
‘Pera’ Orange and ‘Moro’ Blood Orange Juice Improves Oxidative Stress and Inflammatory Response Biomarkers and Modulates the Gut Microbiota of Individuals with Insulin Resistance and Different Obesity Classes

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Population
2.2. Intervention
2.3. Study Protocol
2.4. Anthropometric and Dietary Intake Measurement
2.5. Biochemical Parameters
2.6. Plasma Antioxidant Activity
2.7. Superoxide Dismutase (SOD) and Glutathione Peroxidase (GPx) Activity
2.8. Urinary 8-Isoprostane and 8-Hydroxy-2′-deoxyguanosine (8-OHdG) Content
2.9. Inflammatory Biomarkers
2.10. Identification and Quantification of Urine Metabolites
2.11. Gut Microbiota Profiling
2.12. Short-Chain Fatty Acids (SCFAs)
2.13. Statistical Analysis
3. Results
3.1. Quantification of Urinary Flavanones, Metabolites, and Phenolic Acids after Orange Juice Intake
3.2. Effect of Orange Juice Intake on Anthropometric and Biochemical Parameters
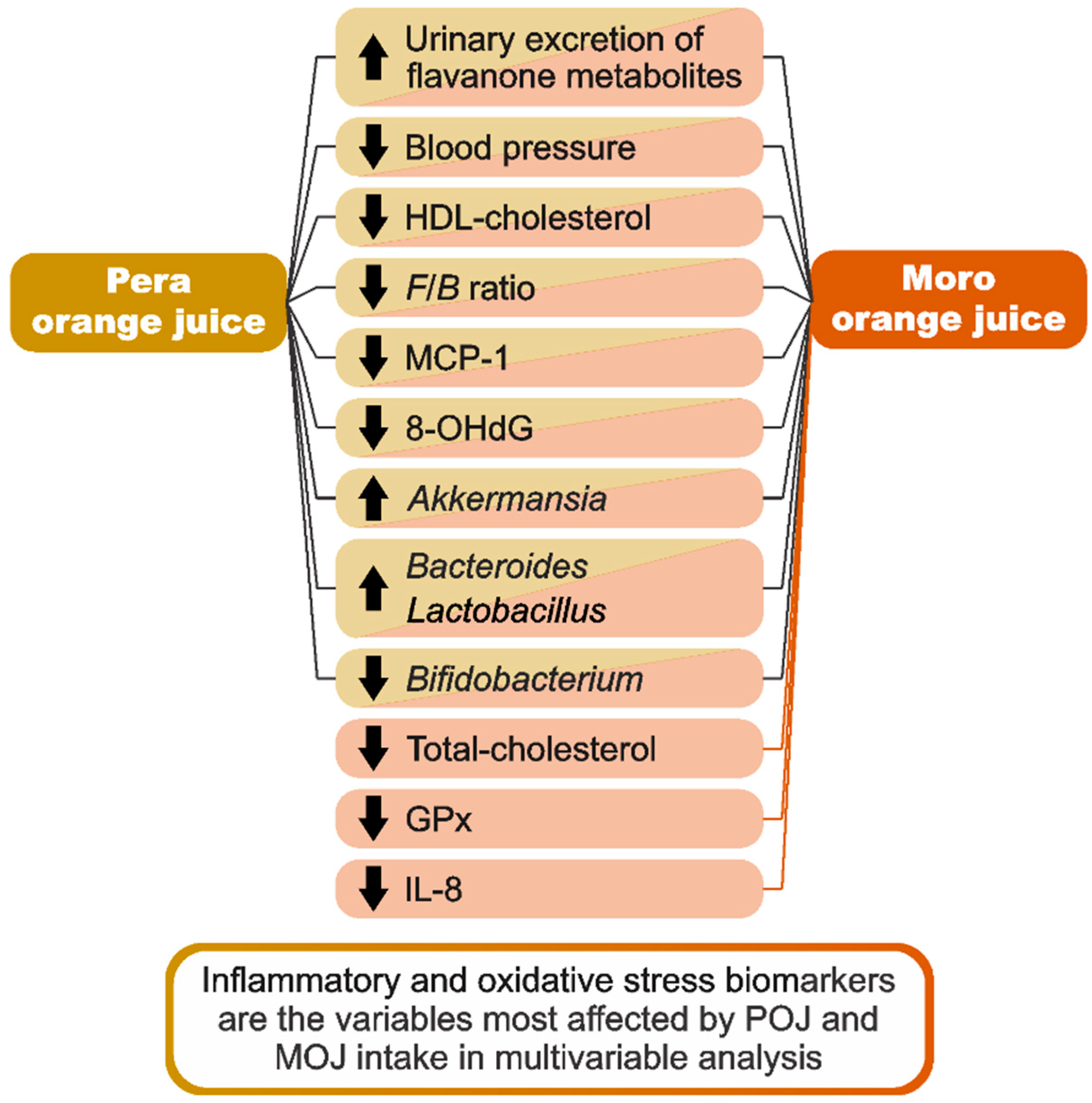
3.3. Inflammatory and Oxidative Stress Biomarker Response after Orange Juice Intake
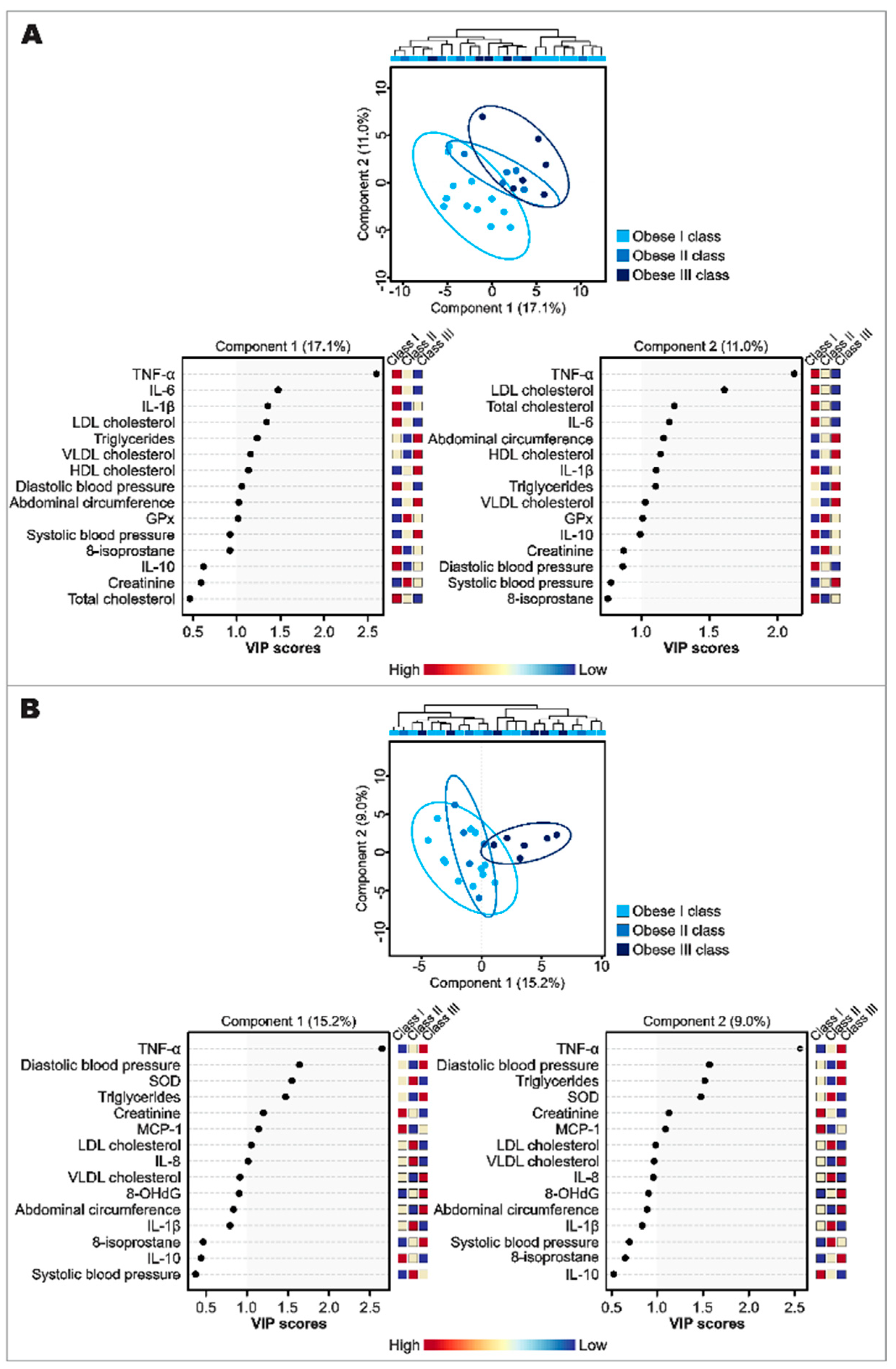
3.4. Effect of Obesity and Insulin Resistance on Gut Microbiota Profile
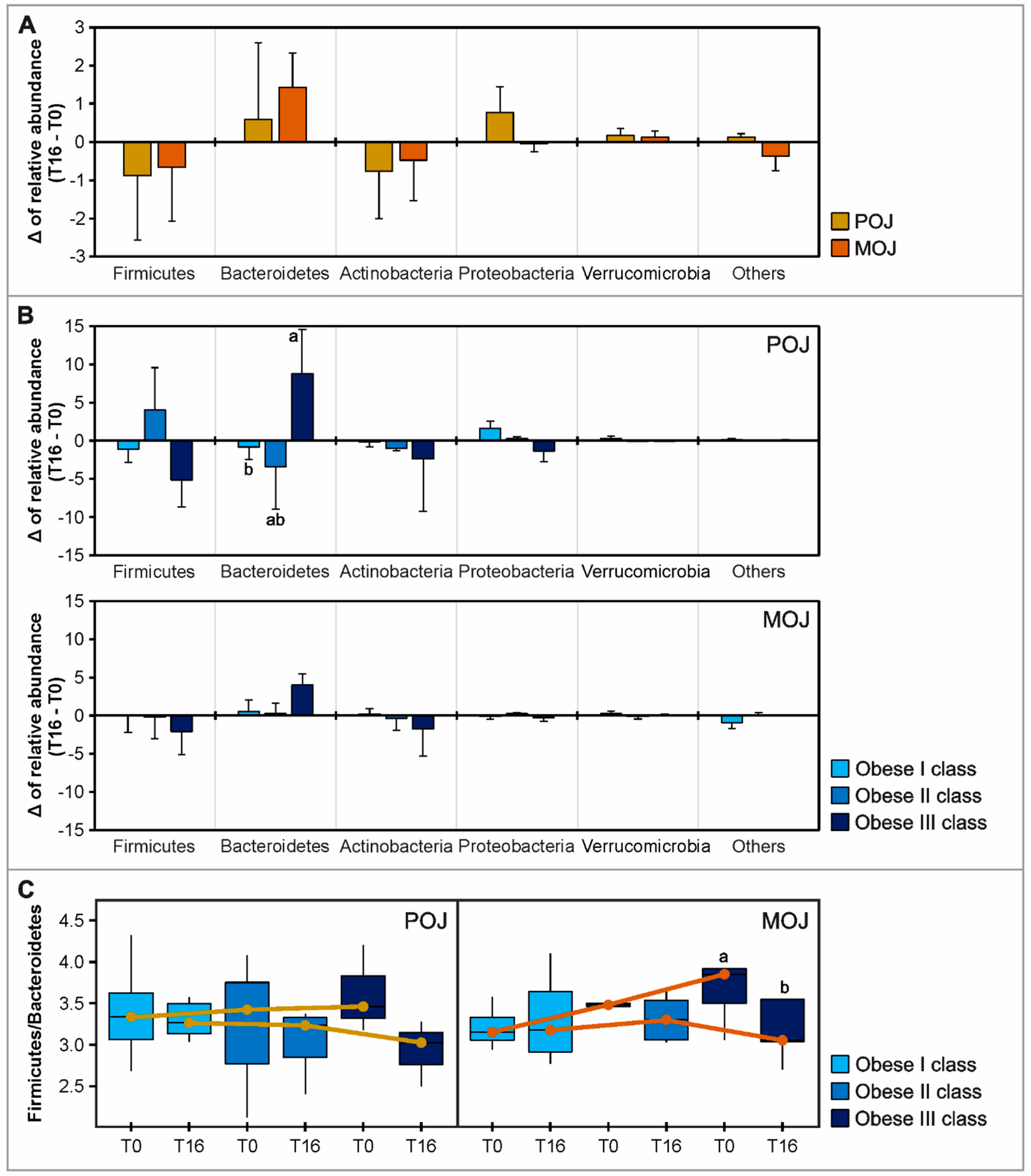
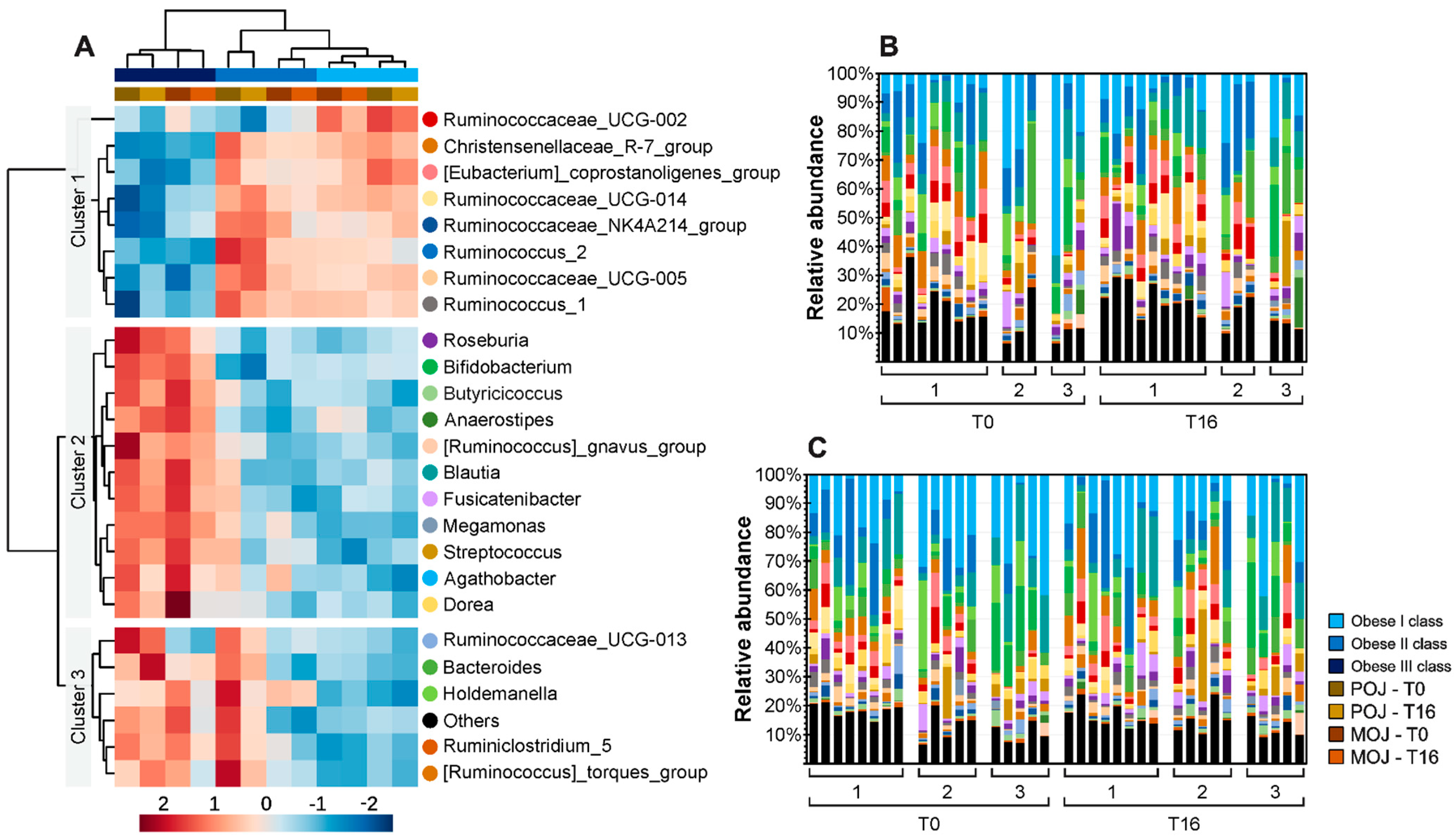
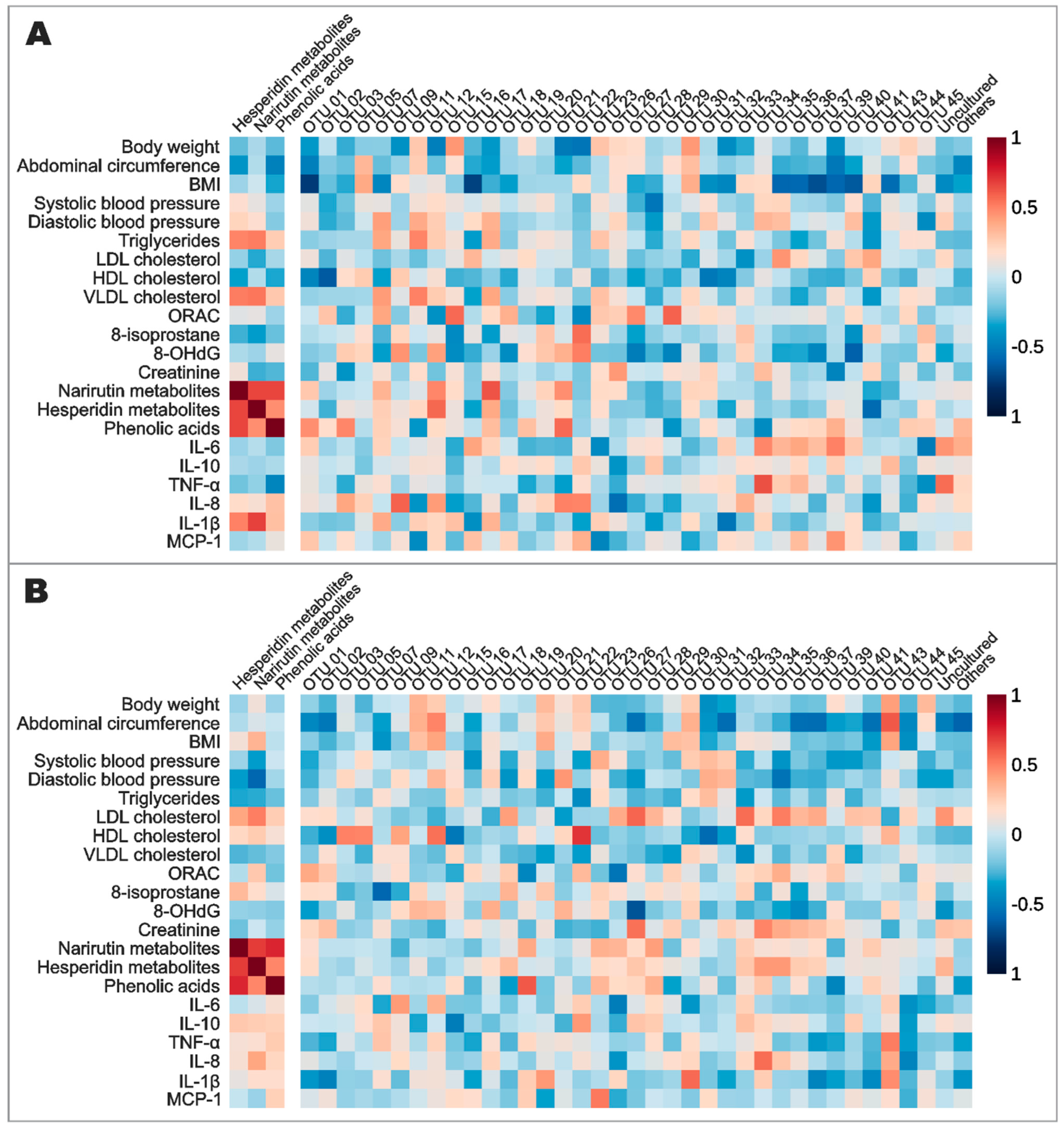
3.5. Effects of Orange Juice Intake on Gut Microbiota Profile and Short-Chain Fatty Acids
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chooi, Y.C.; Ding, C.; Magkos, F. The epidemiology of obesity. Metabolism 2019, 92, 6–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blüher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef]
- Karczewski, J.; Śledzińska, E.; Baturo, A.; Jończyk, I.; Maleszko, A.; Samborski, P.; Begier-Krasińska, B.; Dobrowolska, A. Obesity and inflammation. Eur. Cytokine Netw. 2018, 29, 83–94. [Google Scholar] [CrossRef] [PubMed]
- Hruby, A.; Manson, J.E.; Qi, L.; Malik, V.S.; Rimm, E.B.; Sun, Q.; Willett, W.C.; Hu, F.B. Determinants and Consequences of Obesity. Am. J. Public Health 2016, 106, 1656–1662. [Google Scholar] [CrossRef] [PubMed]
