
The Role of Hypothalamic Neuropeptides in Regulation of Liver Functions in Health and Disease
 and
and
Abstract
:1. Introduction

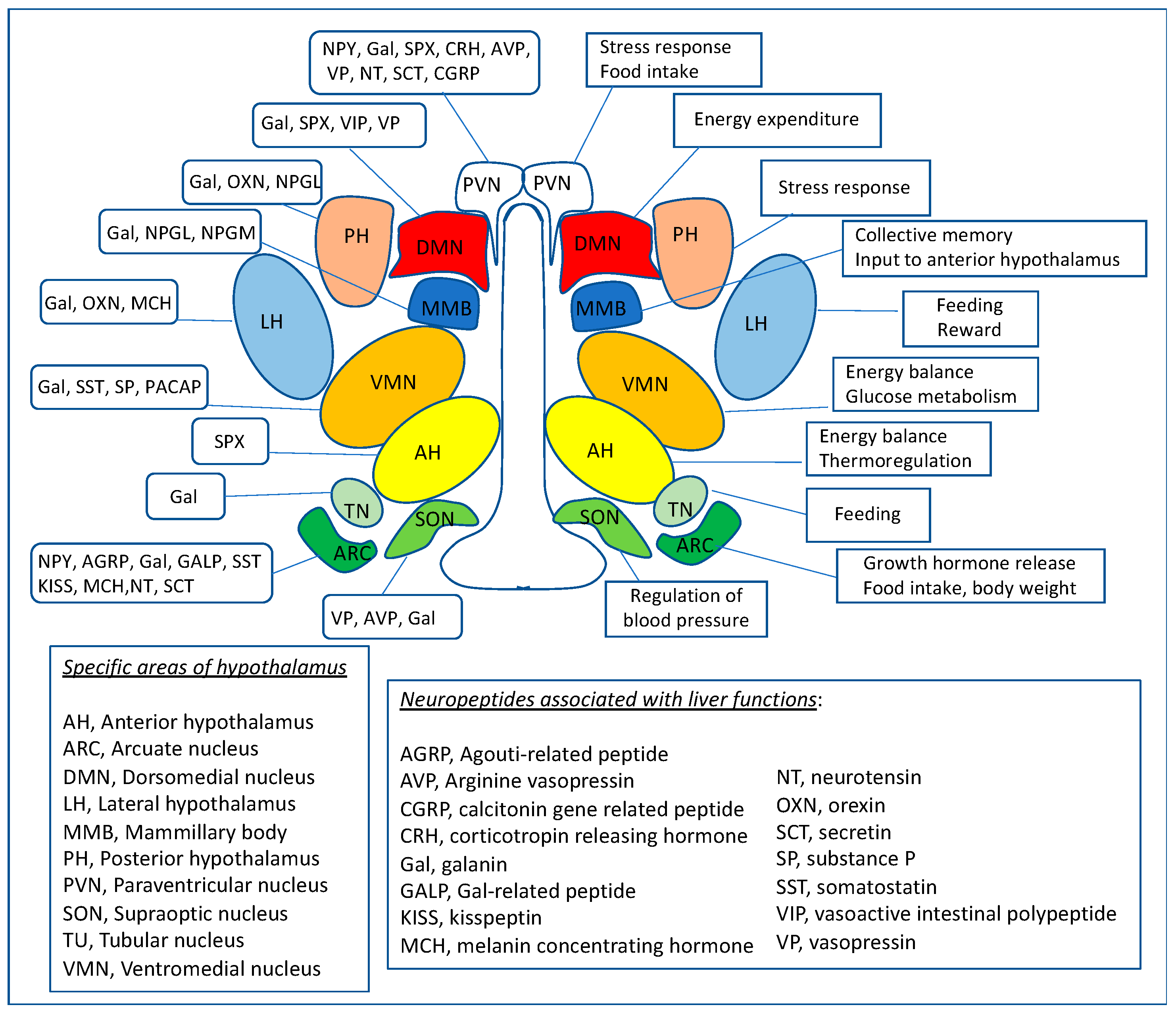{kind=link}
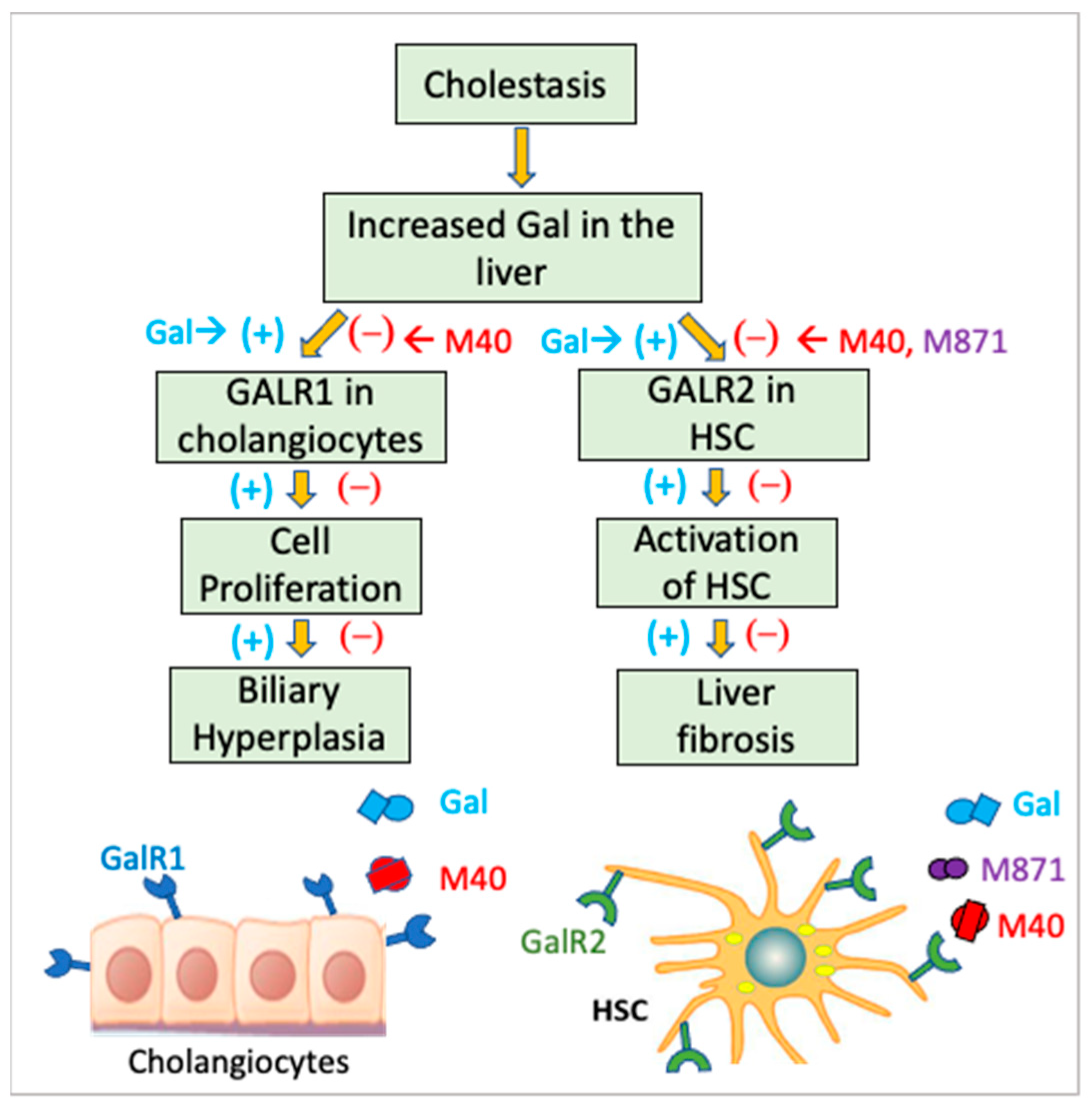{kind=link}
| Neuropeptide | Disease | Dysregulation/ Symptoms | Signaling Pathways/Therapies | References |
|---|---|---|---|---|
| Neuropeptide Y (NPY) | Obesity and NAFLD | NPY genetic variant rs16147 correlated with obesity metabolic syndrome, insulin resistance | NPY increases proinflammatory response | [22] |
| Liver steatosis | Increased intrahepatic NPY-immuno-reactive fibers. | NPY stimulated fat storage in liver and adipose via macrophage activation | [23] | |
| Agouti-related protein (AGRP) | Obesity and NAFLD | Upregulation of AGRP gene in obese mice | AGRP overexpression increased food intake | [24] |
| Galanin (Gal) | Obesity and hepatic steatosis | Higher than normal levels of serum and liver Gal. | Gal increased feeding rate and body weight. | [25] |
| Gal stimulated fat storage in adipose and liver | [26,27] | |||
| Orexins | NAFLD | Increased orexin in the liver | Orexin stimulated lipogenesis via ERK1/2 (Thr202/Tyr185) | [28] |
| Melanin-concentrating hormone (MCH) | NAFLD, obesity | Overeating and imbalanced energy homeostasis. | Antagonists of MCH receptors in obese mice reduced lipogenesis, inflammation, and fibrosis in liver | [29,30] |
| Neurotensin (NT) | NAFLD, metabolic syndrome | Higher than normal levels of plasma pro-NT in obesity | Increased NT enhanced fat absorption in the gut | [31] |
| Corticotropin releasing factor (CRF) | Hepatic steatosis, inflammation and fibrosis | CRF enhances steatosis and liver inflammation | CRF upregulated SREBP1, TNFα, and IL-1β by stimulation of sympathetic nervous system. | [32] |
| Somatostatin (SST) | Hepatic steatosis and NASH | Decreased SST in obesity-induced fatty liver disease | A synthetic analog of SST, octreotide, was tested as therapeutic strategy for high-fat diet-induced NASH | [33] |
| Neurosecretory peptide GL (NPGL) | Steatosis, NASH, metabolic syndrome | Precursor of NPGL is increased in hypothalamus of sugar-induced obesity | Unknown, still to be investigated | [34,35] |
| Secretin | NAFLD | Secretin (Sct)−/− and Sct-receptor−/− mice had less liver steatosis compared to wild-type mice when fed high-fat diet | Sct/SctReceptor/miR-125b axis promotes hepatic steatosis via Elov1 lipogenic gene upregulation | [36,37] |
2. Neuropeptide Y (NPY)
3. Agouti-Related Protein (AGRP) and Melanocortin Receptors
4. Galanin Family (Galanin, Galanin-like Peptide, Spexin, Kisspeptin)
4.1. Galanin (Gal)
4.2. Galanin-like Peptide (GALP)
4.3. Spexin (SPX)
4.4. Kisspeptin (KISS)
5. Orexins
6. Melanin Concentrating Hormone (MCH)
7. Corticotropin Releasing Hormone (CRH)
8. Somatostatin
9. Neurosecretory Protein GL (NPGL) and GM (NPGM)
10. Nesfatin-1
11. Substance P (SP)
12. Calcitonin Gene-Related Peptide (CGRP)
| Neuropeptide | Disease | Dysregulation/Symptoms | Signaling Pathways/ Therapies | References |
|---|---|---|---|---|
| Vasopressin | HRS, Cirrhosis-related hemorrhage | Variceal bleeding in cirrhotic patients | Unknown | [171,172] |
| Arginine Vasopressin (AVP) | Cirrhosis, hepatitis, HCC | Contributes to hepatic inflammation and fibrosis | Blocking AVP receptors may be therapeutic | [173] |
| Substance P | Autoimmune hepatitis | Increased SP-fibers in the liver, close to lymphocytes; | SP colocalization with TNFα and NF-κB in lymphocytes amplifies inflammation. | [174] |
| HCC | Chronic hepatic inflammation and fibrosis, cancer | Blocking SP–HSC–HCC axis causes reduction in HCC development. | [175] | |
| Cholestatic liver injury | Experimental cholestasis by BDL | Systemic application of SP reduced fibrogenic TGFβ and increased anti-inflammatory cytokines; it enhanced regulatory T cells. | [176] | |
| Somatostatin (SST) | Resection/transplantation of liver | Increased portal flow, liver failure | Exogeneous administration regulates portal flow, protects the liver | [177] |
| HCC | Chronic inflammation, fibrosis, carcinogenesis | SST and its analog reduce angiogenesis and hepatoma cell proliferation via apoptosis | [141] | |
| Polycystic liver disease (PLD) | Hepatic inflammation | SST decreases liver volume in PLD | [139] | |
| Pituitary adenylate cyclase-activating peptide (PACAP) | Liver ischemia-reperfusion injury | Acute hepatocyte death in liver transplantation | Promotes hepatocellular protection via CREB and KLF4-enhanced autophagy | [178,179] |
| Vasoactive intestinal peptide (VIP) | Cholestasis | Bile duct injuries and hyperbilirubinemia | Exogenous VIP restores damaged tight junctions in bile ducts | [180] |
| Corticotropin releasing factor (CRF) | Hepatic fibrosis | CRF administration in rats aggravates acute liver injury | CRF enhances TNFα and IL-1β, the latter by stimulation of sympathetic nervous system. | [32] |
| Secretin | Parenteral nutrition (PN)-associated liver disease (PNALD). | Rat model of PNALD: secretin reduced total bilirubin and bile acid | Secretin induces cAMP/PKA activation and increased cholangiocyte proliferation under liver injury | [181] |
| Cholangiocarcinoma | Dysregulation of cAMP/PKA pathway in cholangiocytes | Unknown | [182] |
13. Neurotensin (NT)
14. Secretin, Vasoactive Intestinal Peptide (VIP), and Pituitary Adenylate Cyclase-Activating Polypeptide (PACAP)
14.1. Secretin (SCT)
14.2. VIP
14.3. PACAP
15. Vasopressin and Arginine Vasopressin
16. Translational Studies on Hypothalamic Neuropeptides as Targets for Liver Disease Therapies
17. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Parker, J.A.; Bloom, S.R. Hypothalamic neuropeptides and the regulation of appetite. Neuropharmacology 2012, 63, 18–30. [Google Scholar] [CrossRef]
- Marcos, P.; Covenas, R. Neuropeptidergic Control of Feeding: Focus on the Galanin Family of Peptides. Int. J. Mol. Sci. 2021, 22, 2544. [Google Scholar] [CrossRef]
- Queen, N.J.; Hassan, Q.N., 2nd; Cao, L. Improvements to Healthspan Through Environmental Enrichment and Lifestyle Interventions: Where Are We Now? Front. Neurosci. 2020, 14, 605. [Google Scholar] [CrossRef] [PubMed]
