
How Do Nitrogen Deposition, Mowing, and Deer Grazing Drive Vegetation Changes on Dune Heaths?

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description
2.2. Experimental Design
2.3. Sampling and Data Handling
2.4. Models
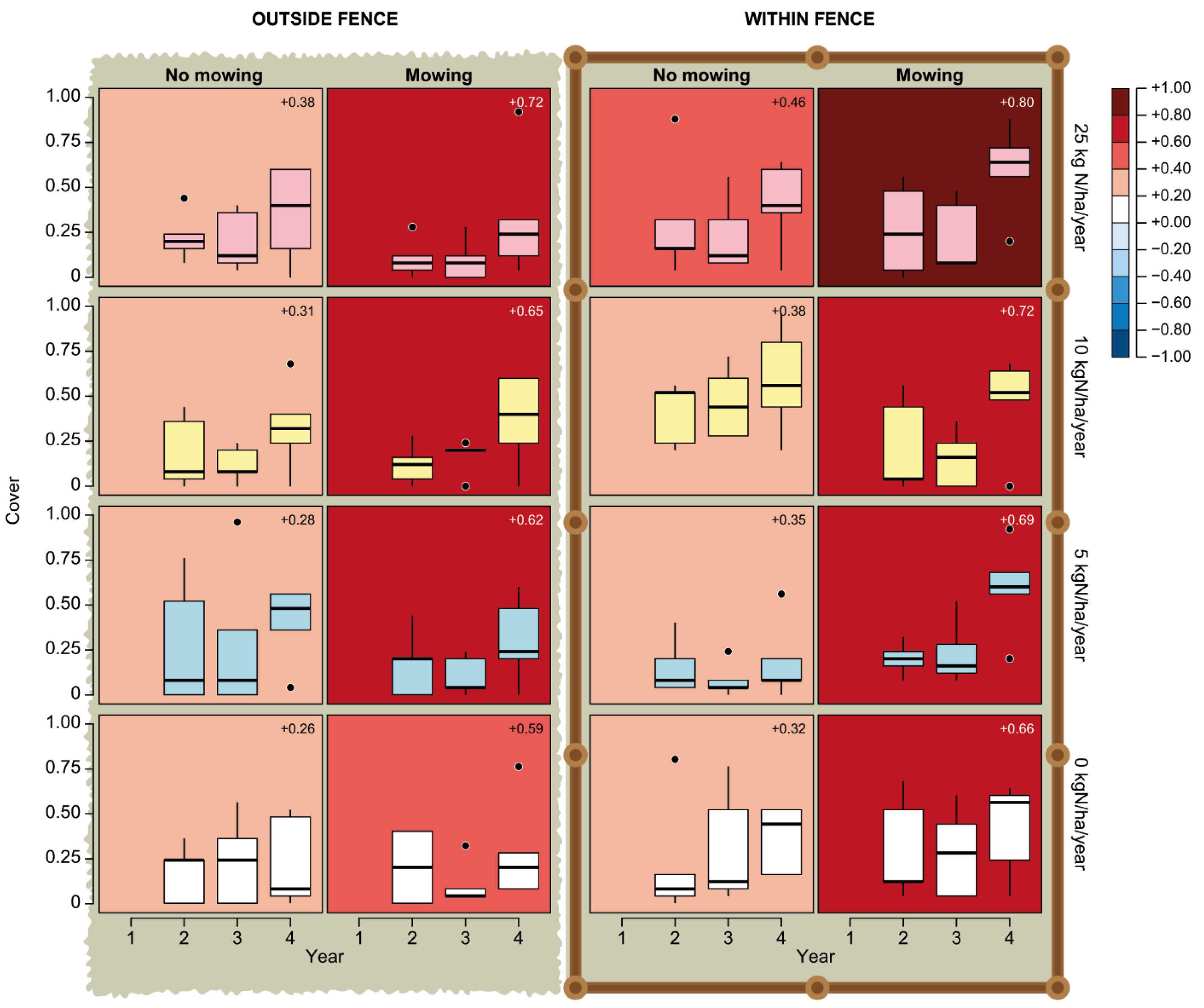
3. Results
4. Discussion
- Bryophytes seem to be affected adversely even at deposition levels at or below the empirical critical load of dune heaths, which is 10 to 15 kg N ha−1 year−1. The decline of bryophytes seems to happen even in plots with annual mowing where the material is removed and with a low dose of added nitrogen (i.e., 5 kg N ha−1 year−1). Therefore, keeping nitrogen deposition levels low is pivotal to protect these sensitive elements.