- Esteve, E.; Ricart, W.; Fernández-Real, J.M. Gut microbiota interactions with obesity, insulin resistance and type 2 diabetes: Did gut microbiote co-evolve with insulin resistance? Curr. Opin. Clin. Nutr. Metab. Care 2011, 14, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Saad, M.J.A.; Santos, A.; Prada, P.O. Linking Gut Microbiota and Inflammation to Obesity and Insulin Resistance. Physiology 2016, 31, 283–293. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K.; Cabral, C.; Kumar, R.; Ganguly, R.; Rana, H.K.; Gupta, A.; Lauro, M.R.; Carbone, C.; Reis, F.; Pandey, A.K. Beneficial effects of dietary polyphenols on gut microbiota and strategies to improve delivery efficiency. Nutrients 2019, 11, 2216. [Google Scholar] [CrossRef] [Green Version]
- Corrêa, T.A.F.; Rogero, M.M.; Hassimotto, N.M.A.; Lajolo, F.M. The Two-Way Polyphenols-Microbiota Interactions and Their Effects on Obesity and Related Metabolic Diseases. Front. Nutr. 2019, 6, 188. [Google Scholar] [CrossRef] [Green Version]
- Cardona, F.; Andrés-Lacueva, C.; Tulipani, S.; Tinahones, F.J.; Queipo-Ortuño, M.I. Benefits of polyphenols on gut microbiota and implications in human health. J. Nutr. Biochem. 2013, 24, 1415–1422. [Google Scholar] [CrossRef] [Green Version]
- Gwiazdowska, D.; Juś, K.; Jasnowska-Małecka, J.; Kluczyńska, K. The impact of polyphenols on Bifidobacterium growth. Acta Biochim. Pol. 2015, 62, 895–901. [Google Scholar] [CrossRef]
- Panche, A.N.; Diwan, A.D.; Chandra, S. Flavonoids: An overview. J. Nutr. Sci. 2016, 5, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milenkovic, D.; Deval, C.; Dubray, C.; Mazur, A.; Morand, C. Hesperidin Displays Relevant Role in the Nutrigenomic Effect of Orange Juice on Blood Leukocytes in Human Volunteers: A Randomized Controlled Cross-Over Study. PLoS ONE 2011, 6, e26669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghanim, H.; Sia, C.L.; Upadhyay, M.; Korzeniewski, K.; Viswanathan, P.; Abuaysheh, S.; Mohanty, P.; Dandona, P. Orange