1. Introduction
IgE-mediated allergy is one of the most common diseases in the world. It occurs in response to harmless non-replicating compounds, such as pollen of trees and grasses, fragments of household microorganisms, food, fungus spores and conidia, etc. Most allergy medications suppress the innate response to allergy mediators, preventing the activation and degranulation of mast cells. To date, the only way to attenuate allergy is allergen-specific immunotherapy (ASIT), which consists of a prolonged subcutaneous administration of small doses of allergen extracts [
1] lasting, in some cases, for years. It was shown that ASIT results in IgG4 production, which partially prevents the development of hypersensitivity to the allergens, and slows down the transition to severe forms of allergies, such as bronchial asthma and atopic dermatitis [
2]. ASIT vaccines are administered in small amounts, gradually increasing the doses due to the risk of anaphylactic shock, which makes it difficult to transit from IgE to IgG response.
Allergen extracts for ASIT are complex mixtures of proteins and non-protein compounds containing a large amount of impurities, therefore, side reactions, including an anaphylactic shock may occur [
3]. The difficulty in the manufacture of allergen extracts is the need to standardize the composition of ASIT preparations, which varies from batch to batch. To increase the effectiveness of ASIT and reduce the possibility of adverse reactions, the surface structure of allergens can be modified, including chemical modification or encapsulation of allergens in microgranules. Preparations with a modified allergen surface makes ASIT lose its ability to interact with IgE, therefore they can be used in doses many times higher than in the standard protocol [
4].
Currently, several possible ASIT mechanisms have been proposed. ASIT has a suppressive effect on the humoral and cellular components of an allergic reaction. After ASIT, the number of mast cells, neutrophils and eosinophils in the skin decreases, which leads to a decrease in the release of inflammatory mediators [
5,
6]. The most significant effect of ASIT is the formation of B cells synthesizing IgG4 and a gradual decrease in IgE titers, preventing a seasonal increase in IgE levels [
2].
Changing the surface structure of antigens reduces the interaction with IgE, so such preparations can be used in doses significantly higher than the standard ones. One of the methods of surface modification is the encapsulation of the antigen in a polymer shell obtained on the basis of chitosan and its derivatives. Zheng-Shun Wen et al. [
7] showed that chitosan nanoparticles can enhance cellular and humoral immune response and induce a balanced Th1/Th2 response. Additionally, a vaccine based on chitosan nanoparticles has been developed that significantly reduces inflammation of the respiratory tract in mice with bronchial asthma [
8]. Krishnendu Roy et al. showed that oral immunization with nanoparticles synthesized by the complexation of peanut allergen DNA with chitosan was effective in reducing anaphylactic reactions in mice, which makes it possible to use such a vaccine for the treatment of food allergies [
9]. None of these studies analyzed IgE production before and after ASIT.
In this work, core–shell Gal-MPs based on chitosan derivatives were developed for subcutaneous ASIT. The core contained recombinant Gal d1 protein immobilized on succinyl chitosan (SC) coated with the shell formed by quaternized chitosan (QC). Earlier, negatively charged core–shell chitosan-alginate nanoparticles of 500–700 nm in diameter able to prevent the allergen contact with mast cells, were developed [
10]. However, negative charge and small size of the particles resulted in a relatively low IgG titers. In this work, positively charged Gal-MPs which effectively induced IgG response and blocked IgE were developed. The developed core–shell structure contributed to the rapid and safe production of IgG antibodies against allergens.
2. Materials and Methods
2.1. Materials
Crab chitosan with MW 200 kDa and a degree of deacetylation (DD) of 96% (Bioprogress, Moscow region, Russia), and N-[(2-hydroxy-3-trimethylammonium)propyl] chitosan chloride (QC) with MW 25 kDa and a degree of substitution (SD) of 98%, (a gift from Dr. B. Shagdarova, Laboratory of biopolymer Engineering of FITZ Biotechnology RAS, Moscow, Russia) were used. Chitosan derivatives were purified via reprecipitation and dialysis. Succinic acid anhydride (Honewell, Lincolnshire, IL, USA), 2-hydroxypropane-1,2,3-tricarboxylic acid, acetic acid, hydrochloric acid (HimMed, Moscow, Russia), N-(3- Dimethylaminopropyl)-N′-ethylcarbodiimide hydrochloride (EDAC), azacitidin, dimethyl sulfoxide (DMSO), paraformaldehyde (PFA), Hoechst 33342, 3,3′5,5′-tetramethylbenzidine (TMB) (Merck KGaA, Darmstadt, Germany), Alexa Fluor 488™ phalloidin (ThermoScientific, Wltham, MA, USA), Mowiol 4.88 (Calbiochem, Nottingham, UK), rodamine B-COOH (a gift of Dr. Yu. Gracheva, Nizhni Novgorod State University, Russia) were used. The remaining reagents had analytical purity and were used without additional purification.