- Petrescu, A.D.; Kain, J.; Liere, V.; Heavener, T.; DeMorrow, S. Hypothalamus-Pituitary-Adrenal Dysfunction in Cholestatic Liver Disease. Front. Endocrinol. 2018, 9, 660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herman, J.P.; McKlveen, J.M.; Ghosal, S.; Kopp, B.; Wulsin, A.; Makinson, R.; Scheimann, J.; Myers, B. Regulation of the Hypothalamic-Pituitary-Adrenocortical Stress Response. Compr. Physiol. 2016, 6, 603–621. [Google Scholar] [CrossRef] [Green Version]
- Kasprzak, A.; Adamek, A. The Neuropeptide System and Colorectal Cancer Liver Metastases: Mechanisms and Management. Int. J. Mol. Sci. 2020, 21, 3494. [Google Scholar] [CrossRef] [PubMed]
- Hillebrand, J.J.; de Wied, D.; Adan, R.A. Neuropeptides, food intake and body weight regulation: A hypothalamic focus. Peptides 2002, 23, 2283–2306. [Google Scholar] [CrossRef]
- Hokfelt, T.; Bartfai, T.; Bloom, F. Neuropeptides: Opportunities for drug discovery. Lancet Neurol. 2003, 2, 463–472. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Francis, H.; Zhou, T.; Meng, F.; Kennedy, L.; Ekser, B.; Baiocchi, L.; Onori, P.; Mancinelli, R.; Gaudio, E.; et al. Neuroendocrine Changes in Cholangiocarcinoma Growth. Cells 2020, 9, 436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrlich, L.; Scrushy, M.; Meng, F.; Lairmore, T.C.; Alpini, G.; Glaser, S. Biliary epithelium: A neuroendocrine compartment in cholestatic liver disease. Clin. Res. Hepatol. Gastroenterol. 2018, 42, 296–305. [Google Scholar] [CrossRef]
- DeMorrow, S.; Onori, P.; Venter, J.; Invernizzi, P.; Frampton, G.; White, M.; Franchitto, A.; Kopriva, S.; Bernuzzi, F.; Francis, H.; et al. Neuropeptide Y inhibits cholangiocarcinoma cell growth and invasion. Am. J. Physiol. Cell. Physiol. 2011, 300, C1078–C1089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeMorrow, S.; Meng, F.; Venter, J.; Leyva-Illades, D.; Francis, H.; Frampton, G.; Pae, H.Y.; Quinn, M.; Onori, P.; Glaser, S.; et al. Neuropeptide Y inhibits biliary hyperplasia of cholestatic rats by paracrine and autocrine mechanisms. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 305, G250–G257. [Google Scholar] [CrossRef] [Green Version]
- McMillin, M.; Frampton, G.; Grant, S.; DeMorrow, S. The Neuropeptide Galanin Is Up-Regulated during Cholestasis and Contributes to Cholangiocyte Proliferation. Am. J. Pathol. 2017, 187, 819–830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrescu, A.D.; Grant, S.; Williams, E.; Frampton, G.; Parks, N.; Blaney, H.; Davies, M.; John, R.; Reinhart, E.H.; McMillin, M.; et al. Coordinated Targeting of Galanin Receptors on Cholangiocytes and Hepatic Stellate Cells Ameliorates Liver Fibrosis in Multidrug Resistance Protein 2 Knockout Mice. Am. J. Pathol. 2020, 190, 586–601. [Google Scholar] [CrossRef] [PubMed]
- Li, R.Y.; Song, H.D.; Shi, W.J.; Hu, S.M.; Yang, Y.S.; Tang, J.F.; Chen, M.D.; Chen, J.L. Galanin inhibits leptin expression and secretion in rat adipose tissue and 3T3-L1 adipocytes. J. Mol. Endocrinol. 2004, 33, 11–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koller, A.; Brunner, S.M.; Bianchini, R.; Ramspacher, A.; Emberger, M.; Locker, F.; Schlager, S.; Kofler, B. Galanin is a potent modulator of cytokine and chemokine expression in human macrophages. Sci. Rep. 2019, 9, 7237. [Google Scholar] [CrossRef]
- Ramspacher, A.; Neudert, M.; Koller, A.; Schlager, S.; Kofler, B.; Brunner, S.M. Influence of the regulatory peptide galanin on cytokine expression in human monocytes. Ann. N. Y Acad. Sci. 2019, 1455, 185–195. [Google Scholar] [CrossRef] [Green Version]
- Jard, S. Vasopressin isoreceptors in the liver and kidney: Relationship between hormone binding and biological response. J. Physiol. 1981, 77, 621–628. [Google Scholar]
- He, L.; Li, Z.; Zhou, D.; Ding, Y.; Xu, L.; Chen, Y.; Fan, J. Galanin receptor 2 mediates antifibrogenic effects of galanin on hepatic stellate cells. Exp. Ther. Med. 2016, 12, 3375–3380. [Google Scholar] [CrossRef] [Green Version]
- Wu, N.; Meng, F.; Zhou, T.; Venter, J.; Giang, T.K.; Kyritsi, K.; Wu, C.; Alvaro, D.; Onori, P.; Mancinelli, R.; et al. The Secretin/Secretin Receptor Axis Modulates Ductular Reaction and Liver Fibrosis through Changes in Transforming Growth Factor-beta1-Mediated Biliary Senescence. Am. J. Pathol. 2018, 188, 2264–2280. [Google Scholar] [CrossRef] [Green Version]
- Moody, T.W.; Ramos-Alvarez, I.; Jensen, R.T. Neuropeptide G Protein-Coupled Receptors as Oncotargets. Front. Endocrinol. 2018, 9, 345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Luis, D.A.; Izaola, O.; de la Fuente, B.; Primo, D.; Aller, R. Association of Neuropeptide Y Gene rs16147 Polymorphism with Cardiovascular Risk Factors, Adipokines, and Metabolic Syndrome in Patients with Obesity. J. Nutr. Nutr. 2016, 9, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Komatsu, T.; Hayashi, H.; Mori, R.; Shimokawa, I. The Role of Neuropeptide Y in Adipocyte-Macrophage Crosstalk during High Fat Diet-Induced Adipose Inflammation and Liver Steatosis. Biomedicines 2021, 9, 1739. [Google Scholar] [CrossRef]
- Shutter, J.R.; Graham, M.; Kinsey, A.C.; Scully, S.; Luthy, R.; Stark, K.L. Hypothalamic expression of ART, a novel gene related to agouti, is up-regulated in obese and diabetic mutant mice. Genes Dev. 1997, 11, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Yun, R.; Dourmashkin, J.T.; Hill, J.; Gayles, E.C.; Fried, S.K.; Leibowitz, S.F. PVN galanin increases fat storage and promotes obesity by causing muscle to utilize carbohydrate more than fat. Peptides 2005, 26, 2265–2273. [Google Scholar] [CrossRef] [PubMed]
- Fang, P.; Yu, M.; Gu, X.; Shi, M.; Zhu, Y.; Zhang, Z.; Bo, P. Circulating galanin and galanin like peptide concentrations are correlated with increased triglyceride concentration in obese patients. Clin. Chim. Acta 2016, 461, 126–129. [Google Scholar] [CrossRef]
- Poritsanos, N.J.; Mizuno, T.M.; Lautatzis, M.E.; Vrontakis, M. Chronic increase of circulating galanin levels induces obesity and marked alterations in lipid metabolism similar to metabolic syndrome. Int. J. Obes. 2009, 33, 1381–1389. [Google Scholar] [CrossRef] [Green Version]
- Greene, E.S.; Zampiga, M.; Sirri, F.; Ohkubo, T.; Dridi, S. Orexin system is expressed in avian liver and regulates hepatic lipogenesis via ERK1/2 activation. Sci. Rep. 2020, 10, 19191. [Google Scholar] [CrossRef]