- It is essential to mention that our mowing method positively affects bryophytes and the moss species P. schreberi. However, we advise managers to avoid annual mowing near the soil surface because it promotes graminoids and causes a decline in E. nigrum, but not to avoid mowing in general.
- Excluding deer enhances growth and increases coverage of C. arenaria at higher N deposition. Therefore, allowing deer numbers to increase might be beneficial for dune heaths.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- European Commission. Interpretation Manual of European Union Habitats; European Environment Agency: Copenhagen, Denmark, 2013; p. 144. [Google Scholar]
- Nygaard, B.; Darngaard, C.; Nielsen, K.E.; Bladt, J.; Ejrnæs, R. Terrestriske Naturtyper 2004–2015 Novana; Aarhus Universitet DCE-Nationalt Center for Miljø og Energi: Aarhus, Denmark, 2016; p. 1201. [Google Scholar]
- Nygaard, B.; Ejrnæs, R.; Fredshavn, J.R. Kortlægning Af Habitatnaturtyper 2019. NOVANA; Aarhus Universitet DCE-Nationalt Center for Miljø og Energi: Aarhus, Denmark, 2021; p. 462. [Google Scholar]
- Aerts, R.; Heil, G.W. Heathlands: Patterns and Processes in a Changing Environment; Kluwer Academic Publishers: London, UK, 1993; p. 223. [Google Scholar]
- De Graaf, M.C.C.; Bobbink, R.; Smits, N.A.C.; Van Diggelen, R.; Roelofs, J.G.M. Biodiversity, vegetation gradients and key biogeochemical processes in the heathland landscape. Biol. Conserv. 2009, 142, 2191–2201. [Google Scholar] [CrossRef]
- Kleijn, D.; Bekker, R.M.; Bobbink, R.; De Graaf, M.C.C.; Roelofs, J.G.M. In search for key biogeochemical factors affecting plant species persistence in heathland and acidic grasslands: A comparison of common and rare species. J. Appl. Ecol. 2008, 45, 680–687. [Google Scholar] [CrossRef]
- ETC/BD; EEA. Article 17 Web Tool-Habitat Assessments at EU Biogeographical Level. Available online: https://nature-art17.eionet.europa.eu/article17/habitat/summary/?period=5&group=Dunes+habitats&subject=2140®ion= (accessed on 15 February 2024).
- Aerts, R.; Bobbink, R. The Impact of Atmospheric Nitrogen Deposition on Vegetation Processes in Terrestrial, Non-Forest Ecosystems; Springer: Dordrecht, The Netherlands, 1999; pp. 85–122. [Google Scholar]
- Bähring, A.; Fichtner, A.; Ibe, K.; Schütze, G.; Temperton, V.M.; von Oheimb, G.; Härdtle, W. Ecosystem functions as indicators for heathland responses to nitrogen fertilisation. Ecol. Indic. 2017, 72, 185–193. [Google Scholar] [CrossRef]
- Heil, G.W.; Bobbink, R. “Calluna”, a simulation model for evaluation of impacts of atmospheric nitrogen deposition on dry heathlands. Ecol. Model. 1993, 68, 161–182. [Google Scholar] [CrossRef]
- Härdtle, W.; Niemeyer, M.; Niemeyer, T.; Assmann, T.; Fottner, S. Can management compensate for atmospheric nutrient deposition in heathland ecosystems? J. Appl. Ecol. 2006, 43, 759–769. [Google Scholar] [CrossRef]
- Barker, C.G.; Power, S.A.; Bell, J.N.B.; Orme, C.D.L. Effects of habitat management on heathland response to atmospheric nitrogen deposition. Biol. Conserv. 2004, 120, 41–52. [Google Scholar] [CrossRef]
- Helliwell, R.C.; Britton, A.J.; Gibbs, S.; Fisher, J.M.; Potts, J.M. Interactive effects of N deposition, land management and weather patterns on soil solution chemistry in a Scottish alpine heath. Ecosystems 2010, 13, 696–711. [Google Scholar] [CrossRef]