juice neutralizes the proinflammatory effect of a high-fat, high-carbohydrate meal and prevents endotoxin increase and Toll-like receptor expression. Am. J. Clin. Nutr. 2010, 91, 940–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anacleto, S.L.; Milenkovic, D.; Kroon, P.A.; Needs, P.W.; Lajolo, F.M.; Hassimotto, N.M.A. Citrus flavanone metabolites protect pancreatic-β cells under oxidative stress induced by cholesterol. Food Funct. 2020, 11, 8612–8624. [Google Scholar] [CrossRef]
- Morand, C.; Dubray, C.; Milenkovic, D.; Lioger, D.; Martin, J.F.; Scalbert, A.; Mazur, A. Hesperidin contributes to the vascular protective effects of orange juice: A randomized crossover study in healthy volunteers. Am. J. Clin. Nutr. 2011, 93, 73–80. [Google Scholar] [CrossRef] [Green Version]
- Rangel-Huerta, O.D.; Aguilera, C.M.; Martin, M.V.; Soto, M.J.; Rico, M.C.; Vallejo, F.; Tomas-Barberan, F.; Perez-de-la-Cruz, A.J.; Gil, A.; Mesa, M.D. Normal or High Polyphenol Concentration in Orange Juice Affects Antioxidant Activity, Blood Pressure, and Body Weight in Obese or Overweight Adults. J. Nutr. 2015, 145, 1808–1816. [Google Scholar] [CrossRef] [Green Version]
- Fraga, L.N.; Coutinho, C.P.; Rozenbaum, A.C.; Tobaruela, E.C.; Lajolo, F.M.; Hassimotto, N.M.A. Blood pressure and body fat % reduction is mainly related to flavanone phase II conjugates and minor extension by phenolic acid after long-term intake of orange juice. Food Funct. 2021, 12, 11278–11289. [Google Scholar] [CrossRef]
- Murphy, M.M.; Barrett, E.C.; Bresnahan, K.A.; Barraj, L.M. 100 % Fruit juice and measures of glucose control and insulin sensitivity: A systematic review and meta-analysis of randomised controlled trials. J. Nutr. Sci. 2017, 6, e59. [Google Scholar] [CrossRef] [Green Version]
- Food and Agriculture Organization of the United Nations. Citrus Fruit Statistical Compendium 2020; Food and Agriculture Organization of the United Nations: Rome, Italy, 2021. [Google Scholar]