2.2. Chitosan Succinylation
To obtain SC, 2 g of Chi with MW 25 kDa, DD 98%, obtained by acid hydrolysis of chitosan 200 kDa and CD 96% [
11], was dissolved in 40 mL of 5% acetic acid followed by the addition of 160 mL of ethanol. The amount of succinic anhydride was calculated based on the ratio of
v/
v anhydride to amino groups (=20). Succinic anhydride was dissolved in a minimum volume of acetone and added to the Chi solution with stirring. The mixture was incubated overnight. The resulting gel was dissolved in distilled water and the pH was adjusted to 10 with 2 M NaOH. The resulting precipitate and solution were dialyzed against distilled water and lyophilized [
12,
13].
2.3. Physicochemical Characteristics of Chitosan Derivatives
The MW of chitosan derivatives were determined using high-performance gel-penetrating chromatography on the S 2100 Sykam chromatograph (Eresing, Germany) with a column (7.8 × 300 mm) Ultahydrogel-250 (Waters, Milford, MA, USA) and a pre-column (4 × 3 mm) GFC-4000 (Phenomenex, Torrance, CA, USA) [
14].
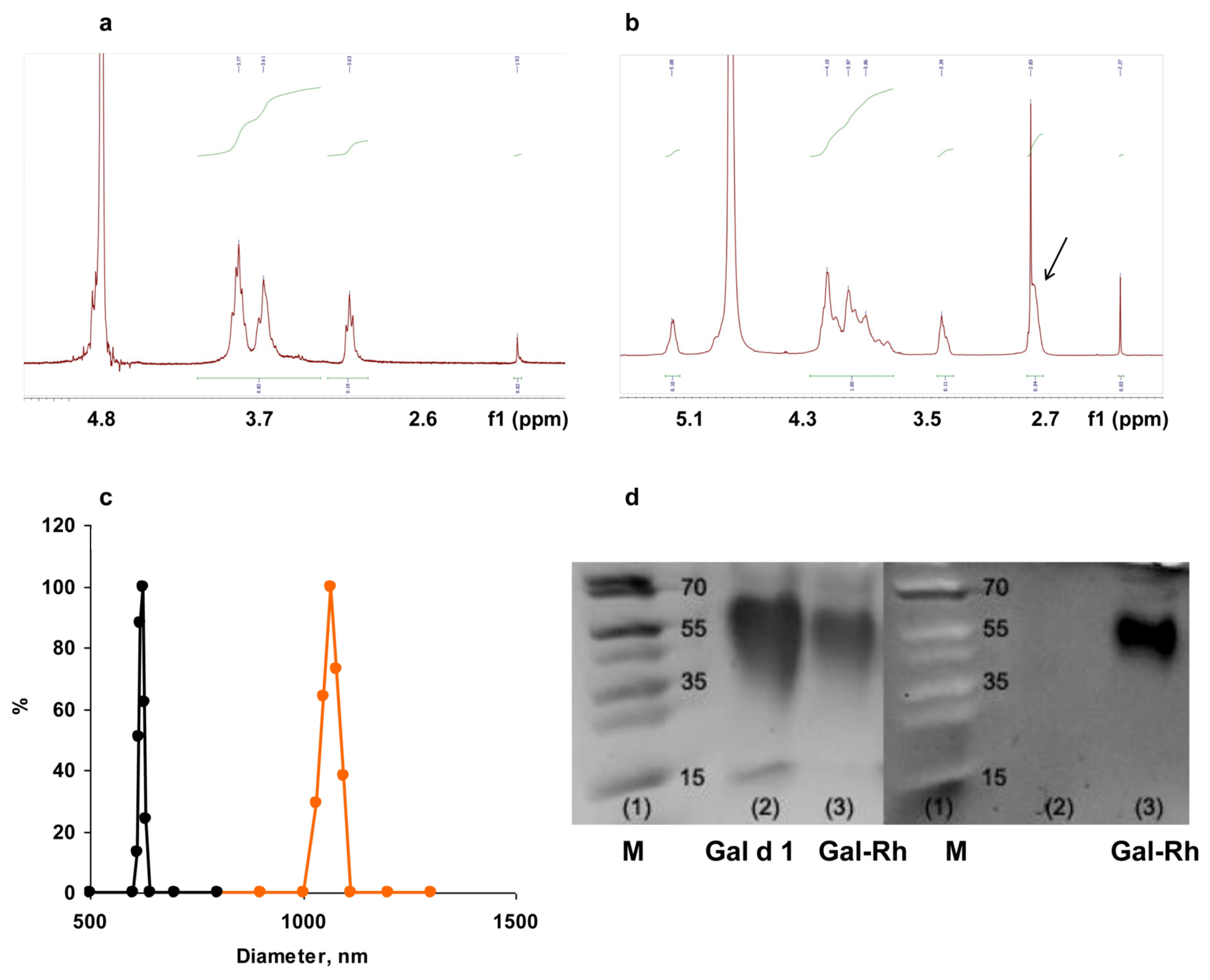
The DD of chitosan samples and the SD of SC were determined using proton magnetic resonance (1H-NMR). Proton spectra were recorded on a Bruker AMX 400 spectrometer (Camarillo, CA, USA). The samples were prepared in deuterated water, and 4,4-dimethyl-4-silapentane sulfonic acid was used as a standard.
2.4. Rhodamine Labeled Gal d1
Rodamine B-COOH (RhB-COOH), 0.8 mM in DMSO was mixed with 15 mM of EDAC in water, pH 6, and incubated for 20 min on ice with stirring. Gal d1, 5 mg, was dissolved in 200 µL of 0.05 M Na-bicarbonate buffer, pH 9.6 and mixed with the activated RhB-COOH. The reaction was carried out on ice overnight. The protein was then dialyzed three times against a phosphate-buffered solution (PBS) and freeze-dried.
2.5. Gel Electrophoresis