- Imbernon, M.; Beiroa, D.; Vazquez, M.J.; Morgan, D.A.; Veyrat-Durebex, C.; Porteiro, B.; Diaz-Arteaga, A.; Senra, A.; Busquets, S.; Velasquez, D.A.; et al. Central melanin-concentrating hormone influences liver and adipose metabolism via specific hypothalamic nuclei and efferent autonomic/JNK1 pathways. Gastroenterology 2013, 144, 636–649 e636. [Google Scholar] [CrossRef] [Green Version]
- Kawata, Y.; Okuda, S.; Hotta, N.; Igawa, H.; Takahashi, M.; Ikoma, M.; Kasai, S.; Ando, A.; Satomi, Y.; Nishida, M.; et al. A novel and selective melanin-concentrating hormone receptor 1 antagonist ameliorates obesity and hepatic steatosis in diet-induced obese rodent models. Eur. J. Pharmacol. 2017, 796, 45–53. [Google Scholar] [CrossRef]
- Barchetta, I.; Baroni, M.G.; Melander, O.; Cavallo, M.G. New Insights in the Control of Fat Homeostasis: The Role of Neurotensin. Int. J. Mol. Sci. 2022, 23, 2209. [Google Scholar] [CrossRef]
- Nakade, Y.; Kitano, R.; Yamauchi, T.; Kimoto, S.; Sakamoto, K.; Inoue, T.; Kobayashi, Y.; Ohashi, T.; Sumida, Y.; Ito, K.; et al. Effect of Central Corticotropin-Releasing Factor on Hepatic Lipid Metabolism and Inflammation-Related Gene Expression in Rats. Int. J. Mol. Sci. 2021, 22, 3940. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.X.; Ye, T.; Li, M.; Li, X.; Qiang, O.; Tang, C.W.; Liu, R. Effects of octreotide on hepatic glycogenesis in rats with high fat diet-induced obesity. Mol. Med. Rep. 2017, 16, 109–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Narimatsu, Y.; Iwakoshi-Ukena, E.; Fukumura, K.; Shikano, K.; Furumitsu, M.; Morishita, M.; Bentley, G.E.; Kriegsfeld, L.J.; Ukena, K. Hypothalamic Overexpression of Neurosecretory Protein GL Leads to Obesity in Male C57BL/6J Mice. Neuroendocrinology 2022, 112, 606–620. [Google Scholar] [CrossRef] [PubMed]
- Narimatsu, Y.; Iwakoshi-Ukena, E.; Naito, M.; Moriwaki, S.; Furumitsu, M.; Ukena, K. Neurosecretory Protein GL Accelerates Liver Steatosis in Mice Fed Medium-Fat/Medium-Fructose Diet. Int. J. Mol. Sci. 2022, 23, 2071. [Google Scholar] [CrossRef]
- Wu, N.; Baiocchi, L.; Zhou, T.; Kennedy, L.; Ceci, L.; Meng, F.; Sato, K.; Wu, C.; Ekser, B.; Kyritsi, K.; et al. Functional Role of the Secretin/Secretin Receptor Signaling During Cholestatic Liver Injury. Hepatology 2020, 72, 2219–2227. [Google Scholar] [CrossRef]
- Chen, L.; Wu, N.; Kennedy, L.; Francis, H.; Ceci, L.; Zhou, T.; Samala, N.; Kyritsi, K.; Wu, C.; Sybenga, A.; et al. Inhibition of Secretin/Secretin Receptor Axis Ameliorates NAFLD Phenotypes. Hepatology 2021, 74, 1845–1863. [Google Scholar] [CrossRef]
- Pedragosa-Badia, X.; Stichel, J.; Beck-Sickinger, A.G. Neuropeptide Y receptors: How to get subtype selectivity. Front. Endocrinol. 2013, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Yi, M.; Li, H.; Wu, Z.; Yan, J.; Liu, Q.; Ou, C.; Chen, M. A Promising Therapeutic Target for Metabolic Diseases: Neuropeptide Y Receptors in Humans. Cell. Physiol. Biochem. 2018, 45, 88–107. [Google Scholar] [CrossRef]
- Chronwall, B.M.; DiMaggio, D.A.; Massari, V.J.; Pickel, V.M.; Ruggiero, D.A.; O’Donohue, T.L. The anatomy of neuropeptide-Y-containing neurons in rat brain. Neuroscience 1985, 15, 1159–1181. [Google Scholar] [CrossRef]
- Durkin, M.M.; Walker, M.W.; Smith, K.E.; Gustafson, E.L.; Gerald, C.; Branchek, T.A. Expression of a novel neuropeptide Y receptor subtype involved in food intake: An in situ hybridization study of Y5 mRNA distribution in rat brain. Exp. Neurol. 2000, 165, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Ferrier, L.; Segain, J.P.; Bonnet, C.; Cherbut, C.; Lehur, P.A.; Jarry, A.; Galmiche, J.P.; Blottiere, H.M. Functional mapping of NPY/PYY receptors in rat and human gastro-intestinal tract. Peptides 2002, 23, 1765–1771. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, C.Y.; Chen, W.C.; Shi, Y.C.; Wang, C.M.; Lin, S.; He, H.F. Regulation of neuropeptide Y in body microenvironments and its potential application in therapies: A review. Cell Biosci. 2021, 11, 151. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; He, H.; Cheng, Z.; Bai, Y.; Ma, X. The Role of Neuropeptide Y and Peptide YY in the Development of Obesity via Gut-brain Axis. Curr. Protein Pept. Sci. 2019, 20, 750–758. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.G.; Fujimura, M.; Mori, A.; Tooyama, I.; Kimura, H. Light and electron microscopy of neuropeptide Y-containing nerves in human liver, gallbladder, and pancreas. Gastroenterology 1991, 101, 1054–1059. [Google Scholar] [CrossRef] [PubMed]
- Yue, Y.; Xu, D.; Wang, Y.; Wang, X.; Xia, F. Effect of inducible nitric oxide synthase and neuropeptide Y in plasma and placentas from intrahepatic cholestasis of pregnancy. J. Obstet. Gynaecol. Res. 2018, 44, 1377–1383. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Wang, J.; Yan, S.; Zhao, J.; Li, H. Expression of neuropeptide Y and pro-opiomelanocortin in hypothalamic arcuate nucleus in 17alpha-ethinyl estradiol-induced intrahepatic cholestasis pregnant rat offspring. J. Obstet. Gynaecol. Res. 2014, 40, 445–452. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Lin, X.; Lin, S. Neuropeptide Y and Metabolism Syndrome: An Update on Perspectives of Clinical Therapeutic Intervention Strategies. Front. Cell Dev. Biol. 2021, 9, 695623. [Google Scholar] [CrossRef]
- Donnelly, K.L.; Smith, C.I.; Schwarzenberg, S.J.; Jessurun, J.; Boldt, M.D.; Parks, E.J. Sources of fatty acids stored in liver and secreted via lipoproteins in patients with nonalcoholic fatty liver disease. J. Clin. Investig. 2005, 115, 1343–1351. [Google Scholar] [CrossRef]
- Kuo, L.E.; Kitlinska, J.B.; Tilan, J.U.; Li, L.; Baker, S.B.; Johnson, M.D.; Lee, E.W.; Burnett, M.S.; Fricke, S.T.; Kvetnansky, R.; et al. Neuropeptide Y acts directly in the periphery on fat tissue and mediates stress-induced obesity and metabolic syndrome. Nat. Med. 2007, 13, 803–811. [Google Scholar] [CrossRef]
- Zhang, W.; Cline, M.A.; Gilbert, E.R. Hypothalamus-adipose tissue crosstalk: Neuropeptide Y and the regulation of energy metabolism. Nutr. Metab. 2014, 11, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, F.; Zhou, Y.; Yang, K.; Shen, M.; Wang, Y. NPY stimulates cholesterol synthesis acutely by activating the SREBP2-HMGCR pathway through the Y1 and Y5 receptors in murine hepatocytes. Life Sci. 2020, 262, 118478. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, C.; Klein, S.; Reul, W.H.; Magdaleno, F.; Groschl, S.; Dietrich, P.; Schierwagen, R.; Uschner, F.E.; Torres, S.; Hieber, C.; et al. Neprilysin-dependent neuropeptide Y cleavage in the liver promotes fibrosis by blocking NPY-receptor 1. Cell Rep. 2023, 42, 112059. [Google Scholar] [CrossRef]
