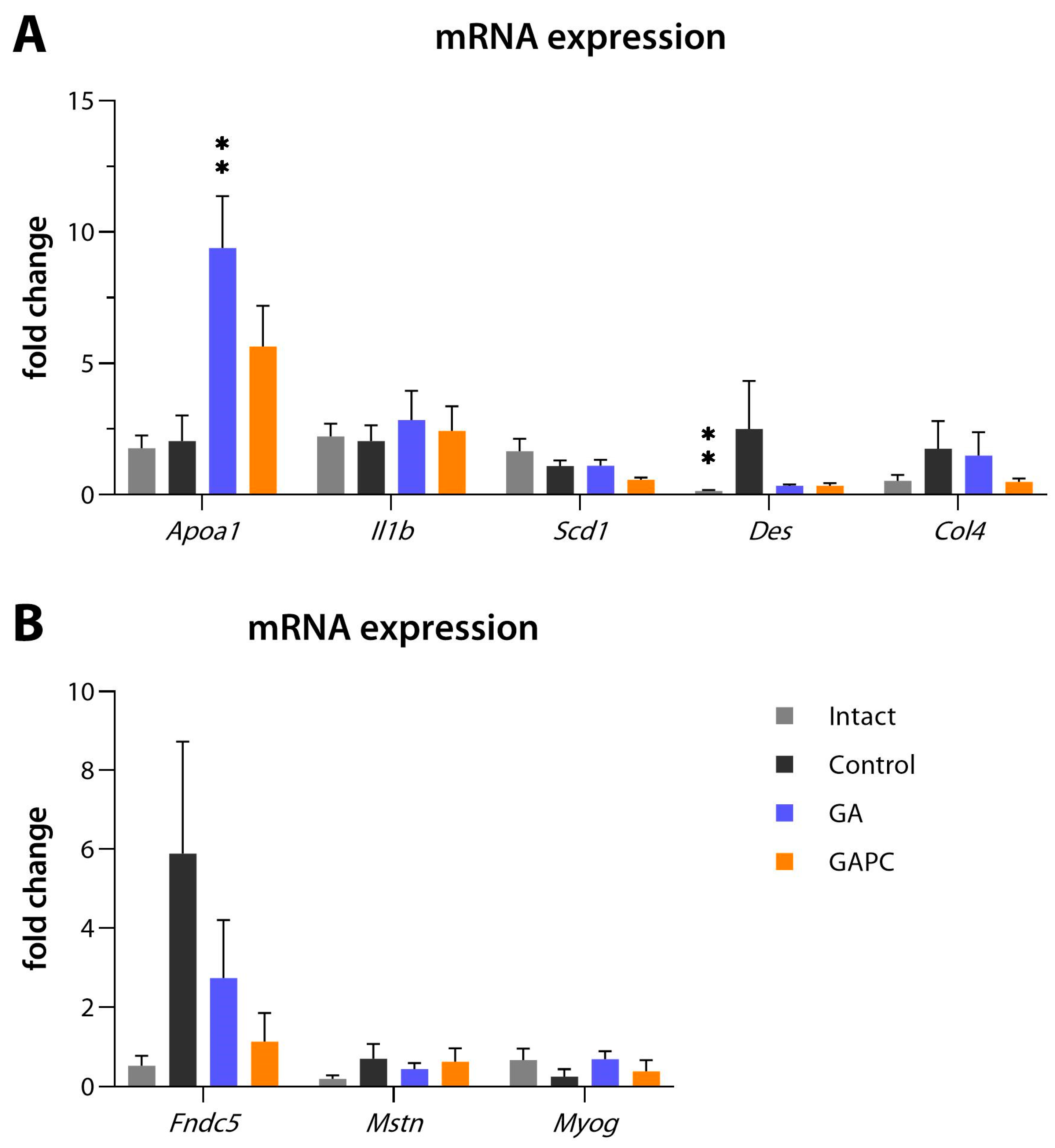
3.4. RT-qPCR
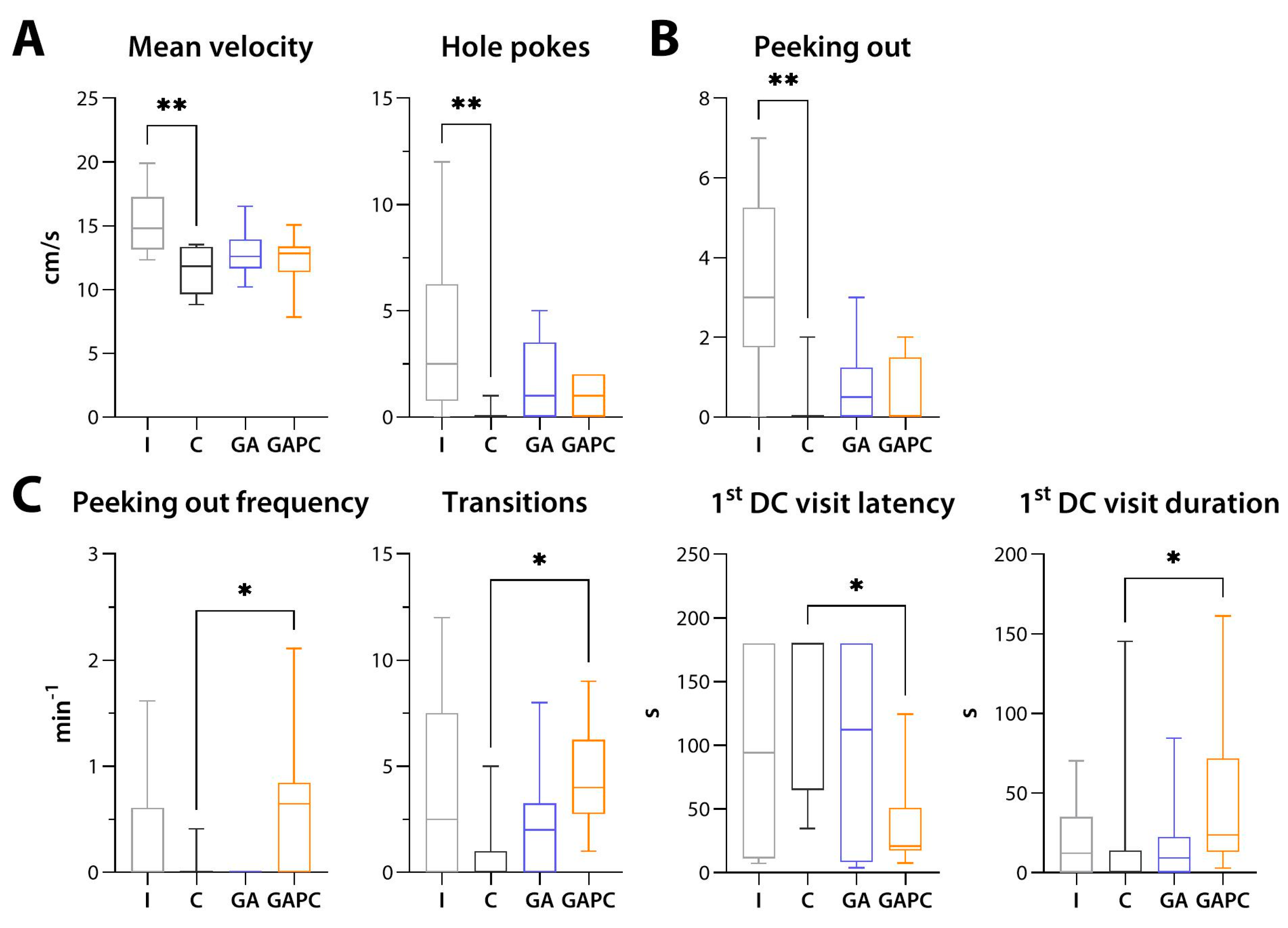
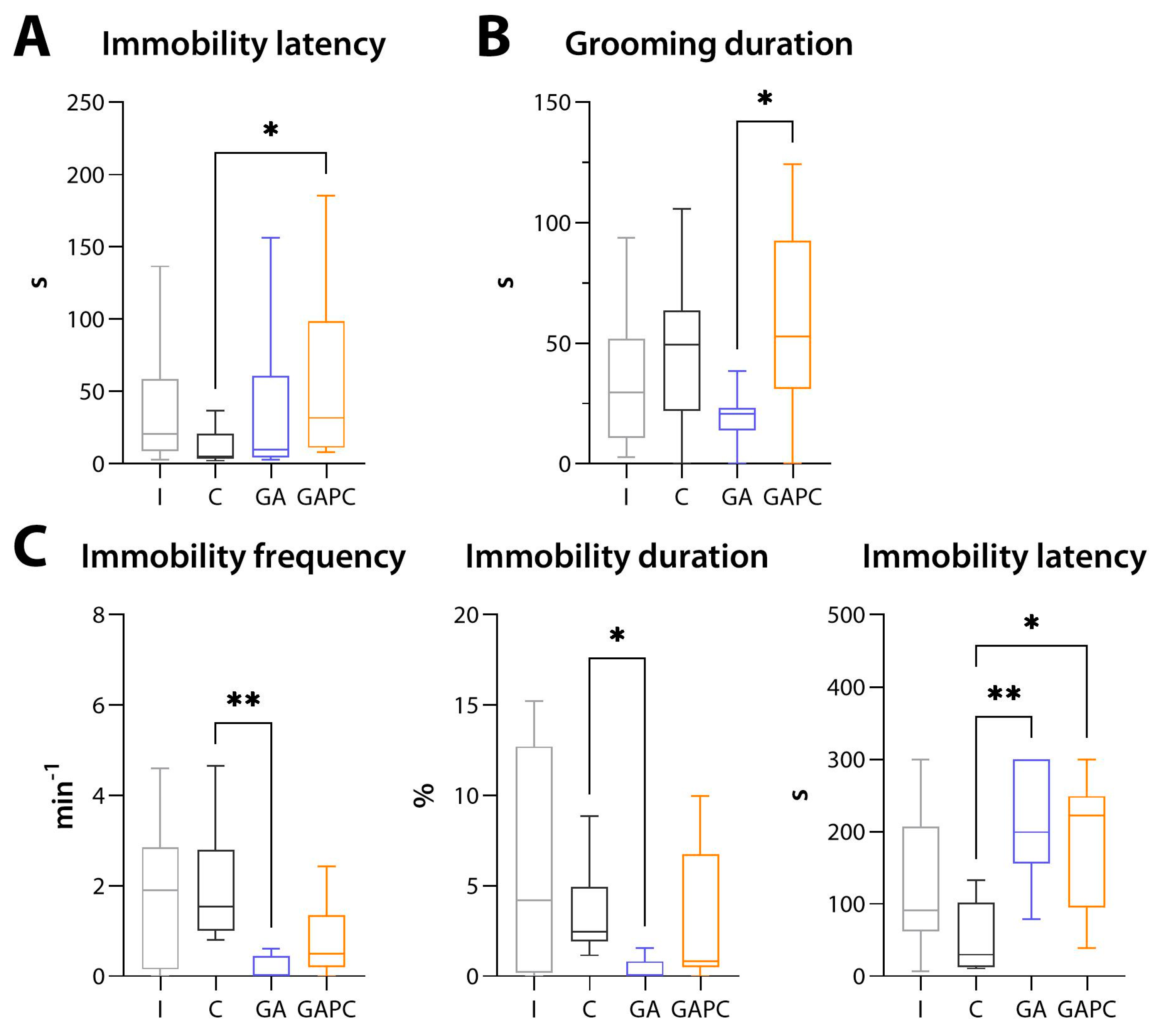
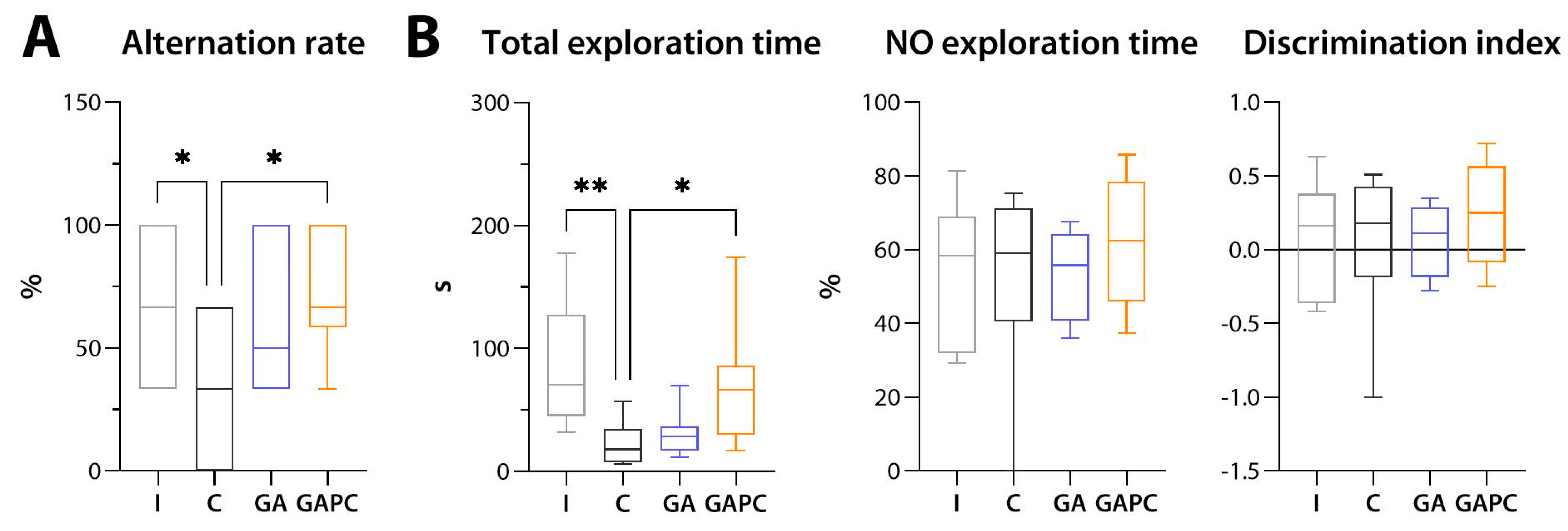
3.5. Locomotion, Behaviour, and Memory
3.5.1. General Locomotion and Anxiety-like Behaviour
3.5.2. Depression-like Behaviour
3.5.3. Memory
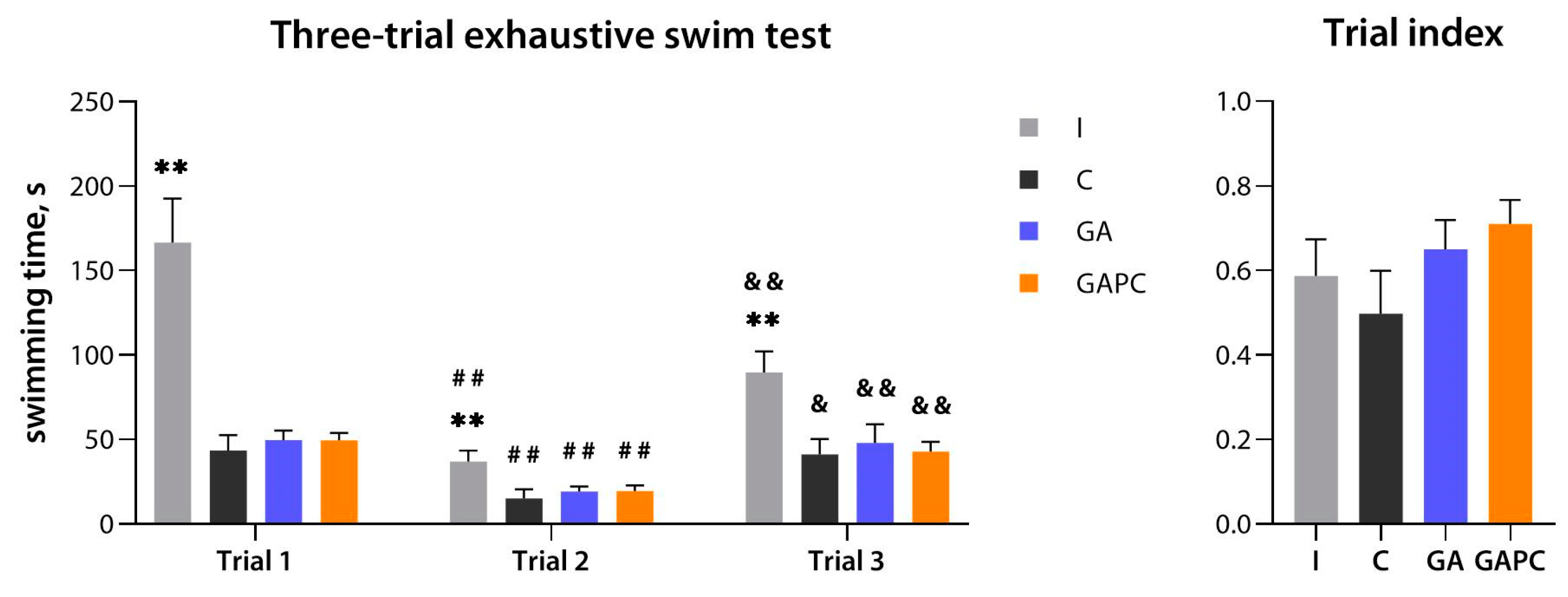
3.6. Physical Performance
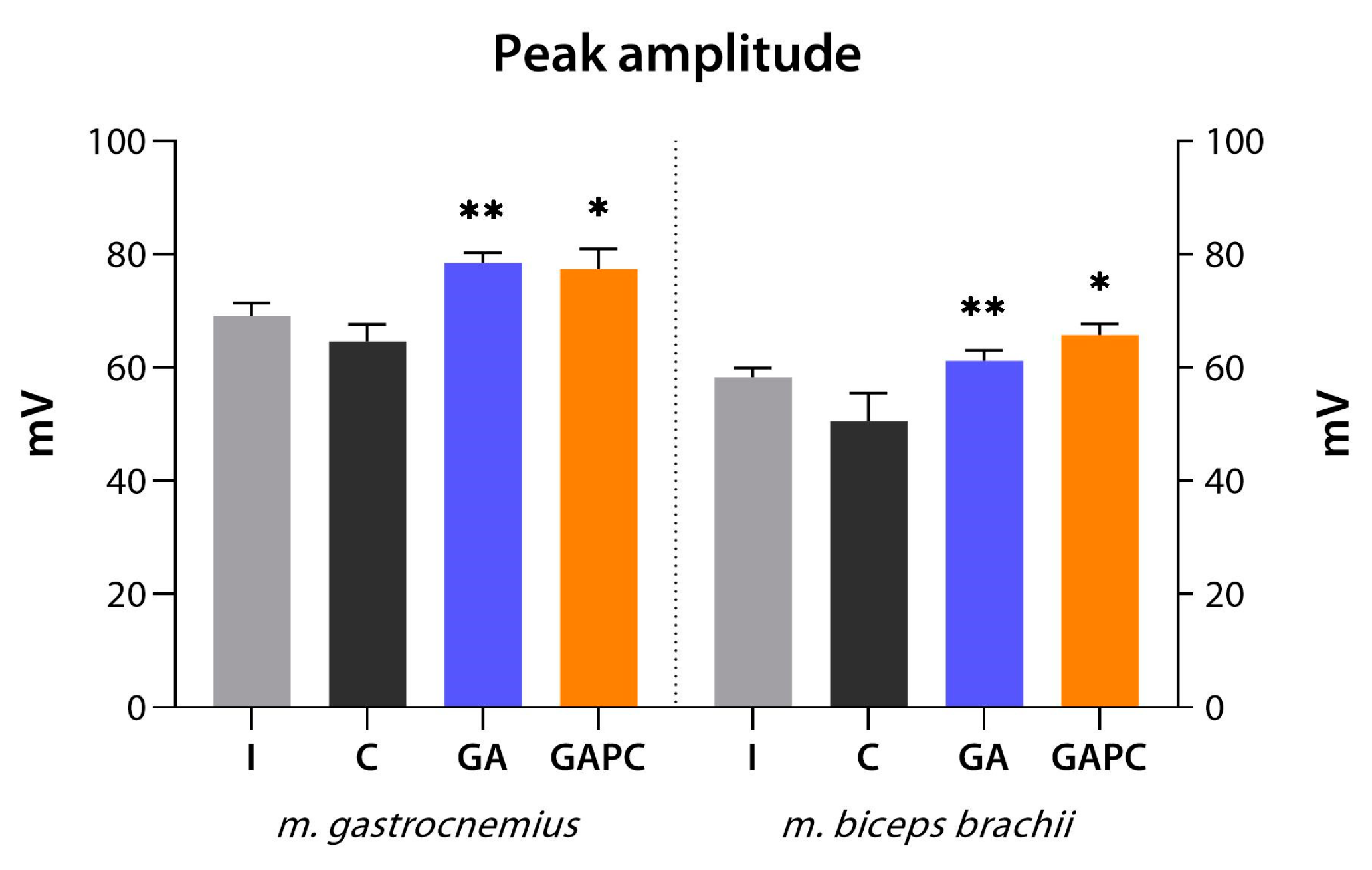
3.7. ENMG
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- European Association for the Study of the Liver (EASL); European Association for the Study of Diabetes (EASD); European Association for the Study of Obesity (EASO). EASL-EASD-EASO Clinical Practice Guidelines for the management of non-alcoholic fatty liver disease. J. Hepatol. 2016, 64, 1388–1402. [Google Scholar] [CrossRef]
- Monelli, F.; Venturelli, F.; Bonilauri, L.; Manicardi, E.; Manicardi, V.; Giorgi Rossi, P.; Massari, M.; Ligabue, G.; Riva, N.; Schianchi, S.; et al. Systematic review of existing guidelines for NAFLD assessment. Hepatoma Res. 2021, 7, 25. [Google Scholar] [CrossRef]
- Eslam, M.; Sanyal, A.J.; George, J.; International Consensus Panel. MAFLD: A Consensus-Driven Proposed Nomenclature for Metabolic Associated Fatty Liver Disease. Gastroenterology 2020, 158, 1999–2014.e1. [Google Scholar] [CrossRef]
- Lombardi, R.; Fargion, S.; Fracanzani, A.L. Brain Involvement in Non-Alcoholic Fatty Liver Disease (NAFLD): A Systematic Review. Dig. Liver Dis. 2019, 51, 1214–1222. [Google Scholar] [CrossRef]
- Kjærgaard, K.; Mikkelsen, A.C.D.; Wernberg, C.W.; Grønkjær, L.L.; Eriksen, P.L.; Damholdt, M.F.; Mookerjee, R.P.; Vilstrup, H.; Lauridsen, M.M.; Thomsen, K.L. Cognitive Dysfunction in Non-Alcoholic Fatty Liver Disease-Current Knowledge, Mechanisms and Perspectives. J. Clin. Med. 2021, 10, 673. [Google Scholar] [CrossRef]