Gel electrophoresis was performed on 12% separating polyacrylamide gel and stained with Coomassie diamond blue R-250. The gels were documented on the Gel Doc EZ Imager device (BioRad Laboratories, Hercules, CA, USA).
2.6. Preparation of Core Particles using Citric Acid and EDAC
SC was dissolved in PBS, pH 7.2, at 2.5 mg/mL and left for 30 min with stirring to complete transparency. Then, 0.5 mg of Gal d1 or Gal-RhB was added. The ratio of antigen to SC was 1:5. Further, 0.6 mL of citric acid solution in water (4 mg/mL) with EDAC (8 mg/mL) was added drop by drop to the resulting solution with stirring to form core particles containing the conjugated allergen. The reaction mixture was stirred for 3 h at room temperature.
2.7. Preparation of Core Particles Using CaCl2
SC was dissolved in PBS, pH 7.2, to a concentration of 2.5 mg/mL, and then Gal d1 was added at a ratio of 5:1. MPs were formed by the dropwise addition of 85 µL of 1% CaCl2 solution was to 1 mL of the resulting solution. The reaction mixture was stirred for 3 h at room temperature.
2.8. Formation of Core–Shell Gal-MPs
Both types of core particles were coated with QC. To this end, 250 µL of QC solution per 1 mL of the core particles was added drop by drop by stirring for 30 min. During this time, a polyelectrolyte complex with a positive charge was formed. The resulting Gal-MPs were sterilized in a microwave oven (1 min in a water bath at maximum power). Sterility was confirmed in vitro.
2.9. Size and Charge of Gal-MPs
The size and charge of core and core–shell Gal-MPs was carried out via dynamic light scattering (DLS) using the 90 Plus Particle Size and ZetaPALS analyzer (Brookhaven, Vernon Hills, IL, USA) at a temperature of 25 °C, a fixed angle of 90° and a laser wavelength of 661 nm. The average values of the effective diameter and zeta potential were calculated from at least 5 independent measurements. The concentration of Gal d1 within the core and core–shell MPs was determined using spectrophotometer NanoDrop One (Thermo Scientific, Walthan, MA, USA).