- Dietrich, P.; Wormser, L.; Fritz, V.; Seitz, T.; De Maria, M.; Schambony, A.; Kremer, A.E.; Gunther, C.; Itzel, T.; Thasler, W.E.; et al. Molecular crosstalk between Y5 receptor and neuropeptide Y drives liver cancer. J. Clin. Investig. 2020, 130, 2509–2526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenfeld, R.D.; Zeni, L.; Welcher, A.A.; Narhi, L.O.; Hale, C.; Marasco, J.; Delaney, J.; Gleason, T.; Philo, J.S.; Katta, V.; et al. Biochemical, biophysical, and pharmacological characterization of bacterially expressed human agouti-related protein. Biochemistry 1998, 37, 16041–16052. [Google Scholar] [CrossRef]
- Yang, Y.K.; Ollmann, M.M.; Wilson, B.D.; Dickinson, C.; Yamada, T.; Barsh, G.S.; Gantz, I. Effects of recombinant agouti-signaling protein on melanocortin action. Mol. Endocrinol. 1997, 11, 274–280. [Google Scholar] [CrossRef]
- Ollmann, M.M.; Wilson, B.D.; Yang, Y.K.; Kerns, J.A.; Chen, Y.; Gantz, I.; Barsh, G.S. Antagonism of central melanocortin receptors in vitro and in vivo by agouti-related protein. Science 1997, 278, 135–138. [Google Scholar] [CrossRef]
- Qian, S.; Chen, H.; Weingarth, D.; Trumbauer, M.E.; Novi, D.E.; Guan, X.; Yu, H.; Shen, Z.; Feng, Y.; Frazier, E.; et al. Neither agouti-related protein nor neuropeptide Y is critically required for the regulation of energy homeostasis in mice. Mol. Cell. Biol. 2002, 22, 5027–5035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butler, A.A. The melanocortin system and energy balance. Peptides 2006, 27, 281–290. [Google Scholar] [CrossRef] [Green Version]
- Albarado, D.C.; McClaine, J.; Stephens, J.M.; Mynatt, R.L.; Ye, J.; Bannon, A.W.; Richards, W.G.; Butler, A.A. Impaired coordination of nutrient intake and substrate oxidation in melanocortin-4 receptor knockout mice. Endocrinology 2004, 145, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Zhang, Y.; Rodrigues, E.; Zheng, D.; Matheny, M.; Cheng, K.Y.; Scarpace, P.J. Melanocortin activation of nucleus of the solitary tract avoids anorectic tachyphylaxis and induces prolonged weight loss. Am. J. Physiol. Endocrinol. Metab. 2007, 293, E252–E258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, G.; Zhang, Y.; Cheng, K.Y.; Scarpace, P.J. Lean rats with hypothalamic pro-opiomelanocortin overexpression exhibit greater diet-induced obesity and impaired central melanocortin responsiveness. Diabetologia 2007, 50, 1490–1499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leckstrom, A.; Lew, P.S.; Poritsanos, N.J.; Mizuno, T.M. Central melanocortin receptor agonist reduces hepatic lipogenic gene expression in streptozotocin-induced diabetic mice. Life Sci. 2011, 88, 664–669. [Google Scholar] [CrossRef] [PubMed]
- Leckstrom, A.; Lew, P.S.; Poritsanos, N.J.; Mizuno, T.M. Treatment with a melanocortin agonist improves abnormal lipid metabolism in streptozotocin-induced diabetic mice. Neuropeptides 2011, 45, 123–129. [Google Scholar] [CrossRef]
- Itoh, M.; Suganami, T.; Nakagawa, N.; Tanaka, M.; Yamamoto, Y.; Kamei, Y.; Terai, S.; Sakaida, I.; Ogawa, Y. Melanocortin 4 receptor-deficient mice as a novel mouse model of nonalcoholic steatohepatitis. Am. J. Pathol. 2011, 179, 2454–2463. [Google Scholar] [CrossRef]
- Itoh, M.; Kato, H.; Suganami, T.; Konuma, K.; Marumoto, Y.; Terai, S.; Sakugawa, H.; Kanai, S.; Hamaguchi, M.; Fukaishi, T.; et al. Hepatic crown-like structure: A unique histological feature in non-alcoholic steatohepatitis in mice and humans. PLoS ONE 2013, 8, e82163. [Google Scholar] [CrossRef] [Green Version]
- Itoh, M.; Suganami, T.; Kato, H.; Kanai, S.; Shirakawa, I.; Sakai, T.; Goto, T.; Asakawa, M.; Hidaka, I.; Sakugawa, H.; et al. CD11c+ resident macrophages drive hepatocyte death-triggered liver fibrosis in a murine model of nonalcoholic steatohepatitis. JCI Insight 2017, 2, e92902. [Google Scholar] [CrossRef]
- Langel, U. Galanin receptor ligands. Springerplus 2015, 4, L18. [Google Scholar] [CrossRef] [Green Version]
- Lang, R.; Gundlach, A.L.; Holmes, F.E.; Hobson, S.A.; Wynick, D.; Hokfelt, T.; Kofler, B. Physiology, signaling, and pharmacology of galanin peptides and receptors: Three decades of emerging diversity. Pharmacol. Rev. 2015, 67, 118–175. [Google Scholar] [CrossRef] [Green Version]
- Amiranoff, B.; Lorinet, A.M.; Laburthe, M. A clonal rat pancreatic delta cell line (Rin14B) expresses a high number of galanin receptors negatively coupled to a pertussis-toxin-sensitive cAMP-production pathway. Eur. J. Biochem. 1991, 195, 459–463. [Google Scholar] [CrossRef]
- Parker, E.M.; Izzarelli, D.G.; Nowak, H.P.; Mahle, C.D.; Iben, L.G.; Wang, J.; Goldstein, M.E. Cloning and characterization of the rat GALR1 galanin receptor from Rin14B insulinoma cells. Brain Res. Mol. Brain Res. 1995, 34, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Webling, K.E.; Runesson, J.; Bartfai, T.; Langel, U. Galanin receptors and ligands. Front. Endocrinol. 2012, 3, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatemoto, K.; Rokaeus, A.; Jornvall, H.; McDonald, T.J.; Mutt, V. Galanin-a novel biologically active peptide from porcine intestine. FEBS Lett. 1983, 164, 124–128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, R.; Gundlach, A.L.; Kofler, B. The galanin peptide family: Receptor pharmacology, pleiotropic biological actions, and implications in health and disease. Pharmacol. Ther. 2007, 115, 177–207. [Google Scholar] [CrossRef] [PubMed]
- Kyrkouli, S.E.; Stanley, B.G.; Seirafi, R.D.; Leibowitz, S.F. Stimulation of feeding by galanin: Anatomical localization and behavioral specificity of this peptide’s effects in the brain. Peptides 1990, 11, 995–1001. [Google Scholar] [CrossRef]
- Fabbrini, E.; Sullivan, S.; Klein, S. Obesity and nonalcoholic fatty liver disease: Biochemical, metabolic, and clinical implications. Hepatology 2010, 51, 679–689. [Google Scholar] [CrossRef] [Green Version]