- Riesch, F.; Tonn, B.; Meißner, M.; Balkenhol, N.; Isselstein, J. Grazing by wild red deer: Management options for the conservation of semi-natural open habitats. J. Appl. Ecol. 2019, 56, 1311–1321. [Google Scholar] [CrossRef]
- Riesch, F.; Wichelhaus, A.; Tonn, B.; Meißner, M.; Rosenthal, G.; Isselstein, J. Grazing by wild red deer can mitigate nutrient enrichment in protected semi-natural open habitats. Oecologia 2022, 199, 471–485. [Google Scholar] [CrossRef]
- Heil, G.W.; Aerts, R. General introduction. In Heathlands: Patterns and Processes in a Changing Environment; Springer: Dordrecht, The Netherlands, 1993; pp. 1–24. [Google Scholar]
- Bobbink, R.; Loran, C.; Tomassen, H. Review and Revision of Empirical Critical Loads of Nitrogen for Europe; Bobbink, R., Loran, C., Tomassen, H., Eds.; German Environment Agency: Dessau-Roßlau, Germany, 2022; p. 358. [Google Scholar]
- Engardt, M.; Simpson, D.; Schwikowski, M.; Granat, L. Deposition of sulphur and nitrogen in Europe 1900–2050. Model calculations and comparison to historical observations. Tellus Ser. B Chem. Phys. Meteorol. 2017, 69, 1–20. [Google Scholar] [CrossRef]
- McClean, C.J.; Van Den Berg, L.J.L.; Ashmore, M.R.; Preston, C.D. Atmospheric nitrogen deposition explains patterns of plant species loss. Glob. Change Biol. 2011, 17, 2882–2892. [Google Scholar] [CrossRef]
- Heil, G.W.; Bobbink, R. Impact of Atmospheric Nitrogen Deposition on Dry Heathlands BT—Heathlands: Patterns and Processes in a Changing Environment; Aerts, R., Heil, G.W., Eds.; Springer: Dordrecht, The Netherlands, 1993; pp. 181–200. [Google Scholar]
- Phoenix, G.K.; Emmett, B.A.; Britton, A.J.; Caporn, S.J.M.; Dise, N.B.; Helliwell, R.; Jones, L.; Leake, J.R.; Leith, I.D.; Sheppard, L.J.; et al. Impacts of atmospheric nitrogen deposition: Responses of multiple plant and soil parameters across contrasting ecosystems in long-term field experiments. Glob. Change Biol. 2012, 18, 1197–1215. [Google Scholar] [CrossRef]
- Fagúndez, J. Heathlands confronting global change: Drivers of biodiversity loss from past to future scenarios. Ann. Bot. 2013, 111, 151–172. [Google Scholar] [CrossRef]
- De Schrijver, A.; De Frenne, P.; Ampoorter, E.; Van Nevel, L.; Demey, A.; Wuyts, K.; Verheyen, K. Cumulative nitrogen input drives species loss in terrestrial ecosystems. Glob. Ecol. Biogeogr. 2011, 20, 803–816. [Google Scholar] [CrossRef]
- Power, S.A.; Ashmore, M.R.; Cousins, D.A. Impacts and fate of experimentally enhanced nitrogen deposition on a British lowland heath. Environ. Pollut. 1998, 102, 27–34. [Google Scholar] [CrossRef]
- Bak, J. Kortlægnings og Analyseprojekt Vedrørende Væsentlige Ammoniakpunktkilder og Sårbare Naturtyper i Det Åbne Land; Skov-og Naturstyrelsen, Wilhjelmudvalget: Copenhagen, Denmark, 2001; pp. 1–27. [Google Scholar]
- Gordon, C.; Wynn, J.M.; Woodin, S.J. Impacts of increased nitrogen supply on high Arctic heath: The importance of bryophytes and phosphorus availability. New Phytol. 2001, 149, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Southon, G.E.; Field, C.; Caporn, S.J.M.; Britton, A.J.; Power, S.A. Nitrogen Deposition Reduces Plant Diversity and Alters Ecosystem Functioning: Field-Scale Evidence from a Nationwide Survey of UK Heathlands. PLoS ONE 2013, 8, 59031. [Google Scholar] [CrossRef] [PubMed]