2.10. Cell Cultures
Murine macrophage cells RAW264.7, fibroblasts L929, and human macrophage-like THP-1 cell line (IBCh collection, Moscow, Russia) were grown in RPMI-1640 supplemented with 10% fetal calf serum (FCS) and pen-strep-glut (complete culture medium) (all from PanEco, Moscow, Russian Federation) in CO2 incubator at 37 °C. Cells were passaged twice a week using Trypsin/EDTA solution (PanEco, Moscow, Russia).
2.11. MTT-Assay
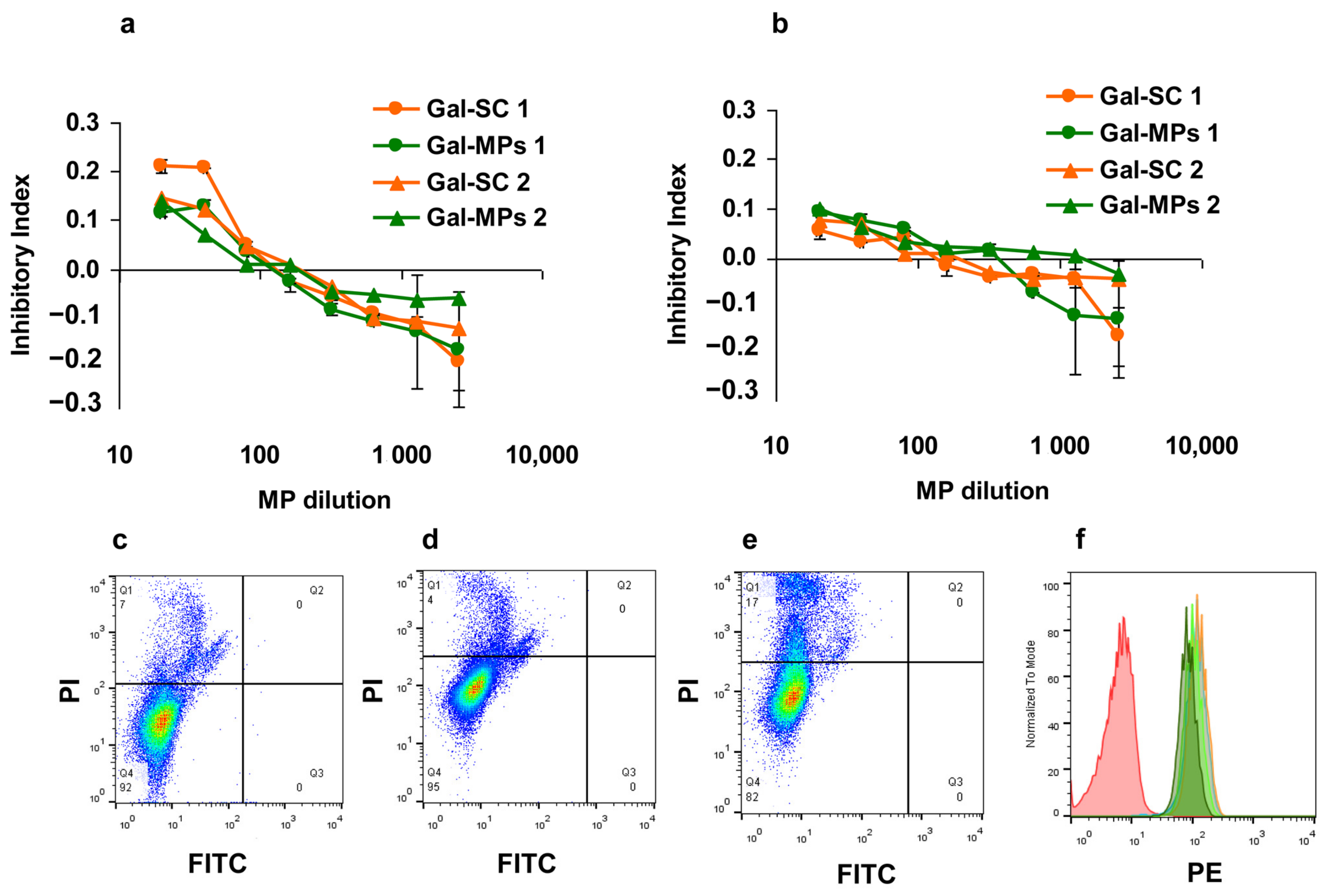
The cytotoxic effect of Gal-MPs was estimated using a standard 3-(4, 5-dimethyl-2-thiazolyl)-2, 5-diphenyl-2H-tetrazolium bromide (MTT, Sigma) assay. Briefly, different dilutions of Gal-MPs were prepared on a separate 96-well flat-bottom plate (Costar, Washington, WA, USA) and then transferred in 100 μL to the plates containing 5 × 103 cells/well. Non-treated cells served as controls. The plates were incubated for 72 h. For the last 3 h 10 µL of MTT (5 mg/mL) was added to each well. After the incubation, the culture medium was removed and 100 μL of DMSO was added to dissolve formazan crystals. Optical density was read on spectrophotometer Titertek (UK) at 540 nm. The results were analyzed using Excel package (Microsoft). The inhibition of proliferation (inhibition index, II) was calculated as II = [1 − (ODexperiment/ODcontrol)], where OD is the optical density. The experiments were repeated twice and average data are shown.