- Fabbrini, E.; Magkos, F.; Mohammed, B.S.; Pietka, T.; Abumrad, N.A.; Patterson, B.W.; Okunade, A.; Klein, S. Intrahepatic fat, not visceral fat, is linked with metabolic complications of obesity. Proc. Natl. Acad. Sci. USA 2009, 106, 15430–15435. [Google Scholar] [CrossRef] [Green Version]
- Acar, S.; Paketci, A.; Kume, T.; Demir, K.; Gursoy Calan, O.; Bober, E.; Abaci, A. Positive correlation of galanin with insulin resistance and triglyceride levels in obese children. Turk. J. Med. Sci. 2018, 48, 560–568. [Google Scholar] [CrossRef]
- Alpini, G.; Lenzi, R.; Sarkozi, L.; Tavoloni, N. Biliary physiology in rats with bile ductular cell hyperplasia. Evidence for a secretory function of proliferated bile ductules. J. Clin. Investig. 1988, 81, 569–578. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Qiao, L.; Wang, B. Primary biliary cirrhosis is a generalized autoimmune epithelitis. Int. J. Mol. Sci. 2015, 16, 6432–6446. [Google Scholar] [CrossRef] [Green Version]
- Dong, M.; Li, J.; Tang, R.; Zhu, P.; Qiu, F.; Wang, C.; Qiu, J.; Wang, L.; Dai, Y.; Xu, P.; et al. Multiple genetic variants associated with primary biliary cirrhosis in a Han Chinese population. Clin. Rev. Allergy Immunol. 2015, 48, 316–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohtaki, T.; Kumano, S.; Ishibashi, Y.; Ogi, K.; Matsui, H.; Harada, M.; Kitada, C.; Kurokawa, T.; Onda, H.; Fujino, M. Isolation and cDNA cloning of a novel galanin-like peptide (GALP) from porcine hypothalamus. J. Biol. Chem. 1999, 274, 37041–37045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lang, R.; Berger, A.; Santic, R.; Geisberger, R.; Hermann, A.; Herzog, H.; Kofler, B. Pharmacological and functional characterization of galanin-like peptide fragments as potent galanin receptor agonists. Neuropeptides 2005, 39, 179–184. [Google Scholar] [CrossRef]
- Hirako, S.; Wada, N.; Kageyama, H.; Takenoya, F.; Izumida, Y.; Kim, H.; Iizuka, Y.; Matsumoto, A.; Okabe, M.; Kimura, A.; et al. Autonomic nervous system-mediated effects of galanin-like peptide on lipid metabolism in liver and adipose tissue. Sci. Rep. 2016, 6, 21481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takenoya, F.; Hirako, S.; Wada, N.; Nonaka, N.; Hirabayashi, T.; Kageyama, H.; Shioda, S. Regulation of Feeding Behavior and Energy Metabolism by Galanin-like Peptide (GALP): A Novel Strategy to Fight Against Obesity. Curr. Pharm. Des. 2018, 24, 3926–3933. [Google Scholar] [CrossRef] [PubMed]
- Ma, A.; Bai, J.; He, M.; Wong, A.O.L. Spexin as a neuroendocrine signal with emerging functions. Gen. Comp. Endocrinol. 2018, 265, 90–96. [Google Scholar] [CrossRef]
- Zheng, B.; Li, S.; Liu, Y.; Li, Y.; Chen, H.; Tang, H.; Liu, X.; Lin, H.; Zhang, Y.; Cheng, C.H.K. Spexin Suppress Food Intake in Zebrafish: Evidence from Gene Knockout Study. Sci. Rep. 2017, 7, 14643. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.K.; Yun, S.; Son, G.H.; Hwang, J.I.; Park, C.R.; Kim, J.I.; Kim, K.; Vaudry, H.; Seong, J.Y. Coevolution of the spexin/galanin/kisspeptin family: Spexin activates galanin receptor type II and III. Endocrinology 2014, 155, 1864–1873. [Google Scholar] [CrossRef] [Green Version]
- Walewski, J.L.; Ge, F.; Lobdell, H.t.; Levin, N.; Schwartz, G.J.; Vasselli, J.R.; Pomp, A.; Dakin, G.; Berk, P.D. Spexin is a novel human peptide that reduces adipocyte uptake of long chain fatty acids and causes weight loss in rodents with diet-induced obesity. Obesity 2014, 22, 1643–1652. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Hossain, J.; Nader, N.; Aguirre, R.; Sriram, S.; Balagopal, P.B. Decreased Circulating Levels of Spexin in Obese Children. J. Clin. Endocrinol. Metab. 2016, 101, 2931–2936. [Google Scholar] [CrossRef] [Green Version]
- Gu, L.; Ma, Y.; Gu, M.; Zhang, Y.; Yan, S.; Li, N.; Wang, Y.; Ding, X.; Yin, J.; Fan, N.; et al. Spexin peptide is expressed in human endocrine and epithelial tissues and reduced after glucose load in type 2 diabetes. Peptides 2015, 71, 232–239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karaca, A.; Bakar-Ates, F.; Ersoz-Gulcelik, N. Decreased Spexin Levels in Patients with Type 1 and Type 2 Diabetes. Med. Princ. Pract. 2018, 27, 549–554. [Google Scholar] [CrossRef] [PubMed]
- Ge, J.F.; Walewski, J.L.; Anglade, D.; Berk, P.D. Regulation of Hepatocellular Fatty Acid Uptake in Mouse Models of Fatty Liver Disease with and without Functional Leptin Signaling: Roles of NfKB and SREBP-1C and the Effects of Spexin. Semin. Liver Dis. 2016, 36, 360–372. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Li, G.; She, Y.; Zhang, Z. Low levels of spexin and adiponectin may predict insulin resistance in patients with non-alcoholic fatty liver. Pract. Lab. Med. 2021, 24, e00207. [Google Scholar] [CrossRef] [PubMed]
- Gu, L.; Ding, X.; Wang, Y.; Gu, M.; Zhang, J.; Yan, S.; Li, N.; Song, Z.; Yin, J.; Lu, L.; et al. Spexin alleviates insulin resistance and inhibits hepatic gluconeogenesis via the FoxO1/PGC-1alpha pathway in high-fat-diet-induced rats and insulin resistant cells. Int. J. Biol. Sci. 2019, 15, 2815–2829. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.Y.; Zhao, L.; Huang, T.; Lu, L.; Khan, M.; Liu, J.; Zhong, L.L.D.; Cai, Z.W.; Fan, B.M.; Wong, A.O.L.; et al. Spexin Acts as Novel Regulator for Bile Acid Synthesis. Front. Physiol. 2018, 9, 378. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.Y.; Huang, T.; Zhao, L.; Zhong, L.L.D.; Lam, W.C.; Fan, B.M.; Bian, Z.X. Circulating Spexin Levels Negatively Correlate With Age, BMI, Fasting Glucose, and Triglycerides in Healthy Adult Women. J. Endocr. Soc. 2018, 2, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.K.; Nguyen, T.; O’Neill, G.P.; Cheng, R.; Liu, Y.; Howard, A.D.; Coulombe, N.; Tan, C.P.; Tang-Nguyen, A.T.; George, S.R.; et al. Discovery of a receptor related to the galanin receptors. FEBS Lett. 1999, 446, 103–107. [Google Scholar] [CrossRef] [Green Version]
- Kotani, M.; Detheux, M.; Vandenbogaerde, A.; Communi, D.; Vanderwinden, J.M.; Le Poul, E.; Brezillon, S.; Tyldesley, R.; Suarez-Huerta, N.; Vandeput, F.; et al. The metastasis suppressor gene KiSS-1 encodes kisspeptins, the natural ligands of the orphan G protein-coupled receptor GPR54. J. Biol. Chem. 2001, 276, 34631–34636. [Google Scholar] [CrossRef] [Green Version]