- Gonzalez, A.; Huerta-Salgado, C.; Orozco-Aguilar, J.; Aguirre, F.; Tacchi, F.; Simon, F.; Cabello-Verrugio, C. Role of Oxidative Stress in Hepatic and Extrahepatic Dysfunctions during Nonalcoholic Fatty Liver Disease (NAFLD). Oxid. Med. Cell Longev. 2020, 2020, 1617805. [Google Scholar] [CrossRef] [PubMed]
- Muzurović, E.; Mikhailidis, D.P.; Mantzoros, C. Non-Alcoholic Fatty Liver Disease, Insulin Resistance, Metabolic Syndrome and Their Association with Vascular Risk. Metabolism 2021, 119, 154770. [Google Scholar] [CrossRef] [PubMed]
- Graebin, C.S. The Pharmacological Activities of Glycyrrhizinic Acid (“Glycyrrhizin”) and Glycyrrhetinic Acid. Sweeteners 2017, 245–261. [Google Scholar] [CrossRef]
- Kim, S.-W.; Jin, Y.; Shin, J.-H.; Kim, I.-D.; Lee, H.-K.; Park, S.; Han, P.-L.; Lee, J.-K. Glycyrrhizic Acid Affords Robust Neuroprotection in the Postischemic Brain via Anti-Inflammatory Effect by Inhibiting HMGB1 Phosphorylation and Secretion. Neurobiol. Dis. 2012, 46, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Sathyamoorthy, Y.; Kaliappan, K.; Nambi, P.; Radhakrishnan, R. Glycyrrhizic Acid Renders Robust Neuroprotection in Rodent Model of Vascular Dementia by Controlling Oxidative Stress and Curtailing Cytochrome-c Release. Nutr. Neurosci. 2020, 23, 955–970. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Gao, J.; Shen, J.; Zhu, X.; Wang, H.; Feng, S.; Huang, C.; Shen, H.; Liu, H. Glycyrrhizic Acid Improves Cognitive Levels of Aging Mice by Regulating T/B Cell Proliferation. Front. Aging Neurosci. 2020, 12, 570116. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Cao, H.; Liu, P.; Cheng, G.; Sun, M. Glycyrrhizic Acid in the Treatment of Liver Diseases: Literature Review. Biomed. Res. Int. 2014, 2014, 872139. [Google Scholar] [CrossRef]
- Chen, S.; Zou, L.; Li, L.; Wu, T. The Protective Effect of Glycyrrhetinic Acid on Carbon Tetrachloride-Induced Chronic Liver Fibrosis in Mice via Upregulation of Nrf2. PLoS ONE 2013, 8, e53662. [Google Scholar] [CrossRef] [PubMed]