- Carnauba, R.A.; Sarti, F.M.; Hassimotto, N.M.A.; Lajolo, F.M. Estimated polyphenol intake and major food sources of the Brazilian population: Changes between 2008–2009 and 2017–2018. Br. J. Nutr. 2022, 7, 1–8. [Google Scholar] [CrossRef]
- Muscatello, M.R.A.; Zoccali, R.A.; Bruno, A. Chapter 11—Citrus Fruit Polyphenols and Flavonoids: Applications to Psychiatric Disorders. Polyphenols: Mechanisms of Action in Human Health and Disease; Academic Press: Cambridge, MA, USA, 2018; pp. 119–131. [Google Scholar]
- Acquaviva, R.; Russo, A.; Galvano, F.; Galvano, G.; Barcellona, M.L.; Volti, G.L.; Vanella, A. Cyanidin and cyanidin 3-O-beta-D-glucoside as DNA cleavage protectors and antioxidants. Cell Biol. Toxicol. 2003, 19, 243–252. [Google Scholar] [CrossRef]
- Shih, P.H.; Yeh, C.T.; Yen, G.C. Anthocyanins induce the activation of phase II enzymes through the antioxidant response element pathway against oxidative stress-induced apoptosis. J. Agric. Food Chem. 2007, 55, 9427–9435. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Qin, J.; Xie, F.; Zhou, K.; Xi, W. Red light-transmittance bagging promotes carotenoid accumulation through xanthophylls esterification during the ripening of blood orange fruit. Food Chem. 2023, 404, 134578. [Google Scholar] [CrossRef] [PubMed]
- Geloneze, B.; Repetto, E.M.; Geloneze, S.R.; Tambascia, M.A.; Ermetice, M.N. The threshold value for insulin resistance (HOMA-IR) in an admixtured population IR in the Brazilian Metabolic Syndrome Study. Diabetes Res. Clin. Pract. 2006, 72, 219–220. [Google Scholar] [CrossRef]
- Geloneze, B.; Vasques, A.C.J.; Stabe, C.F.C.; Pareja, J.C.; Rosado, L.E.F.P.L.; Queiroz, E.C.; Tambascia, M.A. HOMA1-IR and HOMA2-IR indexes in identifying insulin resistance and metabolic syndrome: Brazilian Metabolic Syndrome Study (BRAMS). Arq. Bras. Endocrinol. Metab. 2009, 53, 281–287. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Obesity: Preventing and managing the global epidemic, Report of a World Health Organization Consultation. WHO Obesity Technical Report Series; World Health Organization: Geneva, Switzerland, 2000; Volume 894.