- Aerts, R.; Berendse, F. The effect of increased nutrient availability on vegetation dynamics in wet heathlands. Vegetatio 1988, 76, 63–69. [Google Scholar] [CrossRef]
- Morecroft, M.D.; Sellers, E.K.; Lee, J.A. An Experimental Investigation into the Effects of Atmospheric Nitrogen Deposition on Two Semi-Natural Grasslands. J. Ecol. 1994, 82, 475–483. [Google Scholar] [CrossRef]
- Power, S.A.; Ashmore, M.R.; Cousins, D.A.; Sheppard, L.J. Effects of nitrogen addition on the stress sensitivity of Calluna vulgaris. New Phytol. 1998, 138, 663–673. [Google Scholar] [CrossRef]
- Fremstad, E. Virkninger av Nitrogen på Heivegetasjon. En Litteraturstudie; NINA Oppdragsmelding: Trondheim, Norway, 1992; p. 44. [Google Scholar]
- Cunha, A.; Power, S.; Ashmore, M.; Green, P.; Haworth, B.; Bobbink, R. Whole ecosystem nitrogen manipulation: An updated review. Rep. Jt. Nat. Conserv. Comm. 2002, 44, 126. [Google Scholar]
- Aerts, R. Aboveground Biomass and Nutrient Dynamics of Calluna vulgaris and Molinia caerulea in a Dry Heathland. Oikos 1989, 56, 31–38. [Google Scholar] [CrossRef]
- Schellenberg, J.; Bergmeier, E. The Calluna life cycle concept revisited: Implications for heathland management. Biodivers. Conserv. 2022, 31, 119–141. [Google Scholar] [CrossRef]
- Remke, E.; Brouwer, E.; Kooijman, A.; Blindow, I.; Roelofs, J.G.M. Low atmospheric nitrogen loads lead to grass encroachment in Coastal Dunes, but only on acid soils. Ecosystems 2009, 12, 1173–1188. [Google Scholar] [CrossRef]
- Jones, L.; Stevens, C.; Rowe, E.C.; Payne, R.; Caporn, S.J.M.; Evans, C.D.; Field, C.; Dale, S. Can on-site management mitigate nitrogen deposition impacts in non-wooded habitats? Biol. Conserv. 2017, 212, 464–475. [Google Scholar] [CrossRef]
- Schellenberg, J.; Bergmeier, E. Heathland plant species composition and vegetation structures reflect soil-related paths of development and site history. Appl. Veg. Sci. 2020, 23, 386–405. [Google Scholar] [CrossRef]
- Schmidt, I.K.; Gundersen, P. Kvælstoffjernelse ved Naturpleje—Vidensgrundlag og Opfølgende Forskning; Københavns Universitet. IGN Rapport: København, Denmark, 2018; p. 39. [Google Scholar]
- Byriel, D.B.; Ro-Poulsen, H.; Kepfer-Rojas, S.; Hansen, A.K.; Hansen, R.R.; Justesen, M.J.; Kristensen, E.; Møller, C.B.; Schmidt, I.K. Contrasting responses of multiple insect taxa to common heathland management regimes and old-growth successional stages. Biodivers. Conserv. 2022, 32, 545–565. [Google Scholar] [CrossRef]
- Hansen, R.R.; Nielsen, K.E.; Offenberg, J.; Damgaard, C.; Byriel, D.B.; Schmidt, I.K.; Sørensen, P.B.; Kjær, C.; Strandberg, M.T. Implications of heathland management for ant species composition and diversity—Is heathland management causing biotic homogenization? Biol. Conserv. 2020, 242, 108422. [Google Scholar] [CrossRef]
- Maes, D.; Decleer, K.; De Keersmaeker, L.; Van Uytvanck, J.; Louette, G. Intensified habitat management to mitigate negative effects of nitrogen pollution can be detrimental for faunal diversity: A comment on Jones et al. (2017). Biol. Conserv. 2017, 212, 493–494. [Google Scholar] [CrossRef]
- Britton, A.J.; Marrs, R.H.; Carey, P.D.; Pakeman, R.J. Comparison of techniques to increase Calluna vulgaris cover on heathland invaded by grasses in Breckland, south east England. Biol. Conserv. 2000, 95, 227–232. [Google Scholar] [CrossRef]