2.12. Bone Marrow Derived Macrophages and Dendritic Cells
BALB/c mice were euthanized by cervical dislocation. Bone marrow cells (BMCs) were obtained by flushing out femurs with PBS. Cells were cultured for 7 days at a density of 1.5–2 × 106/cm3 in complete culture medium supplemented with 10% of L929 supernatant as a source of differentiation factors. All non-adhesive cells were discarded, adhesive cells were trypsinized, washed, and used for the analysis.
2.13. Confocal Microscopy
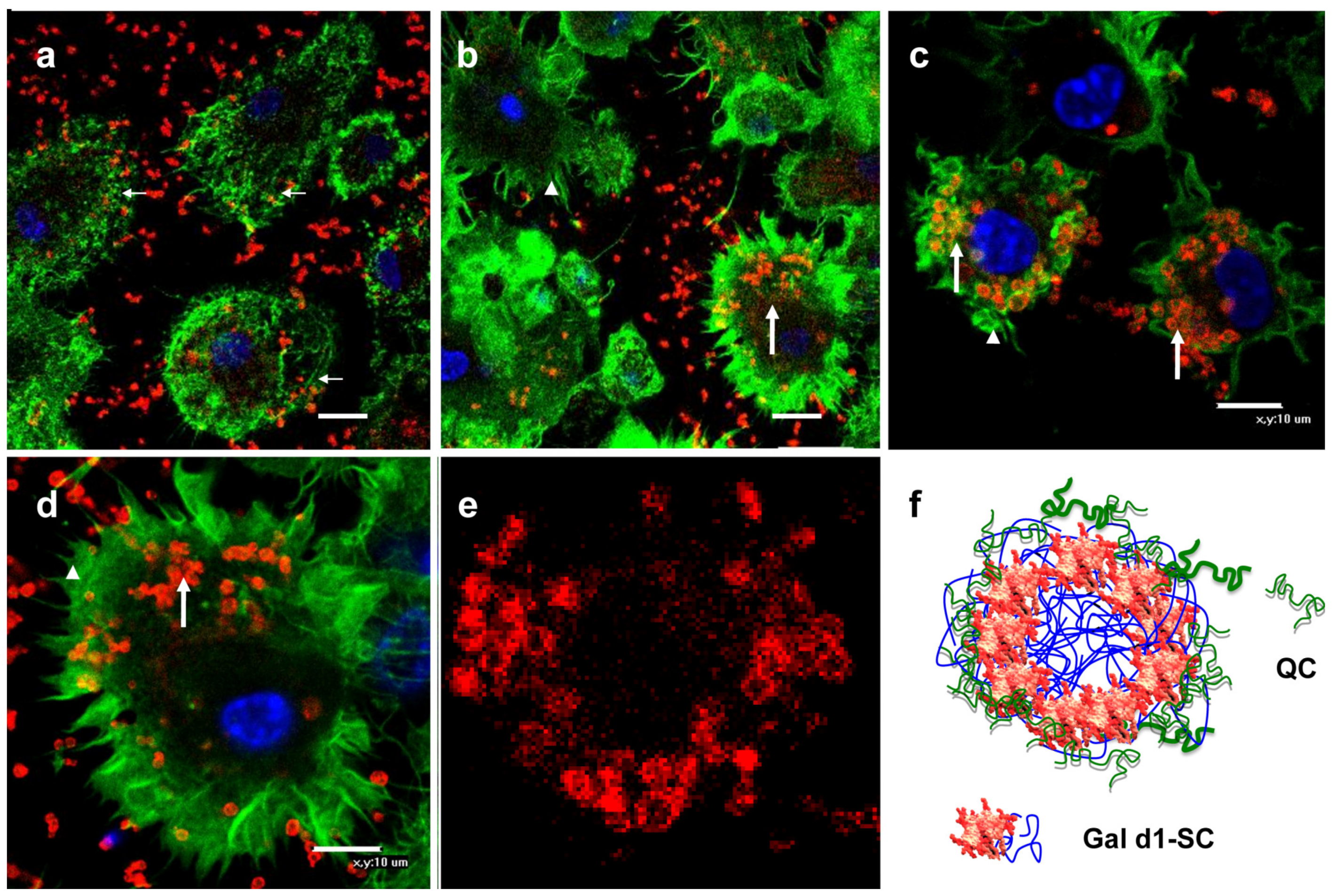
BMC macrophages, 105, were seeded onto sterile cover glasses (105/per well) and incubated in a CO2 incubator at 37 °C until the cells adhered. Rh-labeled Gal-MPs were added and incubated for 18 h at 37 °C. At the end of the incubation, the cells were fixed with 1% PFA and washed three times with PBS 0.01% Triton X100. Fixed and permeabilized BMCs were stained with phalloidin AlexaFluor 488 (Applied Biosystems, Foster City, CA, USA) and nuclear dye Hoechst 33342 (Merck KGaA, Darmstadt, Germany) for 1 h. Cells were polymerized with Mowiol (Calbiochem, Nottingham, UK) and analyzed using a TE 2000 Eclipse confocal microscope (Nikon, Tokyo, Japan).
2.14. Flow Cytometry
To analyze cytotoxicity and Gal–MPs interaction with the antigen presenting cells via flow cytometry, human macrophage-like THP-1 cell line was used. Gal-Rh-MPs were incubated with 106 cells/well for 18 h and stained with 50 μg/mL of propidium iodide (Sigma, Merck KGaA, Darmstadt, Germany), incubated