- Prior, R.L.; Hoang, H.; Gu, L.; Wu, X.; Bacchiocca, M.; Howard, L.; Hampsch-Woodill, M.; Huang, D.; Ou, B.; Jacob, R. Assays for Hydrophilic and Lipophilic Antioxidant Capacity (oxygen radical absorbance capacity (ORACFL)) of Plasma and Other Biological and Food Samples. J. Agric. Food Chem. 2003, 51, 3273–3279. [Google Scholar] [CrossRef] [PubMed]
- Ewing, J.F.; Janero, D.R. Microplate superoxide dismutase assay employing a nonenzymatic superoxide generator. Anal. Biochem. 1995, 232, 243–248. [Google Scholar] [CrossRef] [PubMed]
- Flohé, L.; Günzler, W.A. Assays of glutathione peroxidase. Methods Enzymol. 1984, 105, 114–121. [Google Scholar]
- Nishioka, A.; Tobaruela, E.C.; Fraga, L.N.; Tomás-Barberán, F.A.; Lajolo, F.M.; Hassimotto, N.M.A. Stratification of Volunteers According to Flavanone Metabolite Excretion and Phase II Metabolism Profile after Single Doses of ‘Pera’ Orange and ‘Moro’ Blood Orange Juices. Nutrients 2021, 13, 473. [Google Scholar] [CrossRef]
- Christoff, A.P.; Sereia, A.F.R.; Boberg, D.R.; Moraes, L.R.V.; Oliveira, L.F.V. Bacterial Identification through Accurate Library Preparation and High-Throughput Sequencing; Neoprospecta Microbiome Technologies: Florianopolis, Brazil, 2017; pp. 1–5. [Google Scholar]
- Wang, Y.; Qian, P.Y. Conservative Fragments in Bacterial 16S rRNA Genes and Primer Design for 16S Ribosomal DNA Amplicons in Metagenomic Studies. PLoS ONE 2009, 4, e7401. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Huntley, J.; Fierer, N.; Owens, S.M.; Betley, J.; Fraser, L.; Bauer, M. Ultra-high-throughput microbial community analysis on the Illumina HiSeq and MiSeq platforms. ISME J. 2012, 6, 1621–1624. [Google Scholar] [CrossRef] [Green Version]
- Christoff, A.P.; Cruz, G.N.F.; Sereia, A.F.R.; Yamanaka, L.E.; Silveira, P.P.; Oliveira, L.F.V. End-to-end assessment of fecal bacteriome analysis: From sample processing to DNA sequencing and bioinformatics results. BioRxiv 2020, 1–22. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Kaehler, B.D.; Rideout, J.R.; Dillon, M.; Bolyen, E.; Knight, R.; Huttley, G.A.; Caporaso, J.G. Optimizing taxonomic classification of marker-gene amplicon sequences with QIIME 2’s q2-feature-classifier plugin. Microbiome 2018, 6, 90. [Google Scholar] [CrossRef] [PubMed]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Menezes, E.W.; Dan, M.C.T.; Cardenette, G.H.L.; Goñi, I.; Bello-Pérez, L.A.; Lajolo, F.M. In Vitro Colonic Fermentation and Glycemic Response of Different Kinds of Unripe Banana Flour. Plant Foods Hum. Nutr. 2010, 65, 379–385. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Chong, J.; Zhou, G.; Morais, D.A.L.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the gap between raw spectra and functional insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef] [PubMed]
- Chong, J.; Liu, P.; Zhou, G.; Xia, J. Using MicrobiomeAnalyst for comprehensive statistical, functional, and meta-analysis of microbiome data. Nat. Protoc. 2020, 15, 799–821. [Google Scholar] [CrossRef]
- Shannon, C.E. A Mathematical Theory of Communication. Bell Syst. Tech. J. 1948, 27, 379–423. [Google Scholar]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Institute of Medicine. Dietary Reference Intakes for Energy, Carbohydrate, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids; The National Academies Press: Washington, DC, USA, 2005. [Google Scholar]
- Moreira, V.; Brasili, E.; Fiamoncini, J.; Marini, F.; Miccheli, A.; Daniel, H.; Lee, J.J.H.; Hassimotto, N.M.A.; Lajolo, F.M. Orange juice affects acylcarnitine metabolism in healthy volunteers as revealed by a mass-spectrometry based metabolomics approach. Food Res. Int. 2018, 107, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Rampersaud, G.C.; Valim, M.F. 100% citrus juice: Nutritional contribution, dietary benefits, and association with anthropometric measures. Crit. Rev. Food Sci. Nutr. 2017, 57, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietar (Poly)phenolics in Human Health: Structures, Bioavailability, and Evidence of Protective Effects Against Chronic Diseases. Antioxid. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azzini, E.E.; Venneria, E.; Ciarapica, D.; Foddai, M.S.; Intorre, F.; Zaccaria, M.; Maiani, F.; Palomba, L.; Barnaba, L.; Tubili, C. Effect of Red Orange Juice Consumption on Body Composition and Nutritional Status in Overweight/Obese Female: A Pilot Study. Oxid. Med. Cell Longev. 2017, 2017, 1672567. [Google Scholar] [CrossRef] [Green Version]
- Lima, A.C.D.; Cecatti, C.; Fidélix, M.P.; Adorno, M.A.T.; Sakamoto, I.K.; Cesar, T.B.; Sivieri, K. Effect of Daily Consumption of Orange Juice on the Levels of Blood Glucose, Lipids, and Gut Microbiota Metabolites: Controlled Clinical Trials. J. Med. Food 2019, 22, 202–210. [Google Scholar] [CrossRef]
- Constans, J.; Bennetau-Pelissero, C.; Martin, J.; Rock, E.; Mazur, A.; Bedel, A.; Morand, C.; Bérard, A.M. Marked antioxidant effect of orange juice intake and its phytomicronutrients in a preliminary randomized cross-over trial on mild hypercholesterolemic men. Clin. Nutr. 2015, 34, 1093–1100. [Google Scholar] [CrossRef]
- Ribeiro, C.; Dourado, G.; Cesar, T. Orange juice allied to a reduced-calorie diet results in weight loss and ameliorates obesity-related biomarkers: A randomized controlled trial. Nutrition 2017, 38, 13–19. [Google Scholar] [CrossRef]
- Briskey, D.; Malfa, G.A.; Rao, A. Effectiveness of “Moro” Blood Orange Citrus sinensis Osbeck (Rutaceae) Standardized Extract on Weight Loss in Overweight but Otherwise Healthy Men and Women—A Randomized Double-Blind Placebo-Controlled Study. Nutrients 2022, 14, 427. [Google Scholar] [CrossRef]
- Bondia-Pons, I.; Ryan, L.; Martinez, J.A. Oxidative stress and inflammation interactions in human obesity. J. Physiol. Biochem. 2012, 68, 701–711. [Google Scholar] [CrossRef]
- Diretrizes da Sociedade Brasileira de Diabetes (2019–2020); Editora Clannad: São Paulo, Brazil, 2019.