- Yoke Yin, C.; So Ha, T.; Abdul Kadir, K. Effects of Glycyrrhizic Acid on Peroxisome Proliferator-Activated Receptor Gamma (PPARgamma), Lipoprotein Lipase (LPL), Serum Lipid and HOMA-IR in Rats. PPAR Res. 2010, 2010, 530265. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.-Y.; Cui, S.-C.; Wang, L.; Zhang, Y.-T.; Yan, X.-X.; Lu, H.-L.; Xing, G.-Z.; Ren, J.; Gong, L.-K. 18β-Glycyrrhetinic Acid Protects against Alpha-Naphthylisothiocyanate-Induced Cholestasis through Activation of the Sirt1/FXR Signaling Pathway. Acta Pharmacol. Sin. 2018, 39, 1865–1873. [Google Scholar] [CrossRef] [PubMed]
- Shinu, P.; Gupta, G.L.; Sharma, M.; Khan, S.; Goyal, M.; Nair, A.B.; Kumar, M.; Soliman, W.E.; Rahman, A.; Attimarad, M.; et al. Pharmacological Features of 18β-Glycyrrhetinic Acid: A Pentacyclic Triterpenoid of Therapeutic Potential. Plants 2023, 12, 1086. [Google Scholar] [CrossRef] [PubMed]
- Nicolson, G.L.; Ash, M.E. Lipid Replacement Therapy: A Natural Medicine Approach to Replacing Damaged Lipids in Cellular Membranes and Organelles and Restoring Function. Biomembranes 2014, 1838, 1657–1679. [Google Scholar] [CrossRef]
- Osipova, D.; Kokoreva, K.; Lazebnik, L.; Golovanova, E.; Pavlov, C.; Dukhanin, A.; Orlova, S.; Starostin, K. Regression of Liver Steatosis Following Phosphatidylcholine Administration: A Review of Molecular and Metabolic Pathways Involved. Front. Pharmacol. 2022, 13, 797923. [Google Scholar] [CrossRef]
- Singh, R.P.; Gangadharappa, H.V.; Mruthunjaya, K. Phospholipids: Unique Carriers for Drug Delivery Systems. J. Drug. Deliv. Sci. Technol. 2017, 39, 166–179. [Google Scholar] [CrossRef]
- Tsuchida, T.; Lee, Y.A.; Fujiwara, N.; Ybanez, M.; Allen, B.; Martins, S.; Fiel, M.I.; Goossens, N.; Chou, H.-I.; Hoshida, Y.; et al. A Simple Diet- and Chemical-Induced Murine NASH Model with Rapid Progression of Steatohepatitis, Fibrosis and Liver Cancer. J. Hepatol. 2018, 69, 385–395. [Google Scholar] [CrossRef]