for 15 min, and analyzed. The analysis was carried out using FACScan flow cytometer (BD, Franklin Lakes, NJ, USA). Calculation of the results was carried out using the program FlowJo (USA). BMC macrophages were characterized by the expression of CD11b and CD11c markers using specific antibodies (Biolegend, San Diego, CA, USA).
2.15. Mice
Female BALB/c mice (25 ± 5 g) were obtained from the laboratory animal nursery “Pushchino”, Moscow region. The mice were housed in plastic cages, 10–12 mice per cage, under conventional minimal pathogen conditions. They were kept in a 12 h light/dark cycle at room temperature and fed ad libitum. Mice were euthanized by cervical dislocation. The research protocol was approved by the Ethical Committee of the Institute of Bioorganic Chemistry of the Russian Academy of Sciences, Protocols No. 232 (2018) and No. 327 (2021).
2.16. ASIT
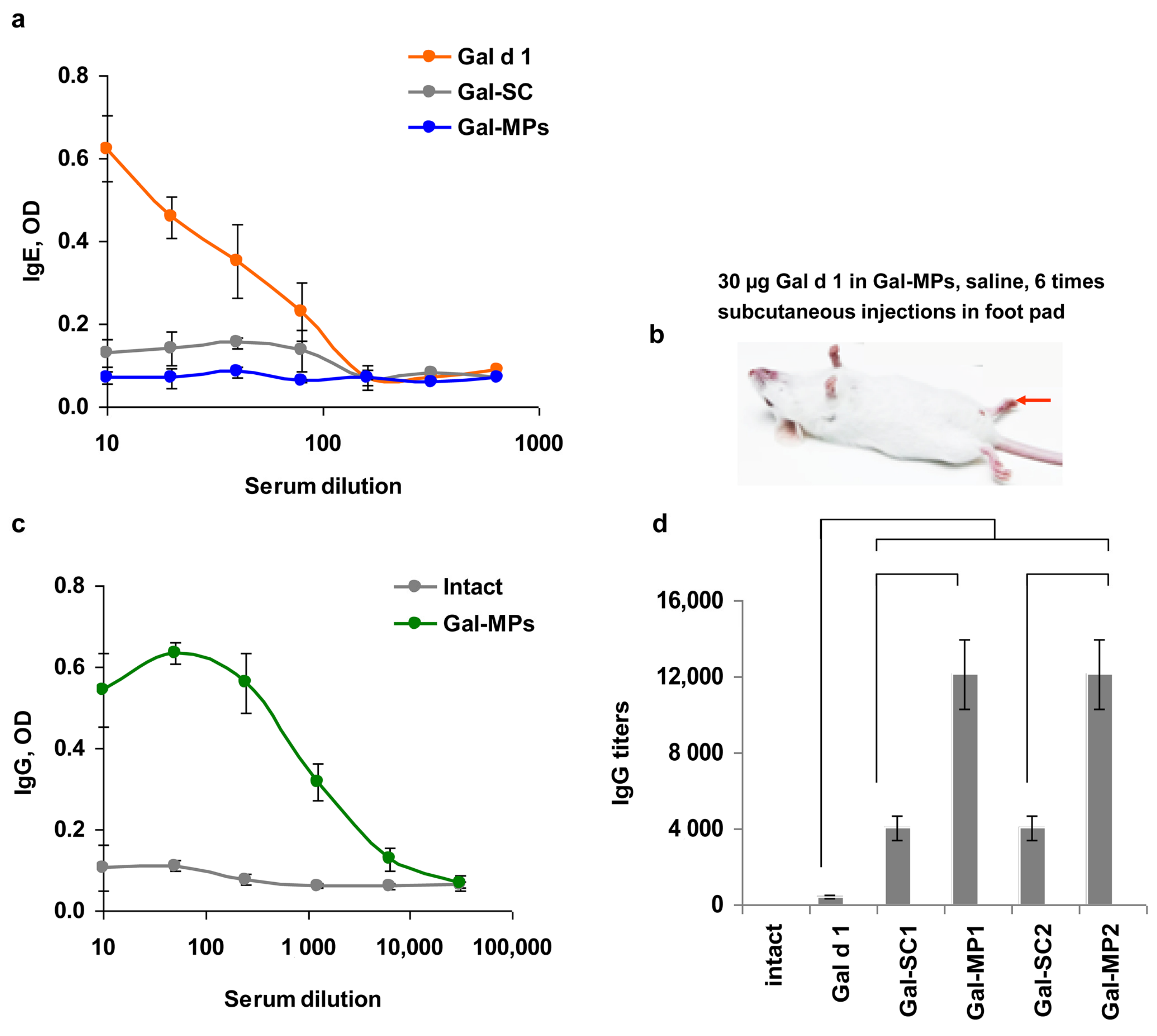
Mice (n = 30) were divided into 6 groups and were injected subcutaneously in a foot pad 6 times with 2–3 day intervals: intact mice (n = 5) were injected with PBS; mice from group 1 (n = 5) were injected with Gal-d1; mice from group 2 (n = 5) were injected with Gal-SC 1; mice from group 3 (n = 5) were injected with Gal-MPs 1; mice from group 4 (n = 5) were injected with Gal-SC 2; and mice from group 5 (n = 5) were injected with Gal-MPs 2. The injection contained 30 µg of Gal d1. Blood was collected from the orbital sinus using isofluran anesthesia before and after immunization. Serum was collected and stored at -20 °C before use.