- Styskal, J.; Remmen, H.V.; Richardson, A.; Salmon, A.B. Oxidative stress and diabetes: What can we learn about insulin resistance from antioxidant mutant mouse models? Free Radic. Biol. Med. 2012, 52, 46–58. [Google Scholar] [CrossRef] [Green Version]
- Khan, M.K.; Zill-E-Huma; Dangles, O. A comprehensive review on flavanones, the major citrus polyphenols. J. Food Compos. Anal. 2014, 33, 85–104. [Google Scholar] [CrossRef]
- Ley, R.E.; Bäckhed, F.; Turnbaugh, P.; Lozupone, C.A.; Knight, R.D.; Gordon, J.I. Obesity alters gut microbial ecology. Proc. Natl. Acad. Sci. USA 2005, 102, 11070–11075. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, A.P.D.; Pereira, A.G.; Todo, M.C.; Fujimori, A.S.S.; Santos, P.P.; Dantas, D.; Fernandes, A.A.; Zanati, S.G.; Hassimotto, N.M.A.; Zornoff, L.A.M.; et al. Pera orange (Citrus sinensis) and Moro orange (Citrus sinensis (L.) Osbeck) juices attenuate left ventricular dysfunction and oxidative stress and improve myocardial energy metabolism in acute doxorubicin-induced cardiotoxicity in rats. Nutrition 2021, 91–92, 111350. [Google Scholar] [CrossRef]
- Fraga, L.N.; Milenkovic, D.; Lajolo, F.M.; Hassimotto, N.M.A. Association between Single Nucleotide Polymorphisms of SULT1A1, SULT1C4, ABCC2 and Phase II Flavanone Metabolites Excretion after Orange Juice Intake. Nutrients 2022, 14, 3770. [Google Scholar] [CrossRef] [PubMed]
- Dorna, M.S.; Barbosa, E.M.S.; Callegari, M.A.; Tanni, S.E.; Chiuso-Minicucci, F.; Felix, T.F.; Seneda, A.L.; Correa, C.R.; Fernandes, A.A.H.; Azevedo, P.S.; et al. Orange Juice Attenuates Circulating miR-150-5p, miR-25-3p, and miR-451a in Healthy Smokers: A Randomized Crossover Study. Front. Nutr. 2021, 8, 775515. [Google Scholar] [CrossRef]
- Quintanilha, B.J.; Chaves, D.F.S.; Brasili, E.; Corrêa, T.A.F.; Capetini, V.C.; Ferreira, F.M.; Castro, I.A.; Hassimotto, N.M.A.; Rogero, M.M.; Lajolo, F.M. Ingestion of orange juice prevents hyperglycemia and increases plasma miR-375 expression. Clin. Nutr. ESPEN 2022, 47, 240e24. [Google Scholar] [CrossRef]
- Capetini, V.C.; Quintanilha, B.J.; Oliveira, D.C.; Nishioka, A.H.; Matos, L.A.; Ferreira, L.R.P.; Ferreira, F.M.; Sampaio, G.R.; Hassimotto, N.M.A.; Lajolo, F.M.; et al. Blood orange juice intake modulates plasma and PBMC microRNA expression in overweight and insulin resistance women: Impact on MAPK and NFB signaling pathways. J. Nutr. Biochem. 2022, 109240. [Google Scholar]
- Stanislawski, M.A.; Dabelea, D.; Lange, L.A.; Wagner, B.D.; Lozupone, C.A. Gut microbiota phenotypes of obesity. NPJ Biofilms Microbiomes 2019, 5, 18. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef]
- Andoh, A.; Nishida, A.; Takahashi, K.; Inatomi, O.; Imaeda, H.; Bamba, S.; Kito, K.; Sugimoto, M.; Kobayashi, T. Comparison of the gut microbial community between obese and lean peoples using 16S gene sequencing in a Japanese population. J. Clin. Biochem. Nutr. 2016, 59, 65–70. [Google Scholar] [CrossRef] [Green Version]
- Jinatham, V.; Kullawong, N.; Kespechara, K.; Gentekaki, E.; Popluechai, S. Comparison of Gut Microbiota between Lean and Obese Adult Thai Individuals. Microbiol. Biotechnol. Lett. 2018, 46, 277–287. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Hamady, M.; Yatsunenko, T.; Cantarel, B.L.; Duncan, A.; Ley, R.E.; Sogin, M.L.; Jones, W.J.; Roe, B.A.; Affourtit, J.P.; et al. A core gut microbiome in obese and lean twins. Nature 2009, 457, 480–484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rogers, A.W.L.; Tsolis, R.M.; Bäumler, A.J. Salmonella versus the Microbiome. Microbiol. Mol. Biol. Rev. 2020, 85, e00027-19. [Google Scholar] [CrossRef]
- Lee, J.; Tsolis, R.M.; Bäumler, A.J. The microbiome and gut homeostasis. Science 2022, 377, 6601. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.F.; Zhu, M.Q.; Zhang, H.; Liu, H.; Xia, B.; Wang, Y.L.; Shi, X.; Peng, L.; Wu, J.W. Neohesperidin attenuates obesity by altering the composition of the gut microbiota in high-fat diet-fed mice. FASEB J. 2020, 34, 12053–12071. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Knight, R.; Gordon, J.I. The effect of diet on the human gut microbiome: A metagenomic analysis in humanized gnotobiotic mice. Sci. Transl. Med. 2009, 1, 6ra14. [Google Scholar] [CrossRef] [Green Version]
- Azad, M.B.; Bridgman, S.L.; Becker, A.B.; Kozyrskyj, A.L. Infant antibiotic exposure and the development of childhood overweight and central adiposity. Int. J. Obes. 2014, 38, 1290–1298. [Google Scholar] [CrossRef]
- Azad, M.B.; Konya, T.; Persaud, R.R.; Guttman, D.S.; Chari, R.S.; Field, C.J.; Sears, M.R.; Mandhane, P.J.; Turvey, S.E.; Subbarao, P. Impact of maternal intrapartum antibiotics, method of birth and breastfeeding on gut microbiota during the first year of life: A prospective cohort study. BJOG 2016, 123, 983–993. [Google Scholar] [CrossRef]
- Ottosson, F.F.; Brunkwall, L.; Ericson, U.; Nilsson, P.M.; Almgren, P.; Fernandez, C.; Melander, O.; Orho-Melander, M. Connection Between BMI-Related Plasma Metabolite Profile and Gut Microbiota. J. Clin. Endocrinol. Metab. 2018, 103, 1491–1501. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Moreno, M.; Perez-Herrera, A.; Locia-Morales, D.; Dizzel, S.; Meyre, D.; Stearns, J.C.; Cruz, M. Association of gut microbiome with fasting triglycerides, fasting insulin and obesity status in Mexican children. Pediatr. Obes. 2021, 16, e12748. [Google Scholar] [CrossRef]