- Nair, A.B.; Jacob, S. A Simple Practice Guide for Dose Conversion between Animals and Human. J. Basic Clin. Pharm. 2016, 7, 27–31. [Google Scholar] [CrossRef] [PubMed]
- Phosphogliv Forte. Registration Certificate. Available online: https://grls.rosminzdrav.ru/Grls_View_v2.aspx?routingGuid=49b3de04-9280-43ea-a3c6-e88c872a924c (accessed on 6 December 2023).
- Walsh, R.N.; Cummins, R.A. The Open-Field Test: A Critical Review. Psychol. Bull. 1976, 83, 482–504. [Google Scholar] [CrossRef] [PubMed]
- Walf, A.A.; Frye, C.A. The Use of the Elevated plus Maze as an Assay of Anxiety-Related Behavior in Rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef] [PubMed]
- Bourin, M.; Hascoët, M. The Mouse Light/Dark Box Test. Eur. J. Pharmacol. 2003, 463, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Can, A.; Dao, D.T.; Terrillion, C.E.; Piantadosi, S.C.; Bhat, S.; Gould, T.D. The Tail Suspension Test. J. Vis. Exp. 2012, 59, e3769. [Google Scholar] [CrossRef]
- Can, A.; Dao, D.T.; Arad, M.; Terrillion, C.E.; Piantadosi, S.C.; Gould, T.D. The Mouse Forced Swim Test. J. Vis. Exp. 2012, 59, e3638. [Google Scholar] [CrossRef]
- Isingrini, E.; Camus, V.; Le Guisquet, A.-M.; Pingaud, M.; Devers, S.; Belzung, C. Association between Repeated Unpredictable Chronic Mild Stress (UCMS) Procedures with a High Fat Diet: A Model of Fluoxetine Resistance in Mice. PLoS ONE 2010, 5, e10404. [Google Scholar] [CrossRef]
- Deacon, R.M.J.; Rawlins, J.N.P. T-Maze Alternation in the Rodent. Nat. Protoc. 2006, 1, 7–12. [Google Scholar] [CrossRef]
- Leger, M.; Quiedeville, A.; Bouet, V.; Haelewyn, B.; Boulouard, M.; Schumann-Bard, P.; Freret, T. Object Recognition Test in Mice. Nat. Protoc. 2013, 8, 2531–2537. [Google Scholar] [CrossRef]
- Prikhodko, V.A.; Sysoev, Y.I.; Poveryaeva, M.A.; Bunyat, A.V.; Karev, V.E.; Ivkin, D.Y.; Sukhanov, D.S.; Shustov, E.B.; Okovityi, S.V. Effects of empagliflozin and L-ornithine L-aspartate on behavior, cognitive functions, and physical performance in mice with experimentally induced steatohepatitis. Bull. RSMU 2020, 3, 49–57. [Google Scholar] [CrossRef]