2.17. Allergy Model
A low-dose allergy model was used to induce IgE to Gal d1 [
10]. Briefly, mice (n = 5) were injected into the withers 9 times with 100 ng/mouse of Gal d1 in saline. To estimate the ASIT effect, a group of mice (n = 5) was previously vaccinated with Gal-MPs 1, as described above. One month after vaccination, mice were treated to induce allergic IgE response. To estimate the safety of ASIT, mice with Gal d1-induced IgE were injected with Gal-MPs. Local reaction was analyzed based on body temperature and skin rash reaction.
2.18. Enzyme Immunoassay (ELISA)
ELISA was performed on 96-well microtitration plates (Costar, Washington, WA, USA) coated with 5 µg/mL of Gal d1 or Gal-MPs in 0.05 M Na-bicarbonate buffer, with pH 9.6. The plates were incubated overnight at +4 °C. The plates were washed between each incubations with PBS 0.05% Tween-20 (PBS-T). All incubations with serum and specific antibodies were conducted in PBS-T with 5% non-fat dry milk at room temperature for 1 h. Anti-mouse IgG-HRP or IgE-HRP (Abcam, Cambridge, UK) levels were estimated using TMB substrate. The optical density was measured using a spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) at 450 nm with subtraction of the optical density at 620 nm. The number of antibodies was evaluated as titers corresponding to the maximum serum dilutions, at which the optical density (OD) was 3 SD deviations higher than the average background.