- Milligan, A.L.; Putwain, P.D.; Cox, E.S.; Ghorbani, J.; Le Duc, M.G.; Marrs, R.H. Developing an integrated land management strategy for the restoration of moorland vegetation on Molinia caerulea-dominated vegetation for conservation purposes in upland Britain. Biol. Conserv. 2004, 119, 371–385. [Google Scholar] [CrossRef]
- Webb, N.R. The traditional management of European heathlands. J. Appl. Ecol. 1998, 35, 987–990. [Google Scholar] [CrossRef]
- Hofmann, R.R. Evolutionary steps of ecophysiological adaptation and diversification of ruminants: A comparative view of their digestive system. Oecologia 1989, 78, 443–457. [Google Scholar] [CrossRef] [PubMed]
- Sunde, P.; Haugaard, L. Bæredygtig Krondyrforvaltning. Populationsbiologiske Analyser af Krondyrbestandene på Oksbøl og Djursland Med Reference til Jagtlig Forvaltning; Aarhus Universitet DCE—Nationalt Center for Miljø og Energi: Aarhus, Denmark, 2014; p. 76. [Google Scholar]
- Bülow-Olsen, A. Changes in the species composition in an area dominated by Deschamsia flexuosa (L.) trin. as a result of cattle grazing. Biol. Conserv. 1980, 18, 257–270. [Google Scholar] [CrossRef]
- Hartley, S.E.; Mitchell, R.J. Manipulation of nutrients and grazing levels on heather moorland: Changes in Calluna dominance and consequences for community composition. J. Ecol. 2005, 93, 990–1004. [Google Scholar] [CrossRef]
- Riesch, F.; Tonn, B.; Stroh, H.G.; Meißner, M.; Balkenhol, N.; Isselstein, J. Grazing by wild red deer maintains characteristic vegetation of semi-natural open habitats: Evidence from a three-year exclusion experiment. Appl. Veg. Sci. 2020, 23, 522–538. [Google Scholar] [CrossRef]
- Smith, S.W.; Johnson, D.; Quin, S.L.O.; Munro, K.; Pakeman, R.J.; van der Wal, R.; Woodin, S.J. Combination of herbivore removal and nitrogen deposition increases upland carbon storage. Glob. Chang. Biol. 2015, 21, 3036–3048. [Google Scholar] [CrossRef]
- Hope, D.; Picozzi, N.; Catt, D.C.; Moss, R. Effects of reducing sheep grazing in the Scottish highlands. J. Range Manag. 1996, 49, 301–310. [Google Scholar] [CrossRef]
- Danmarks Arealinformation. Available online: https://arealinformation.miljoeportal.dk/html5/index.html?viewer=distribution (accessed on 17 February 2024).
- Olsen, O.N. Topographisk Kaartsignaturer; Den Kongelige Danske Generalqvarteermesterstab: København, Denmark, 1831. [Google Scholar]
- Dmi. Danmarks Meterologiske Institut. Available online: https://www.dmi.dk/ (accessed on 26 April 2020).
- Ellerman, T.; Bossi, R.; Nygaard, J.; Christensen, J.; Løfstrøm, P.; Monies, C.; Geels, C.; Nilesen, I.E.; Poulsen, M.B. Atmosfærisk Deposition 2019 Videnskabelig Rapport fra DCE-Nationalt Center for Miljø og Energi; Aarhus Universitet, DCE—Nationalt Center for Miljø og Energi ©: Aarhus, Denmark, 2021; p. 90. [Google Scholar]
- Adhikari, K.; Minasny, B.; Greve, M.B.; Greve, M.H. Constructing a soil class map of Denmark based on the FAO legend using digital techniques. Geoderma 2014, 214–215, 101–113. [Google Scholar] [CrossRef]
- KB. Danmark Set Fra Luften—Det Kgl. Bibliotek. Available online: https://www.kb.dk/danmarksetfraluften/?q_fritekst=&q_stednavn=&q_bygningsnavn=&q_person=¬Before=1890¬After=2015&category=subject243&itemType=all&thumbnailSize=&correctness=-1&thumbnailSize=&sortby=filename&sortorder=asc#zoom=17&lat=57.12397339005477&lng=9.011675119400026 (accessed on 5 November 2022).