- Pollari, E.; Prior, R.; Robberecht, W.; Van Damme, P.; Van Den Bosch, L. In Vivo Electrophysiological Measurement of Compound Muscle Action Potential from the Forelimbs in Mouse Models of Motor Neuron Degeneration. J. Vis. Exp. 2018, 136, 57741. [Google Scholar] [CrossRef]
- Gutiérrez-de-Juan, V.; López de Davalillo, S.; Fernández-Ramos, D.; Barbier-Torres, L.; Zubiete-Franco, I.; Fernández-Tussy, P.; Simon, J.; Lopitz-Otsoa, F.; de las Heras, J.; Iruzubieta, P.; et al. A Morphological Method for Ammonia Detection in Liver. PLoS ONE 2017, 12, e0173914. [Google Scholar] [CrossRef]
- Botsoglou, N.A.; Fletouris, D.J.; Papageorgiou, G.E.; Vassilopoulos, V.N.; Mantis, A.J.; Trakatellis, A.G. Rapid, Sensitive, and Specific Thiobarbituric Acid Method for Measuring Lipid Peroxidation in Animal Tissue, Food, and Feedstuff Samples. J. Agricult. Food Chem. 1994, 42, 1931–1937. [Google Scholar] [CrossRef]
- Góth, L. A Simple Method for Determination of Serum Catalase Activity and Revision of Reference Range. Clin. Chim. Acta 1991, 196, 143–151. [Google Scholar] [CrossRef]
- Kostiuk, V.A.; Potapovich, A.I.; Kovaleva, Z.V. A simple and sensitive method of determination of superoxide dismutase activity based on the reaction of quercetin oxidation. Vopr. Med. Khim. 1990, 36, 88–91. (In Russian) [Google Scholar] [PubMed]
- Levine, R.L.; Garland, D.; Oliver, C.N.; Amici, A.; Climent, I.; Lenz, A.G.; Ahn, B.W.; Shaltiel, S.; Stadtman, E.R. Determination of Carbonyl Content in Oxidatively Modified Proteins. Methods Enzymol. 1990, 186, 464–478. [Google Scholar] [CrossRef]
- Hervé, M. RVAideMemoire: Testing and Plotting Procedures for Biostatistics. R Package Version 0.9-83. 2023. Available online: https://CRAN.R-project.org/package=RVAideMemoire (accessed on 28 June 2023).