- Muir, A.I.; Chamberlain, L.; Elshourbagy, N.A.; Michalovich, D.; Moore, D.J.; Calamari, A.; Szekeres, P.G.; Sarau, H.M.; Chambers, J.K.; Murdock, P.; et al. AXOR12, a novel human G protein-coupled receptor, activated by the peptide KiSS-1. J. Biol. Chem. 2001, 276, 28969–28975. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, Y.; Sakamoto, M.; Fujii, G.; Tsuiji, H.; Kenetaka, K.; Asaka, M.; Hirohashi, S. Overexpression of orphan G-protein-coupled receptor, Gpr49, in human hepatocellular carcinomas with beta-catenin mutations. Hepatology 2003, 37, 528–533. [Google Scholar] [CrossRef]
- Ikeguchi, M.; Hirooka, Y.; Kaibara, N. Quantitative reverse transcriptase polymerase chain reaction analysis for KiSS-1 and orphan G-protein-coupled receptor (hOT7T175) gene expression in hepatocellular carcinoma. J. Cancer Res. Clin. Oncol. 2003, 129, 531–535. [Google Scholar] [CrossRef]
- Wang, T.; Cui, X.; Xie, L.; Xing, R.; You, P.; Zhao, Y.; Yang, Y.; Xu, Y.; Zeng, L.; Chen, H.; et al. Kisspeptin Receptor GPR54 Promotes Adipocyte Differentiation and Fat Accumulation in Mice. Front. Physiol. 2018, 9, 209. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Fu, W.; Huang, Y.; Ni, Y. Effects of kisspeptin-10 on lipid metabolism in cultured chicken hepatocytes. Asian-Australas. J. Anim. Sci. 2012, 25, 1229–1236. [Google Scholar] [CrossRef] [PubMed]
- Flynn, C.R.; Albaugh, V.L.; Tamboli, R.A.; Gregory, J.M.; Bosompem, A.; Sidani, R.M.; Winnick, J.J. Roux-en-Y gastric bypass surgery improves hepatic glucose metabolism and reduces plasma kisspeptin levels in morbidly obese patients with type 2 diabetes. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 318, G370–G374. [Google Scholar] [CrossRef] [PubMed]
- Guzman, S.; Dragan, M.; Kwon, H.; de Oliveira, V.; Rao, S.; Bhatt, V.; Kalemba, K.M.; Shah, A.; Rustgi, V.K.; Wang, H.; et al. Targeting hepatic kisspeptin receptor ameliorates non-alcoholic fatty liver disease in a mouse model. J. Clin. Investig. 2022, 132, e145889. [Google Scholar] [CrossRef]
- Hudson, A.D.; Kauffman, A.S. Metabolic actions of kisspeptin signaling: Effects on body weight, energy expenditure, and feeding. Pharmacol. Ther. 2022, 231, 107974. [Google Scholar] [CrossRef]
- Sakurai, T.; Amemiya, A.; Ishii, M.; Matsuzaki, I.; Chemelli, R.M.; Tanaka, H.; Williams, S.C.; Richardson, J.A.; Kozlowski, G.P.; Wilson, S.; et al. Orexins and orexin receptors: A family of hypothalamic neuropeptides and G protein-coupled receptors that regulate feeding behavior. Cell 1998, 92, 573–585. [Google Scholar] [CrossRef] [Green Version]
- de Lecea, L.; Kilduff, T.S.; Peyron, C.; Gao, X.; Foye, P.E.; Danielson, P.E.; Fukuhara, C.; Battenberg, E.L.; Gautvik, V.T.; Bartlett, F.S., 2nd; et al. The hypocretins: Hypothalamus-specific peptides with neuroexcitatory activity. Proc. Natl. Acad. Sci. USA 1998, 95, 322–327. [Google Scholar] [CrossRef] [Green Version]
- Sakurai, T. Orexins and orexin receptors: Implication in feeding behavior. Regul. Pept. 1999, 85, 25–30. [Google Scholar] [CrossRef]
- Ju, S.J.; Zhao, Y.; Chang, X.; Guo, L. Orexin A protects cells from apoptosis by regulating FoxO1 and mTORC1 through the OX1R/PI3K/AKT signaling pathway in hepatocytes. Int. J. Mol. Med. 2014, 34, 153–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, X.; Liu, Y.; Zhao, Y.; Sun, X.; Fan, D.; Guo, L. Orexin A affects HepG2 human hepatocellular carcinoma cells glucose metabolism via HIF-1alpha-dependent and -independent mechanism. PLoS ONE 2017, 12, e0184213. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; He, T.; Wan, B.; Wang, X.; Zhang, L. Orexin A ameliorates HBV X protein-induced cytotoxicity and inflammatory response in human hepatocytes. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2003–2009. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomori, A.; Ishihara, A.; Ito, M.; Matsushita, H.; Ito, M.; Mashiko, S.; Iwaasa, H.; Matsuda, M.; Bednarek, M.A.; Qian, S.; et al. Blockade of MCH1 receptor signalling ameliorates obesity and related hepatic steatosis in ovariectomized mice. Br. J. Pharmacol. 2007, 151, 900–908. [Google Scholar] [CrossRef] [Green Version]
- Chee, M.J.; Pissios, P.; Prasad, D.; Maratos-Flier, E. Expression of melanin-concentrating hormone receptor 2 protects against diet-induced obesity in male mice. Endocrinology 2014, 155, 81–88. [Google Scholar] [CrossRef] [PubMed]
- Otieno, M.A.; Bhaskaran, V.; Janovitz, E.; Callejas, Y.; Foster, W.B.; Washburn, W.; Megill, J.R.; Lehman-McKeeman, L.; Gemzik, B. Mechanisms for Hepatobiliary Toxicity in Rats Treated with an Antagonist of Melanin Concentrating Hormone Receptor 1 (MCHR1). Toxicol. Sci. 2017, 155, 379–388. [Google Scholar] [CrossRef] [Green Version]
- Imbernon, M.; Sanchez-Rebordelo, E.; Romero-Pico, A.; Kallo, I.; Chee, M.J.; Porteiro, B.; Al-Massadi, O.; Contreras, C.; Ferno, J.; Senra, A.; et al. Hypothalamic kappa opioid receptor mediates both diet-induced and melanin concentrating hormone-induced liver damage through inflammation and endoplasmic reticulum stress. Hepatology 2016, 64, 1086–1104. [Google Scholar] [CrossRef] [Green Version]
- Caruso, A.; Gaetano, A.; Scaccianoce, S. Corticotropin-Releasing Hormone: Biology and Therapeutic Opportunities. Biology 2022, 11, 1785. [Google Scholar] [CrossRef]
- Autelitano, D.J.; Lundblad, J.R.; Blum, M.; Roberts, J.L. Hormonal regulation of POMC gene expression. Annu. Rev. Physiol. 1989, 51, 715–726. [Google Scholar] [CrossRef]
- Wardlaw, S.L. Hypothalamic proopiomelanocortin processing and the regulation of energy balance. Eur. J. Pharmacol. 2011, 660, 213–219. [Google Scholar] [CrossRef] [Green Version]
- Liang, Q.; Zhong, L.; Zhang, J.; Wang, Y.; Bornstein, S.R.; Triggle, C.R.; Ding, H.; Lam, K.S.; Xu, A. FGF21 maintains glucose homeostasis by mediating the cross talk between liver and brain during prolonged fasting. Diabetes 2014, 63, 4064–4075. [Google Scholar] [CrossRef] [Green Version]
- McMillin, M.; DeMorrow, S. Effects of bile acids on neurological function and disease. FASEB J. 2016, 30, 3658–3668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quinn, M.; McMillin, M.; Galindo, C.; Frampton, G.; Pae, H.Y.; DeMorrow, S. Bile acids permeabilize the blood brain barrier after bile duct ligation in rats via Rac1-dependent mechanisms. Dig. Liver Dis. 2014, 46, 527–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMillin, M.; Frampton, G.; Quinn, M.; Divan, A.; Grant, S.; Patel, N.; Newell-Rogers, K.; DeMorrow, S. Suppression of the HPA Axis During Cholestasis Can Be Attributed to Hypothalamic Bile Acid Signaling. Mol. Endocrinol. 2015, 29, 1720–1730. [Google Scholar] [CrossRef] [Green Version]