- Miljøstyrelsen DCE & GEUS. NOVANA Det Nationale Overvågningsprogram for Vandmiljø og Natur 2017–21 Programbeskrivelse; Miljøstyrelsen: Aarhus, Denmark, 2017; p. 183. [Google Scholar]
- Fredshavn, J.; Søgaard, B.; Nygaard, B.; Johansson Sander, J.; Wiberg-Larsen, P.; Dahl, K.; Sveegaard, S.; Galatius, A.; Teilman, J. Bevaringsstatus for Naturtyper og arter. Habitatdirektivets Artikel 17 Rapportering; Aarhus Universitet DCE—Nationalt Center for Miljø og Energi, Videnskabelig rapport nr. 98: Aarhus, Denmark, 2014; p. 54. [Google Scholar]
- Harley, J.L.; Harley, E.L. A Check-List of Mycorrhiza in the British Flora. Source New Phytol. 1987, 105, 1–102. [Google Scholar] [CrossRef]
- Damgaard, C.; Weiner, J. The need for alternative plant species interaction models. J. Plant Ecol. 2021, 14, 771–780. [Google Scholar] [CrossRef]
- Ogliari, P.J.; Andrade, D.F. Analysing longitudinal data via nonlinear models in randomized block designs. Comput. Stat. Data Anal. 2001, 36, 319–332. [Google Scholar] [CrossRef]
- Damgaard, C.F.; Irvine, K.M. Using the beta distribution to analyse plant cover data. J. Ecol. 2019, 107, 2747–2759. [Google Scholar] [CrossRef]
- Zhang, X.; Pei, Y.F.; Zhang, L.; Guo, B.; Pendegraft, A.H.; Zhuang, W.; Yi, N. Negative binomial mixed models for analyzing longitudinal microbiome data. Front. Microbiol. 2018, 9, 1683. [Google Scholar] [CrossRef]
- Brooks, M.E.; Kristensen, K.; Benthem, V.K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. glmmTMB Balances Speed and Flexibility Among Packages for Zero-inflated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Wickham, H.; Averick, M.; Bryan, J.; Chang, W.; McGowan, L.; François, R.; Grolemund, G.; Hayes, A.; Henry, L.; Hester, J.; et al. Welcome to the tidyverse. J. Open Source Softw. 2019, 4, 1686. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Hartig, F. 2022, DHARMa: Residual Diagnostics for Hierarchical (Multi-Level/Mixed) Regression Models. Available online: https://CRAN.R-project.org/package=DHARMa (accessed on 17 February 2024).
- Degn, H.J. Lyng og Græs på Randbøl Hede 2005—de Store Linier; Randbøl Statsskovdistrikt: København, Denmark, 2006; p. 12. [Google Scholar]
- Clrtap. Chapter III. Mapping critical levels for vegetation. In Convention on Long-Range Transboundary Air Pollution (CLRTAP) Manual on Methodologies and Criteria for Modelling and Mapping Critical Loads and Levels and Air Pollution Effects, Risks and Trends; Umweltbundesamt: Dessau-Roßlau, Germany, 2023; p. 338. [Google Scholar]
- Riis-Nielsen, T. Effects of Nitrogen on the Stability and Dynamics of Danish Heathland Vegetation. Ph.D. Thesis, University of Copenhagen, Copenhagen, Denmark, 1997. [Google Scholar]
- Jones, A.G.; Power, S.A. Field-scale evaluation of effects of nitrogen on the functioning of heathland ecosystems. J. Ecol. 2012, 100, 331–342. [Google Scholar] [CrossRef]
- Salemaa, M.; Makipää, R.; Oksanen, J. Differences in the growth response of three bryophyte species to nitrogen. Environ. Pollut. 2008, 152, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Haworth, B.J.; Ashmore, M.R.; Headley, A.D. Effects of Nitrogen Deposition on Bryophyte Species Composition of Calcareous Grasslands; Springer: Dordrecht, The Netherlands, 2007; pp. 111–117. [Google Scholar]
- Carroll, J.A.; Caporn, S.J.M.; Cawley, L.; Read, D.J.; Lee, J.A. The effect of increased deposition of atmospheric nitrogen on Calluna vulgaris in upland Britain. New Phytol. 1999, 141, 423–431. [Google Scholar] [CrossRef]