- Prikhodko, V.A.; Karev, V.E.; Sysoev, Y.I.; Ivkin, D.Y.; Okovityi, S.V. A Simple Algorithm for Semiquantitative Analysis of Scored Histology Data in the R Environment, on the Example of Murine Non-Alcoholic Steatohepatitis Pharmacotherapy. Livers 2022, 2, 412–424. [Google Scholar] [CrossRef]
- Brown, G.T.; Kleiner, D.E. Histopathology of Nonalcoholic Fatty Liver Disease and Nonalcoholic Steatohepatitis. Metabolism 2016, 65, 1080–1086. [Google Scholar] [CrossRef]
- Feng, T.-T.; Yang, X.-Y.; Hao, S.-S.; Sun, F.-F.; Huang, Y.; Lin, Q.-S.; Pan, W. TLR-2-Mediated Metabolic Reprogramming Participates in Polyene Phosphatidylcholine-Mediated Inhibition of M1 Macrophage Polarization. Immunol. Res. 2020, 68, 28–38. [Google Scholar] [CrossRef]
- Lu, Y.; Feng, T.; Zhao, J.; Jiang, P.; Xu, D.; Zhou, M.; Dai, M.; Wu, J.; Sun, F.; Yang, X.; et al. Polyene Phosphatidylcholine Ameliorates High Fat Diet-Induced Non-Alcoholic Fatty Liver Disease via Remodeling Metabolism and Inflammation. Front. Physiol. 2022, 13, 810143. [Google Scholar] [CrossRef]
- Treede, I.; Braun, A.; Sparla, R.; Kühnel, M.; Giese, T.; Turner, J.R.; Anes, E.; Kulaksiz, H.; Füllekrug, J.; Stremmel, W.; et al. Anti-Inflammatory Effects of Phosphatidylcholine. J. Biol. Chem. 2007, 282, 27155–27164. [Google Scholar] [CrossRef]
- Chen, M.; Pan, H.; Dai, Y.; Zhang, J.; Tong, Y.; Huang, Y.; Wang, M.; Huang, H. Phosphatidylcholine regulates NF-κB activation in attenuation of LPS-induced inflammation: Evidence from in vitro study. Anim. Cells Syst. 2017, 22, 7–14. [Google Scholar] [CrossRef]
- Eu, C.H.A.; Lim, W.Y.A.; Ton, S.H.; bin Abdul Kadir, K. Glycyrrhizic Acid Improved Lipoprotein Lipase Expression, Insulin Sensitivity, Serum Lipid and Lipid Deposition in High-Fat Diet-Induced Obese Rats. Lipids Health Dis. 2010, 9, 81. [Google Scholar] [CrossRef]
- Sun, X.; Duan, X.; Wang, C.; Liu, Z.; Sun, P.; Huo, X.; Ma, X.; Sun, H.; Liu, K.; Meng, Q. Protective Effects of Glycyrrhizic Acid against Non-Alcoholic Fatty Liver Disease in Mice. Eur. J. Pharmacol. 2017, 806, 75–82. [Google Scholar] [CrossRef]
- Ye, J.M.; Doyle, P.J.; Iglesias, M.A.; Watson, D.G.; Cooney, G.J.; Kraegen, E.W. Peroxisome Proliferator-Activated Receptor (PPAR)-Alpha Activation Lowers Muscle Lipids and Improves Insulin Sensitivity in High Fat-Fed Rats: Comparison with PPAR-Gamma Activation. Diabetes 2001, 50, 411–417. [Google Scholar] [CrossRef]
- Peeters, A.; Baes, M. Role of PPARα in Hepatic Carbohydrate Metabolism. PPAR Res. 2010, 2010, 572405. [Google Scholar] [CrossRef] [PubMed]
- Dentin, R.; Girard, J.; Postic, C. Carbohydrate Responsive Element Binding Protein (ChREBP) and Sterol Regulatory Element Binding Protein-1c (SREBP-1c): Two Key Regulators of Glucose Metabolism and Lipid Synthesis in Liver. Biochimie 2005, 87, 81–86. [Google Scholar] [CrossRef]
- Saito, H.; Ishihara, K. Antioxidant activity and active sites of phospholipids as antioxidants. J. Am. Oil Chem. Soc. 1997, 74, 1531–1536. [Google Scholar] [CrossRef]
- Del Ben, M.; Polimeni, L.; Carnevale, R.; Bartimoccia, S.; Nocella, C.; Baratta, F.; Loffredo, L.; Pignatelli, P.; Violi, F.; Angelico, F. NOX2-Generated Oxidative Stress Is Associated with Severity of Ultrasound Liver Steatosis in Patients with Non-Alcoholic Fatty Liver Disease. BMC Gastroenterol. 2014, 14, 81. [Google Scholar] [CrossRef]
- Yang, Z.; Yang, J.; Cai, J.; Zhang, X.-J.; Zhang, P.; She, Z.-G.; Li, H. The Transition of Cardiovascular Disease Risks from NAFLD to MAFLD. Rev. Cardiovasc. Med. 2023, 24, 157. [Google Scholar] [CrossRef]
- Kessoku, T.; Kobayashi, T.; Imajo, K.; Tanaka, K.; Yamamoto, A.; Takahashi, K.; Kasai, Y.; Ozaki, A.; Iwaki, M.; Nogami, A.; et al. Endotoxins and Non-Alcoholic Fatty Liver Disease. Front. Endocrinol. 2021, 12, 770986. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Li, H.; Song, J.; Zhang, C.; Li, M.; Mao, Y.; Liu, A.; Du, J. Role of Apolipoprotein A1 in PPAR Signaling Pathway for Nonalcoholic Fatty Liver Disease. PPAR Res. 2022, 2022, 4709300. [Google Scholar] [CrossRef] [PubMed]