- Dautzenberg, F.M.; Hauger, R.L. The CRF peptide family and their receptors: Yet more partners discovered. Trends Pharmacol. Sci. 2002, 23, 71–77. [Google Scholar] [CrossRef]
- Grammatopoulos, D.K. Insights into mechanisms of corticotropin-releasing hormone receptor signal transduction. Br. J. Pharmacol. 2012, 166, 85–97. [Google Scholar] [CrossRef] [Green Version]
- Chrousos, G.P.; Zoumakis, E. Milestones in CRH Research. Curr. Mol. Pharmacol. 2017, 10, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.C.; Truax, A.D.; Giamouridis, D.; Lai, N.C.; Guo, T.; Hammond, H.K.; Gao, M.H. Significant alteration of liver metabolites by AAV8.Urocortin 2 gene transfer in mice with insulin resistance. PLoS ONE 2019, 14, e0224428. [Google Scholar] [CrossRef] [Green Version]
- Gao, M.H.; Giamouridis, D.; Lai, N.C.; Walenta, E.; Paschoal, V.A.; Kim, Y.C.; Miyanohara, A.; Guo, T.; Liao, M.; Liu, L.; et al. One-time injection of AAV8 encoding urocortin 2 provides long-term resolution of insulin resistance. JCI Insight 2016, 1, e88322. [Google Scholar] [CrossRef] [Green Version]
- Paruthiyil, S.; Hagiwara, S.I.; Kundassery, K.; Bhargava, A. Sexually dimorphic metabolic responses mediated by CRF(2) receptor during nutritional stress in mice. Biol. Sex. Differ. 2018, 9, 49. [Google Scholar] [CrossRef] [Green Version]
- Patel, Y.C. Somatostatin and its receptor family. Front. Neuroendocrinol. 1999, 20, 157–198. [Google Scholar] [CrossRef]
- Minami, S.; Kamegai, J.; Sugihara, H.; Suzuki, N.; Wakabayashi, I. Growth hormone inhibits its own secretion by acting on the hypothalamus through its receptors on neuropeptide Y neurons in the arcuate nucleus and somatostatin neurons in the periventricular nucleus. Endocr. J. 1998, 45, S19–S26. [Google Scholar] [CrossRef] [Green Version]
- Scharf, J.G.; Dombrowski, F.; Ramadori, G. The IGF axis and hepatocarcinogenesis. Mol. Pathol. 2001, 54, 138–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, N.M.; Ragy, M.M.; Ahmed, S.M. Somatostatin analogue, Octreotide, improves restraint stress-induced liver injury by ameliorating oxidative stress, inflammatory response, and activation of hepatic stellate cells. Cell Stress. Chaperones 2018, 23, 1237–1245. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.Y.; Gao, J.H.; Zhao, C.; Wen, S.L.; Tang, C.W.; Wang, Y.F. Cyclooxygenase-2 up-regulates hepatic somatostatin receptor 2 expression. Sci. Rep. 2018, 8, 11033. [Google Scholar] [CrossRef] [Green Version]
- El-Sisi, A.E.E.; Sokar, S.S.; Shebl, A.M.; Mohamed, D.Z.; Abu-Risha, S.E. Octreotide and melatonin alleviate inflammasome-induced pyroptosis through inhibition of TLR4-NF-kappaB-NLRP3 pathway in hepatic ischemia/reperfusion injury. Toxicol. Appl. Pharmacol. 2021, 410, 115340. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Tong, H.; Gao, J.H.; Tang, S.H.; Yang, W.J.; Wang, G.M.; Zhou, H.Y.; Wen, S.L. Anti-inflammation treatment for protection of hepatocytes and amelioration of hepatic fibrosis in rats. Exp. Ther. Med. 2021, 22, 1213. [Google Scholar] [CrossRef]
- Griffiths, J.; Mills, M.T.; Ong, A.C. Long-acting somatostatin analogue treatments in autosomal dominant polycystic kidney disease and polycystic liver disease: A systematic review and meta-analysis. BMJ Open 2020, 10, e032620. [Google Scholar] [CrossRef] [Green Version]
- Garofalo, C.; Capuano, I.; Pennino, L.; De Gregorio, I.; Riccio, E.; Provenzano, M.; Crocetto, F.; Buonanno, P.; Pandolfo, S.D.; Andreucci, M.; et al. The effects of somatostatin analogues on liver volume and quality of life in polycystic liver disease: A meta-analysis of randomized controlled trials. Sci. Rep. 2021, 11, 23500. [Google Scholar] [CrossRef]
- Fiebiger, W.C.; Scheithauer, W.; Traub, T.; Kurtaran, A.; Gedlicka, C.; Kornek, G.V.; Virgolini, I.; Raderer, M. Absence of therapeutic efficacy of the somatostatin analogue lanreotide in advanced primary hepatic cholangiocellular cancer and adenocarcinoma of the gallbladder despite in vivo somatostatin-receptor expression. Scand. J. Gastroenterol. 2002, 37, 222–225. [Google Scholar] [CrossRef]
- Periferakis, A.; Tsigas, G.; Periferakis, A.T.; Badarau, I.A.; Scheau, A.E.; Tampa, M.; Georgescu, S.R.; Didilescu, A.C.; Scheau, C.; Caruntu, C. Antitumoral and Anti-inflammatory Roles of Somatostatin and Its Analogs in Hepatocellular Carcinoma. Anal. Cell. Pathol. 2021, 2021, 1840069. [Google Scholar] [CrossRef]
- Huang, C.Z.; Huang, A.M.; Liu, J.F.; Wang, B.; Lin, K.C.; Ye, Y.B. Somatostatin Octapeptide Inhibits Cell Invasion and Metastasis in Hepatocellular Carcinoma Through PEBP1. Cell. Physiol. Biochem. 2018, 47, 2340–2349. [Google Scholar] [CrossRef]
- Raderer, M.; Hejna, M.H.; Kurtaran, A.; Kornek, G.V.; Valencak, J.B.; Oberhuber, G.; Vorbeck, F.; Virgolini, I.; Scheithauer, W. Successful treatment of an advanced hepatocellular carcinoma with the long-acting somatostatin analog lanreotide. Am. J. Gastroenterol. 1999, 94, 278–279. [Google Scholar] [CrossRef] [PubMed]
- Lasfer, M.; Vadrot, N.; Schally, A.V.; Nagy, A.; Halmos, G.; Pessayre, D.; Feldmann, G.; Reyl-Desmars, F.J. Potent induction of apoptosis in human hepatoma cell lines by targeted cytotoxic somatostatin analogue AN-238. J. Hepatol. 2005, 42, 230–237. [Google Scholar] [CrossRef]
- Tan, C.K.; Podila, P.V.; Taylor, J.E.; Nagorney, D.M.; Wiseman, G.A.; Gores, G.J.; LaRusso, N.F. Human cholangiocarcinomas express somatostatin receptors and respond to somatostatin with growth inhibition. Gastroenterology 1995, 108, 1908–1916. [Google Scholar] [CrossRef]
- Ciresi, A.; Guarnotta, V.; Campo, D.; Giordano, C. Hepatic Steatosis Index in Acromegaly: Correlation with Insulin Resistance Regardless of the Disease Control. Int. J. Endocrinol. 2018, 2018, 5421961. [Google Scholar] [CrossRef] [Green Version]
- Brighi, N.; Lamberti, G.; Maggio, I.; Manuzzi, L.; Ricci, C.; Casadei, R.; Santini, D.; Mosconi, C.; Lisotti, A.; Ambrosini, V.; et al. Biliary stone disease in patients receiving somatostatin analogs for neuroendocrine neoplasms. A retrospective observational study. Dig. Liver Dis. 2019, 51, 689–694. [Google Scholar] [CrossRef] [PubMed]