2.19. Histology
Withers tissue samples were fixed in 4% paraformaldehyde and embedded in paraffin. Samples were cut into sections with a thickness of 4 μm using Leica RM 2145 RTS microtome (Leica Biosystems, Amsterdam, The Netherlands). After dewaxing of tissue sections, they were stained with H&E according to the manufacturer’s protocols. Then, the sections were dehydrated, clarified, and enclosed under a cover glass using the Consul-Mount histological medium.
2.20. Statistical Analysis
Graphs were created using MS Excel software. The data are represented as mean ± SD of at least two independent experiments or as one representative experiment. The parametric t-test was used to compare the experimental groups. Levels of p < 0.05 were considered statistically significant.
4. Discussion
Chitosan is an aminopolysaccharide obtained from chitin via N-deacetylation. It has free amino groups (-NH
2), which opens up wide possibilities for chitosan modification. SC derivatives are obtained via acylation with succinic anhydride in an organo-aqueous medium, containing1% acetic acid and acetone, to dissolve the anhydride. The reaction proceeds with the opening of the anhydride ring. The SD of an acyl residue depends on the ratio of reagents in the reaction [
18]. SC is used in various drug delivery systems to deliver insulin, anticancer agents, genetic material [
19,
20,
21], etc. Methods to obtain chitosan-based nanoparticles include ionic gelation, emulsion, reverse micellar method, coacervation, deposition, nanoprecipitation, and sieving method [
22]. Chitosan-based nanoparticles are good carriers due to properties, such as hydrophilicity, non-toxicity, biodegradability, biocompatibility and bioadhesiveness [
23]. In this work, the allergen Gal d1 ovomucoid with MW 28 kDa, which is a major hen egg’s allergen [
24], was used to be encapsulated into MPs. Core Gal-SCs were obtained using either a crosslinking agent citric acid activated using carbodiimide or CaCl
2 precipitation. As the results of the confocal microscopy showed, Gal d1 was found on the surface of SC particles. This packaging may not completely cover some epitopes on the surface of the particles sufficient for their recognition by IgE. Therefore, the next step was to cover Gal d1 by the shell. To develop the shell, we used positively charged chitosan derivative QC with a high SD. SC and QC formed a polyelectrolyte complex due to electrostatic interactions.
Biomedical applications of any preparation requires high safety requirements. Previously, it has been shown that the toxicity of chitosan and its derivatives is determined mostly by their charge. Regardless of the degree of acetylation, molecular weight, and hydrophobicity, only highly positively charged chitosan derivatives, namely quaternized one with SD more than 80%, at high concentration inhibited the proliferation of cells [
25]. Multiple studies show that chitosan-based materials mainly are low toxic to cells [
26,
27]. However, some toxicity of positively charged MPs can serve as an additional adjuvant for the tissue resident antigen-presenting cells [
28]. Multiple studies have shown that chitosan-based delivery systems stimulate antibody production, cytokine synthesis, and functional effects of proteins, peptides, and DNA encapsulated into nano- and microparticles [
7,
8,
9,
28]. Our results supported these findings and showed the induction of IgG response to Gal d1 in mice immunized by six injections of Gal-MPs. Gal-SC particles were less effective in IgG induction in a comparison with core–shell Gal-MPs. This can be a result of the positive charge of Gal-MPs, however, the size of the particles also can play a role.
The specific characteristic of Gal-MPs was their bi-layer structure designed to cover possible allergen IgE binding sites. Analysis of IgE binding showed that even core Gal-SC particles did not bind IgE, however, we think that for a better safety core–shell particles are preferable.
The size of the delivery system depends on the planned application. The delivery vehicle must be small (50–150 nm) for the intravenous injections or tissue injections, such as the eye, where the cargo must be released within a prolonged time [
29]. Contrary to it, vaccines are designed to be quickly engulfed by the resident tissue macrophages and dendritic cells. Evolutionally macrophages better phagocyte large objects while dendritic cells are able to uptake soluble antigens by endocytosis. We showed that larger Gal-MPs were better delivered to the antigen presenting cells than Gal-SC, resulting in a lower IgG production.
The last part of our work was devoted to the verification that the vaccination with the encapsulated allergens can modify allergic response. It has been shown that the preventive vaccination with Gal-MPs can abrogate IgE formation to Gal d1.

 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}