- Aerts, R. Competition between dominant plant species in heathlands. In Heathlands: Patterns and Processes in a Changing Environment; Aerts, R., Heil, G.W., Eds.; Kluwer Academic Publishers: London, UK, 1993; pp. 125–151. [Google Scholar]
- Aerts, R.; Berendse, F.; Caluwe, H.D.; Schmitz, M. Competition in Heathland along an Experimental Gradient of Nutrient Availability Published by: Wiley on behalf of Nordic Society Oikos Stable. Oikos 1990, 57, 310–318. Available online: https://www.jstor.org/stable/3565959 (accessed on 17 February 2024). [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Models for Summer | Year | Dose | Fence | Harvest | Year:Dose | Year:Fence | Year:Harvest | Dose:Fence | Dose:Harvest | Fence:Harvest | Year:Dose:Fence |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Grasses | * ↓ | * ↑ | |||||||||
| Dwarf shrubs | * ↓ | *** ↓ | |||||||||
| Empetrum nigrum | * ↓ | *** ↓ | |||||||||
| Avenella flexuosa | * ↑ | * ↓ | * ↑ | ||||||||
| Calluna vulgaris | *** ↓ | ** ↓ | ** ↑ | ||||||||
| Carex arenaria | * ↑ | * ↓ | * ↑ | ||||||||
| Lichens | ** ↓ | ||||||||||
| Bryophytes | *** ↑ | * ↓ | |||||||||
| Pleurozium schreberi | *** ↑ | ||||||||||
| Dicranum spp. | *** ↑ | *↑ | |||||||||
| Models for autumn | Year | Dose | Fence | Harvest | Year:Dose | Year:Fence | Year:Harvest | Dose:Fence | Dose:Harvest | Fence:Harvest | Year:Dose:Fence |
| Grasses | ** ↑ | ||||||||||
| Dwarf shrubs | *** ↓ | ||||||||||
| Empetrum nigrum | *** ↓ | ||||||||||
| Avenella flexuosa | ** ↑ | ||||||||||
| Calluna vulgaris | *** ↓ | ||||||||||
| Carex arenaria | * ↑ | ||||||||||
| Lichens | * ↓ | * ↑ | ** ↓ | ** ↓ | |||||||
| Bryophytes | ** ↓ | ** ↑ | * ↓ | ||||||||
| Pleurozium schreberi | * ↓ | * ↑ | * ↑ | * ↓ | |||||||
| Dicranum spp. | ** ↑ |
| Models for summer | Year | Dose | Fence | Harvest | Year:Dose | Year:Fence | Year:Harvest | Dose:Fence | Dose:Harvest | Fence:Harvest | Year:Dose:Fence |
|---|---|---|---|---|---|---|---|---|---|---|---|
| Grasses | * ↓ | ** ↑ | |||||||||
| Dwarf shrubs | *** ↓ | *** ↓ | |||||||||
| Empetrum nigrum | * ↓ | *** ↓ | |||||||||
| Avenella flexuosa | * ↑ | ** ↑ | |||||||||
| Calluna vulgaris | ** ↓ | ||||||||||
| Carex arenaria | ** ↑ | * ↓ | * ↑ | ||||||||
| Models for autumn | Year | Dose | Fence | Harvest | Year:Dose | Year:Fence | Year:Harvest | Dose:Fence | Dose:Harvest | Fence:Harvest | Year:Dose:Fence |
| Grasses | *** ↑ | ||||||||||
| Dwarf shrubs | *** ↓ | ||||||||||
| Empetrum nigrum | *** ↓ | ||||||||||
| Avenella flexuosa | *** ↑ | ||||||||||
| Calluna vulgaris | |||||||||||
| Carex arenaria | ** ↑ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaae, M.E.; Hu, F.; Bak, J.L.; Strandberg, M.T.; Damgaard, C.F. How Do Nitrogen Deposition, Mowing, and Deer Grazing Drive Vegetation Changes on Dune Heaths? Ecologies 2024, 5, 116-131. https://doi.org/10.3390/ecologies5010008
Kaae ME, Hu F, Bak JL, Strandberg MT, Damgaard CF. How Do Nitrogen Deposition, Mowing, and Deer Grazing Drive Vegetation Changes on Dune Heaths? Ecologies. 2024; 5(1):116-131. https://doi.org/10.3390/ecologies5010008
Chicago/Turabian StyleKaae, Mathias Emil, Fenjuan Hu, Jesper Leth Bak, Morten Tune Strandberg, and Christian Frølund Damgaard. 2024. "How Do Nitrogen Deposition, Mowing, and Deer Grazing Drive Vegetation Changes on Dune Heaths?" Ecologies 5, no. 1: 116-131. https://doi.org/10.3390/ecologies5010008