- Karavia, E.A.; Papachristou, D.J.; Liopeta, K.; Triantaphyllidou, I.-E.; Dimitrakopoulos, O.; Kypreos, K.E. Apolipoprotein A-I Modulates Processes Associated with Diet-Induced Nonalcoholic Fatty Liver Disease in Mice. Mol. Med. 2012, 18, 901–912. [Google Scholar] [CrossRef] [PubMed]
- Geerts, A.; Eliasson, C.; Niki, T.; Wielant, A.; Vaeyens, F.; Pekny, M. Formation of Normal Desmin Intermediate Filaments in Mouse Hepatic Stellate Cells Requires Vimentin. Hepatology 2001, 33, 177–188. [Google Scholar] [CrossRef] [PubMed]
- Rabiee, F.; Lachinani, L.; Ghaedi, S.; Nasr-Esfahani, M.H.; Megraw, T.L.; Ghaedi, K. New Insights into the Cellular Activities of Fndc5/Irisin and Its Signaling Pathways. Cell Biosci. 2020, 10, 51. [Google Scholar] [CrossRef] [PubMed]
- Stefano, J.T.; Guedes, L.V.; de Souza, A.A.A.; Vanni, D.S.; Alves, V.A.F.; Carrilho, F.J.; Largura, A.; Arrese, M.; Oliveira, C.P. Usefulness of Collagen Type IV in the Detection of Significant Liver Fibrosis in Nonalcoholic Fatty Liver Disease. Ann. Hepatol. 2021, 20, 100253. [Google Scholar] [CrossRef] [PubMed]
- Mridha, A.R.; Wree, A.; Robertson, A.A.B.; Yeh, M.M.; Johnson, C.D.; Van Rooyen, D.M.; Haczeyni, F.; Teoh, N.C.-H.; Savard, C.; Ioannou, G.N.; et al. NLRP3 Inflammasome Blockade Reduces Liver Inflammation and Fibrosis in Experimental NASH in Mice. J. Hepatol. 2017, 66, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.-M.; Jiang, Q.-H.; Cai, C.; Qu, M.; Shen, W. SCD1 Negatively Regulates Autophagy-Induced Cell Death in Human Hepatocellular Carcinoma through Inactivation of the AMPK Signaling Pathway. Cancer Lett. 2015, 358, 180–190. [Google Scholar] [CrossRef]
- Ryan, A.S.; Li, G. Skeletal Muscle Myostatin Gene Expression and Sarcopenia in Overweight and Obese Middle-Aged and Older Adults. JCSM Clin. Rep. 2021, 6, 137–142. [Google Scholar] [CrossRef]
- Langley, B.; Thomas, M.; Bishop, A.; Sharma, M.; Gilmour, S.; Kambadur, R. Myostatin Inhibits Myoblast Differentiation by Down-Regulating MyoD Expression. J. Biol. Chem. 2002, 277, 49831–49840. [Google Scholar] [CrossRef]
- Veniaminova, E.; Cespuglio, R.; Markova, N.; Mortimer, N.; Cheung, C.W.; Steinbusch, H.W.; Lesch, K.-P.; Strekalova, T. Behavioral features of mice fed with a cholesterol-enriched diet: Deficient novelty exploration and unaltered aggressive behavior. Transl. Neurosci. Clin. 2016, 2, 87–95. [Google Scholar] [CrossRef]
- Wang, B.; Lian, Y.-J.; Dong, X.; Peng, W.; Liu, L.-L.; Su, W.-J.; Gong, H.; Zhang, T.; Jiang, C.-L.; Li, J.-S.; et al. Glycyrrhizic Acid Ameliorates the Kynurenine Pathway in Association with Its Antidepressant Effect. Behav. Brain Res. 2018, 353, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Gupta, G.L.; Sharma, L.; Sharma, M. 18β-Glycyrrhetinic Acid Ameliorates Neuroinflammation Linked Depressive Behavior Instigated by Chronic Unpredictable Mild Stress via Triggering BDNF/TrkB Signaling Pathway in Rats. Neurochem. Res. 2023, 48, 551–569. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.-Y.; Liu, Y.-Z.; Li, J.-M.; Ruan, Y.-M.; Yan, W.-J.; Zhong, S.-Y.; Zhang, T.; Liu, L.-L.; Wu, R.; Wang, B.; et al. Glycyrrhizic Acid as an Adjunctive Treatment for Depression through Anti-Inflammation: A Randomized Placebo-Controlled Clinical Trial. J. Affect. Disord. 2020, 265, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Magaquian, D.; Delgado Ocaña, S.; Perez, C.; Banchio, C. Phosphatidylcholine Restores Neuronal Plasticity of Neural Stem Cells under Inflammatory Stress. Sci. Rep. 2021, 11, 22891. [Google Scholar] [CrossRef] [PubMed]
- Leone, P.; Mincheva, G.; Balzano, T.; Malaguarnera, M.; Felipo, V.; Llansola, M. Rifaximin Improves Spatial Learning and Memory Impairment in Rats with Liver Damage-Associated Neuroinflammation. Biomedicines 2022, 10, 1263. [Google Scholar] [CrossRef] [PubMed]
- Prikhodko, V.A. Effects of ornithine aspartate and empagliflozin on memory deficit symptoms in experimental steatohepatitis. J. Biomed. 2022, 18, 128–132. [Google Scholar] [CrossRef]
- Broadbent, N.J.; Squire, L.R.; Clark, R.E. Spatial Memory, Recognition Memory, and the Hippocampus. Proc. Natl. Acad. Sci. USA 2004, 101, 14515–14520. [Google Scholar] [CrossRef] [PubMed]
- Ban, J.Y.; Park, H.K.; Kim, S.K. Effect of Glycyrrhizic Acid on Scopolamine-Induced Cognitive Impairment in Mice. Int. Neurourol. J. 2020, 24, S48–S55. [Google Scholar] [CrossRef]