- Companys, J.; Gosalbes, M.J.; Pla-Pagà, L.; Calderón-Pérez, L.; Llauradó, E.; Pedret, A.; Valls, R.M.; Jiménez-Hernández, N.; Sandoval-Ramirez, B.A.; Del Bas, J.M.; et al. Gut Microbiota Profile and Its Association with Clinical Variables and Dietary Intake in Overweight/Obese and Lean Subjects: A Cross-Sectional Study. Nutrients 2021, 13, 2032. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Li, D.; He, Y.; Li, Y.; Yang, Z.; Zhao, X.; Liu, Y.; Wang, Y.; Sun, J.; Feng, X.; et al. Discrepant gut microbiota markers for the classification of obesity-related metabolic abnormalities. Sci. Rep. 2019, 9, 13424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, K.; Yu, W.; Li, S.; Guan, X.; Liu, J.; Song, H.; Liu, D.; Duan, R. Effect of embryo-remaining oat rice on the lipid profile and intestinal microbiota in high-fat diet fed rats. Food Res. Int. 2020, 129, 108816. [Google Scholar] [CrossRef] [PubMed]
- Braune, A.; Blaut, M. Bacterial species involved in the conversion of dietary flavonoids in the human gut. Gut Microbes 2016, 7, 216–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Zhao, S.; Wang, J.; Shi, J.; Sun, Y.; Wang, W.; Ning, G.; Hong, J.; Liu, R. Grape seed proanthocyanidin extract ameliorates inflammation and adiposity by modulating gut microbiota in high-fat diet mice. Mol. Nutr. Food Res. 2017, 61, 1601082. [Google Scholar] [CrossRef]
- Mayta-Apaza, A.C.; Pottgen, E.; Bodt, J.D.; Papp, N.; Marasini, D.; Howard, L.; Abranko, L.; Wiele, T.V.; Lee, S.O.; Carbonero, F. Impact of tart cherries polyphenols on the human gut microbiota and phenolic metabolites in vitro and in vivo. J. Nutr. Biochem. 2018, 59, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Petersen, C.; Wankhade, U.D.; Bharat, D.; Wong, K.; Mueller, J.E.; Chintapalli, S.V.; Piccolo, B.D.; Jalili, T.; Jia, Z.; Symons, J.D.; et al. Dietary supplementation with strawberry induces marked changes in the composition and functional potential of the gut microbiome in diabetic mice. J. Nutr. Biochem. 2019, 66, 63–69. [Google Scholar] [CrossRef]
- Mas-Capdevila, A.; Teichenne, J.; Domenech-Coca, C.; Caimari, A.; Del Bas, J.M.; Escoté, X.; Crescenti, A. Effect of Hesperidin on Cardiovascular Disease Risk Factors: The Role of Intestinal Microbiota on Hesperidin Bioavailability. Nutrients 2020, 12, 1488. [Google Scholar] [CrossRef]
- Dueñas, M.; Muñoz-González, I.; Cueva, C.; Jiménez-Girón, A.; Sánchez-Patán, F.; Santos-Buelga, C.; Moreno-Arribas, M.V.; Bartolomé, B. A Survey of Modulation of Gut Microbiota by Dietary Polyphenols. Biomed Res. Int. 2015, 850902, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Chang, H.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of diet on the gut microbiome and implications for human health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [Green Version]
- Gentile, C.L.; Weir, T.L. The gut microbiota at the intersection of diet and human health. Science 2018, 362, 776–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Association of Official Analytical Chemists (AOAC). Official Methods of Analysis, 16th ed.; AOAC International: Arlington, TX, USA, 1995. [Google Scholar]
- Brasili, E.; Chaves, D.F.; Xavier, A.A.; Mercadante, A.Z.; Hassimotto, N.M.A.; Lajolo, F.M. Effect of Pasteurization on Flavonoids and Carotenoids in Citrus sinensis (L.) Osbeck cv. ‘Cara Cara’ and ‘Bahia’ Juices. J. Agric. Food Chem. 2017, 65, 1371–1377. [Google Scholar] [CrossRef] [PubMed]
- Hillebrand, S.; Schwarz, M.; Winterhalter, P. Characterization of Anthocyanins and Pyranoanthocyanins from Blood Orange [Citrus sinensis (L.) Osbeck] Juice. J. Agric. Food Chem. 2004, 52, 7331–7338. [Google Scholar] [CrossRef] [PubMed]
- Shiga, T.; Soares, C.A.; Nascimento, J.R.O.; Purgatto, E.; Lajolo, F.M.; Cordenunsi, B.R. Ripening-associated changes in the amounts of starch and non-starch polysaccharides and their contributions to fruit softening in three banana cultivars. J. Sci. Food Agric. 2011, 91, 1511–1516. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Stella, S.P.; Ferrarezi, A.C.; Santos, K.O.; Monteiro, M. Antioxidant activity of commercial ready-to-drink orange juice and nectar. J. Food Sci. 2011, 76, 392–397. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Baseline Parameters | Obesity | Reference Values | ||||
|---|---|---|---|---|---|---|
| Class I (n = 12) | Class II (n = 5) | Class III (n = 6) | Ideal | Prediabetes | ||
| Body weight (kg) | 94.52 ± 2.72 | 87.48 ± 3.38 | 99.78 ± 2.59 | 104.22 ± 5.15 | - | - |
| BMI (kg·m−2) | 36.69 ± 1.26 | 32.36 ± 0.38 | 37.29 ± 0.80 | 44.85 ± 2.19 | - | - |
| Fasting glucose (mg·dL−1) | 100.96 ± 2.35 | 103.08 ± 3.66 | 98.00 ± 3.49 | 99.17 ± 4.71 | <99 * | 100 to 125 * |
| Fasting insulin (µIU·mL−1) | 16.43 ± 0.82 | 15.55 ± 1.12 | 15.82 ± 1.64 | 18.68 ± 1.62 | 1.9 to 23 * | - |
| HbA1c (%) | 5.96 ± 0.07 | 5.98 ± 0.06 | 5.94 ± 0.28 | 5.93 ± 0.15 | <5.7 * | 5.7 to 6.4 * |