- Song, J.-H.; Lee, J.-W.; Shim, B.; Lee, C.-Y.; Choi, S.; Kang, C.; Sohn, N.-W.; Shin, J.-W. Glycyrrhizin Alleviates Neuroinflammation and Memory Deficit Induced by Systemic Lipopolysaccharide Treatment in Mice. Molecules 2013, 18, 15788–15803. [Google Scholar] [CrossRef]
- Liu, W.; Huang, S.; Li, Y.; Zhang, K.; Zheng, X. Suppressive Effect of Glycyrrhizic Acid against Lipopolysaccharide-Induced Neuroinflammation and Cognitive Impairment in C57 Mice via Toll-like Receptor 4 Signaling Pathway. Food Nutr. Res. 2019, 63, 1516. [Google Scholar] [CrossRef]
- Jayakumar, A.R.; Norenberg, M.D. Hyperammonemia in Hepatic Encephalopathy. J. Clin. Exp. Hepatol. 2018, 8, 272–280. [Google Scholar] [CrossRef]
- Correa-de-Araujo, R.; Addison, O.; Miljkovic, I.; Goodpaster, B.H.; Bergman, B.C.; Clark, R.V.; Elena, J.W.; Esser, K.A.; Ferrucci, L.; Harris-Love, M.O.; et al. Myosteatosis in the Context of Skeletal Muscle Function Deficit: An Interdisciplinary Workshop at the National Institute on Aging. Front. Physiol. 2020, 11, 963. [Google Scholar] [CrossRef] [PubMed]
- Serra, A.J.; Pinto, J.R.; Prokić, M.D.; Arsa, G.; Vasconsuelo, A. Oxidative Stress in Muscle Diseases: Current and Future Therapy 2019. Oxid. Med. Cell. Longev. 2020, 2020, 6030417. [Google Scholar] [CrossRef]
- Tuttle, C.S.L.; Thang, L.A.N.; Maier, A.B. Markers of Inflammation and Their Association with Muscle Strength and Mass: A Systematic Review and Meta-Analysis. Ageing Res. Rev. 2020, 64, 101185. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-W.; Dunn, M.A. Muscle at Risk: The Multiple Impacts of Ammonia on Sarcopenia and Frailty in Cirrhosis. Clin. Transl. Gastroenterol. 2016, 7, e170. [Google Scholar] [CrossRef] [PubMed]
- Di Cola, S.; Nardelli, S.; Ridola, L.; Gioia, S.; Riggio, O.; Merli, M. Ammonia and the Muscle: An Emerging Point of View on Hepatic Encephalopathy. J. Clin. Med. 2022, 11, 611. [Google Scholar] [CrossRef] [PubMed]
- Mizoguchi, K.; Kanno, H.; Ikarashi, Y.; Kase, Y. Specific Binding and Characteristics of 18β-Glycyrrhetinic Acid in Rat Brain. PLoS ONE 2014, 9, e95760. [Google Scholar] [CrossRef]
- Javaid, S.; Farooq, T.; Rehman, Z.; Afzal, A.; Ashraf, W.; Rasool, M.F.; Alqahtani, F.; Alsanea, S.; Alasmari, F.; Alanazi, M.M.; et al. Dynamics of Choline-Containing Phospholipids in Traumatic Brain Injury and Associated Comorbidities. Int. J. Mol. Sci. 2021, 22, 11313. [Google Scholar] [CrossRef]


















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer, 5′-3′ | Reverse Primer, 5′-3′ |
|---|---|---|
| Apoa1 | GTGGCTCTGGTCTTCCTGAC | ACGGTTGAACCCAGAGTGTC |
| Il1b | CTGCAGCTGGAGAGTGTGGAT | CTCCACTTTGCTCTTGACTTCTATCTT |
| Scd1 | CCGGAGACCCCTTAGATCG | TAGCCTGTAAAAGATTTCTGCAAACC |
| Des | AAGATGGCCTTGGATGTGGA | GTTGATCCTGCTCTCCTCGC |
| Col4 | TGATAAAGGTTCCCGAGGAG | ATCCTGGTGTCCCACTAAGG |
| Fndc5 | ACAGGCAGAGAGCAGAGAGC | GAAGTCTGCTGCCACATCAA |
| Mstn | AGTGGATCTAAATGAGGGCAGT | GTTTCCAGGCGCAGCTTAC |
| Myog | GGGCAATGCACTGGAGTT | CACGATGGACGTAAGGGAGT |
| Actb | AAGATCCTGACCGAGCGTGGCT | AGGGAGGAAGAGGATGCGGCAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Prikhodko, V.A.; Matuzok, T.M.; Karev, V.E.; Karavaeva, A.V.; Spasenkova, O.M.; Kirillova, N.V.; Ivkin, D.Y.; Okovityi, S.V. Glycyrrhizinic Acid and Phosphatidylcholine Combination as a Preventive Therapy for Experimental Murine Non-Alcoholic Steatohepatitis. Livers 2024, 4, 63-83. https://doi.org/10.3390/livers4010006
Prikhodko VA, Matuzok TM, Karev VE, Karavaeva AV, Spasenkova OM, Kirillova NV, Ivkin DY, Okovityi SV. Glycyrrhizinic Acid and Phosphatidylcholine Combination as a Preventive Therapy for Experimental Murine Non-Alcoholic Steatohepatitis. Livers. 2024; 4(1):63-83. https://doi.org/10.3390/livers4010006
Chicago/Turabian StylePrikhodko, Veronika A., Tatyana M. Matuzok, Vadim E. Karev, Anna V. Karavaeva, Olga M. Spasenkova, Nadezhda V. Kirillova, Dmitry Yu. Ivkin, and Sergey V. Okovityi. 2024. "Glycyrrhizinic Acid and Phosphatidylcholine Combination as a Preventive Therapy for Experimental Murine Non-Alcoholic Steatohepatitis" Livers 4, no. 1: 63-83. https://doi.org/10.3390/livers4010006