| HOMA-IR | 4.05 ± 0.19 | 3.93 ± 0.27 | 3.79 ± 0.35 | 4.52 ± 0.37 | ≤2.71 [26] | >2.71 [26] |
| POJ | MOJ | |||||
|---|---|---|---|---|---|---|
| T0 | T16 | p-Value | T0 | T16 | p-Value | |
| Body weight (kg) | 94.48 ± 2.69 | 94.43 ± 2.71 | 0.603 | 93.74 ± 2.65 | 93.61 ± 2.70 | 0.503 |
| Abdominal circumference (cm) | 115.36 ± 24.07 | 115.22 ± 23.97 | 0.594 | 114.46 ± 23.84 | 113.37 ± 23.66 | 0.887 |
| BMI (kg·m−2) | 36.66 ± 7.65 | 36.63 ± 7.60 | 0.661 | 36.37 ± 7.58 | 36.30 ± 7.66 | 0.456 |
| Class I obesity (n = 12) | 32.48 ± 0.40 | 32.54 ± 0.36 | 0.367 | 32.28 ± 0.33 | 32.21 ± 0.34 | 0.638 |
| Class II obesity (n = 5) | 37.35 ± 0.83 | 36.95 ± 0.98 | 0.273 | 36.51 ± 1.11 | 36.61 ± 1.26 | 0.686 |
| Class III obesity (n = 6) | 44.46 ± 2.33 | 44.54 ± 2.18 | 0.600 | 44.41 ± 1.84 | 44.21 ± 1.73 | 0.249 |
| Blood pressure (mmHg) | ||||||
| Systolic | 132.43 ± 27.64 | 125.78 ± 26.31 | 0.083 | 129.17 ± 27.03 | 126.30 ± 26.38 | 0.403 |
| Diastolic | 82.30 ± 17.14 | 79.74 ± 16.74 | 0.144 | 81.35 ± 16.82 | 79.43 ± 16.76 | 0.700 |
| Cholesterol (mg·dL−1) | ||||||
| HDL | 45.91 ± 9.43 a | 43.26 ± 9.15 b | 0.045 | 47.78 ± 10.02 a | 45.70 ± 9.67 b | 0.042 |
| LDL | 107.87 ± 22.45 | 105.35 ± 23.12 | 0.260 | 115.83 ± 23.29 | 110.00 ± 23.69 | 0.136 |
| VLDL | 30.09 ± 6.19 | 28.52 ± 6.10 | 0.501 | 32.57 ± 6.52 | 30.17 ± 6.40 | 0.253 |
| Total | 184.17 ± 38.13 | 177.39 ± 38.42 | 0.119 | 197.61 ± 40.12 a | 184.57 ± 39.49 b | 0.012 |
| Triglycerides (mg·dL−1) | 151.26 ± 31.11 | 142.39 ± 30.44 | 0.426 | 174.04 ± 34.97 | 151.00 ± 32.13 | 0.457 |
| Blood glucose (mg·dL−1) | 94.61 ± 19.45 | 94.52 ± 19.96 | 0.896 | 96.83 ± 20.21 | 97.13 ± 20.19 | 0.807 |
| POJ | MOJ | |||||
|---|---|---|---|---|---|---|
| T0 | T16 | p-Value | T0 | T16 | p-Value | |
| Inflammatory biomarkers | ||||||
| IL-6 (pg·mL−1) | 5.28 ± 1.10 | 4.88 ± 1.18 | 0.349 | 5.76 ± 1.21 | 3.79 ± 0.93 | 0.179 |
| IL-10 (pg·mL−1) | 1.02 ± 0.22 | 1.28 ± 0.30 | 0.349 | 1.26 ± 0.28 | 1.28 ± 0.29 | 0.501 |
| TNF-α (pg·mL−1) | 95.50 ± 26.57 | 163.86 ± 43.11 | 0.068 | 148.83 ± 33.21 | 146.22 ± 36.89 | 0.619 |
| IL-8 (pg·mL−1) | 652.26 ± 146.08 | 650.84 ± 155.01 | 0.747 | 733.76 ± 157.15a | 604.62 ± 137.97b | 0.025 |
| IL-1β (pg·mL−1) | 253.60 ± 63.60 | 235.53 ± 64.60 | 0.433 | 251.35 ± 53.63 | 209.10 ± 53.95 | 0.074 |
| MCP-1 (pg·mL−1) | 5571.72 ± 1336.21 | 3603.72 ± 1286.96 | 0.231 | 10787.93 ± 3501.45 | 3746.00 ± 766.01 | 0.391 |
| Oxidative stress biomarkers | ||||||
| Plasma | ||||||
| ORAC (µM eq Trolox·mL−1) | 2.45 ± 0.54b | 2.77 ± 0.55a | 0.006 | 3.02 ± 0.63 | 3.03 ± 0.62 | 0.910 |
| GPx (U·mg−1) | 0.93 ± 0.17 | 0.74 ± 0.20 | 0.263 | 1.10 ± 0.20a | 0.75 ± 0.17b | 0.023 |
| SOD (U·mg−1) | 2.48 ± 0.61 | 3.10 ± 0.70 | 0.276 | 3.68 ± 0.72 | 3.39 ± 0.70 | 0.360 |
| Urine | ||||||
| 8-isoprostane (pg·mg−1) | 2.59 ± 0.58 | 2.02 ± 0.48 | 0.848 | 2.59 ± 0.55 | 2.46 ± 0.49 | 0.831 |
| 8-OHdG (ng·mg−1) | 1.63 ± 0.32a | 0.47 ± 0.12b | <0.001 | 1.21 ± 0.23a | 0.55 ± 0.14b | <0.001 |
| Global | Class I Obesity | Class II Obesity | Class III Obesity | p-Value | |
|---|---|---|---|---|---|
| Firmicutes | 87.43 ± 1.51 | 86.09 ± 1.78 | 91.89 ± 2.67 | 86.32 ± 4.29 | 0.233 |
| Bacteroidetes | 4.51 ± 1.01 | 5.58 ± 1.42 | 4.49 ± 2.67 | 1.86 ± 0.60 | 0.203 |
| F/B | 50.28 ± 13.52 | 39.16 ± 19.38 | 55.49 ± 25.12 | 72.86 ± 30.56 | 0.203 |
| Actinobacteria | 6.51 ± 1.37 | 6.16 ± 1.46 | 2.56 ± 0.83 | 11.34 ± 4.26 | 0.253 |
| Proteobacteria | 0.45 ± 0.08 | 0.54 ± 0.13 | 0.38 ± 0.17 | 0.29 ± 0.10 | 0.557 |
| Verrucomicrobia | 0.63 ± 0.23 | 0.64 ± 0.22 | 0.80 ± 0.76 | 0.08 ± 0.00 | 0.307 |
| Tenericutes | 0.37 ± 0.13 | 0.40 ± 0.15 | N.D. | 0.21 ± 0.09 | - |
| Others | 0.47 ± 0.30 | 0.78 ± 0.54 | 0.09 ± 0.05 | 0.07 ± 0.05 | 0.133 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
de Santana, A.A.; de Castro Tobaruela, E.; dos Santos, K.G.; Sparvoli, L.G.; do Amaral, C.K.; Magnoni, C.D.; Taddei, C.R.; dos Santos, R.V.T.; Hassimotto, N.M.A.; Lajolo, F.M. ‘Pera’ Orange and ‘Moro’ Blood Orange Juice Improves Oxidative Stress and Inflammatory Response Biomarkers and Modulates the Gut Microbiota of Individuals with Insulin Resistance and Different Obesity Classes. Obesities 2022, 2, 389-412. https://doi.org/10.3390/obesities2040033
de Santana AA, de Castro Tobaruela E, dos Santos KG, Sparvoli LG, do Amaral CK, Magnoni CD, Taddei CR, dos Santos RVT, Hassimotto NMA, Lajolo FM. ‘Pera’ Orange and ‘Moro’ Blood Orange Juice Improves Oxidative Stress and Inflammatory Response Biomarkers and Modulates the Gut Microbiota of Individuals with Insulin Resistance and Different Obesity Classes. Obesities. 2022; 2(4):389-412. https://doi.org/10.3390/obesities2040033
Chicago/Turabian Stylede Santana, Aline Alves, Eric de Castro Tobaruela, Karina Gama dos Santos, Luiz Gustavo Sparvoli, Cristiane Kovacs do Amaral, Carlos Daniel Magnoni, Carla R. Taddei, Ronaldo Vagner Thomatieli dos Santos, Neuza Mariko Aymoto Hassimotto, and Franco Maria Lajolo. 2022. "‘Pera’ Orange and ‘Moro’ Blood Orange Juice Improves Oxidative Stress and Inflammatory Response Biomarkers and Modulates the Gut Microbiota of Individuals with Insulin Resistance and Different Obesity Classes" Obesities 2, no. 4: 389-412. https://doi.org/10.3390/obesities2040033