- Visentin, M.; Stieger, B.; Merz, M.; Kullak-Ublick, G.A. Octreotide inhibits the bilirubin carriers organic anion transporting polypeptides 1B1 and 1B3 and the multidrug resistance-associated protein 2. J. Pharmacol. Exp. Ther. 2015, 355, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Ukena, K.; Iwakoshi-Ukena, E.; Taniuchi, S.; Bessho, Y.; Maejima, S.; Masuda, K.; Shikano, K.; Kondo, K.; Furumitsu, M.; Tachibana, T. Identification of a cDNA encoding a novel small secretory protein, neurosecretory protein GL, in the chicken hypothalamic infundibulum. Biochem. Biophys. Res. Commun. 2014, 446, 298–303. [Google Scholar] [CrossRef]
- Ukena, K. Avian and murine neurosecretory protein GL participates in the regulation of feeding and energy metabolism. Gen. Comp. Endocrinol. 2018, 260, 164–170. [Google Scholar] [CrossRef] [PubMed]
- Shikano, K.; Bessho, Y.; Kato, M.; Iwakoshi-Ukena, E.; Taniuchi, S.; Furumitsu, M.; Tachibana, T.; Bentley, G.E.; Kriegsfeld, L.J.; Ukena, K. Localization and function of neurosecretory protein GM, a novel small secretory protein, in the chicken hypothalamus. Sci. Rep. 2018, 8, 704. [Google Scholar] [CrossRef] [Green Version]
- Rupp, S.K.; Wolk, E.; Stengel, A. Nesfatin-1 Receptor: Distribution, Signaling and Increasing Evidence for a G Protein-Coupled Receptor-A Systematic Review. Front. Endocrinol. 2021, 12, 740174. [Google Scholar] [CrossRef]
- Kras, K.; Muszynski, S.; Tomaszewska, E.; Arciszewski, M.B. Minireview: Peripheral Nesfatin-1 in Regulation of the Gut Activity-15 Years since the Discovery. Animals 2022, 12, 101. [Google Scholar] [CrossRef]
- Basar, O.; Akbal, E.; Koklu, S.; Kocak, E.; Tuna, Y.; Ekiz, F.; Gultuna, S.; Yiotalmaz, F.M.; Aydogan, T. A novel appetite peptide, nesfatin-1 in patients with non-alcoholic fatty liver disease. Scand. J. Clin. Lab. Investig. 2012, 72, 479–483. [Google Scholar] [CrossRef] [PubMed]
- Tan, B.K.; Hallschmid, M.; Kern, W.; Lehnert, H.; Randeva, H.S. Decreased cerebrospinal fluid/plasma ratio of the novel satiety molecule, nesfatin-1/NUCB-2, in obese humans: Evidence of nesfatin-1/NUCB-2 resistance and implications for obesity treatment. J. Clin. Endocrinol. Metab. 2011, 96, E669–E673. [Google Scholar] [CrossRef]
- Yin, Y.; Li, Z.; Gao, L.; Li, Y.; Zhao, J.; Zhang, W. AMPK-dependent modulation of hepatic lipid metabolism by nesfatin-1. Mol. Cell. Endocrinol. 2015, 417, 20–26. [Google Scholar] [CrossRef]
- Fan, X.T.; Tian, Z.; Li, S.Z.; Zhai, T.; Liu, J.L.; Wang, R.; Zhang, C.S.; Wang, L.X.; Yuan, J.H.; Zhou, Y.; et al. Ghrelin Receptor Is Required for the Effect of Nesfatin-1 on Glucose Metabolism. Front. Endocrinol. 2018, 9, 633. [Google Scholar] [CrossRef] [PubMed]
- Solmaz, A.; Gulcicek, O.B.; Ercetin, C.; Yigitbas, H.; Yavuz, E.; Arici, S.; Erzik, C.; Zengi, O.; Demirturk, P.; Celik, A.; et al. Nesfatin-1 alleviates extrahepatic cholestatic damage of liver in rats. Bosn. J. Basic. Med. Sci. 2016, 16, 247–253. [Google Scholar] [CrossRef] [Green Version]
- Palma, C. Tachykinins and their receptors in human malignancies. Curr. Drug. Targets 2006, 7, 1043–1052. [Google Scholar] [CrossRef]
- Uyama, N.; Geerts, A.; Reynaert, H. Neural connections between the hypothalamus and the liver. Anat. Rec. A Discov. Mol. Cell. Evol. Biol. 2004, 280, 808–820. [Google Scholar] [CrossRef] [PubMed]
- Glaser, S.; Gaudio, E.; Renzi, A.; Mancinelli, R.; Ueno, Y.; Venter, J.; White, M.; Kopriva, S.; Chiasson, V.; DeMorrow, S.; et al. Knockout of the neurokinin-1 receptor reduces cholangiocyte proliferation in bile duct-ligated mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2011, 301, G297–G305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, Y.; Meng, F.; Wu, N.; Zhou, T.; Venter, J.; Francis, H.; Kennedy, L.; Glaser, T.; Bernuzzi, F.; Invernizzi, P.; et al. Substance P increases liver fibrosis by differential changes in senescence of cholangiocytes and hepatic stellate cells. Hepatology 2017, 66, 528–541. [Google Scholar] [CrossRef] [Green Version]
- Bang, R.; Biburger, M.; Neuhuber, W.L.; Tiegs, G. Neurokinin-1 receptor antagonists protect mice from CD95− and tumor necrosis factor-alpha-mediated apoptotic liver damage. J. Pharmacol. Exp. Ther. 2004, 308, 1174–1180. [Google Scholar] [CrossRef] [Green Version]
- Meng, F.; DeMorrow, S.; Venter, J.; Frampton, G.; Han, Y.; Francis, H.; Standeford, H.; Avila, S.; McDaniel, K.; McMillin, M.; et al. Overexpression of membrane metalloendopeptidase inhibits substance P stimulation of cholangiocarcinoma growth. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 306, G759–G768. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Yan, M.; Zhang, H.; Wang, X. Substance P participates in immune-mediated hepatic injury induced by concanavalin A in mice and stimulates cytokine synthesis in Kupffer cells. Exp. Ther. Med. 2013, 6, 459–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bracq, S.; Clement, B.; Pidoux, E.; Moukhtar, M.S.; Jullienne, A. CGRP is expressed in primary cultures of human hepatocytes and in normal liver. FEBS Lett. 1994, 351, 63–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berthoud, H.R. Anatomy and function of sensory hepatic nerves. Anat. Rec. A Discov. Mol. Cell. Evol. Biol. 2004, 280, 827–835. [Google Scholar] [CrossRef]
- Bendtsen, F.; Schifter, S.; Henriksen, J.H. Increased circulating calcitonin gene-related peptide (CGRP) in cirrhosis. J. Hepatol. 1991, 12, 118–123. [Google Scholar] [CrossRef]
- Glaser, S.S.; Ueno, Y.; DeMorrow, S.; Chiasson, V.L.; Katki, K.A.; Venter, J.; Francis, H.L.; Dickerson, I.M.; DiPette, D.J.; Supowit, S.C.; et al. Knockout of alpha-calcitonin gene-related peptide reduces cholangiocyte proliferation in bile duct ligated mice. Lab. Investig. 2007, 87, 914–926. [Google Scholar] [CrossRef] [Green Version]
- Wan, Y.; Ceci, L.; Wu, N.; Zhou, T.; Chen, L.; Venter, J.; Francis, H.; Bernuzzi, F.; Invernizzi, P.; Kyritsi, K.; et al. Knockout of alpha-calcitonin gene-related peptide attenuates cholestatic liver injury by differentially regulating cellular senescence of hepatic stellate cells and cholangiocytes. Lab. Investig. 2019, 99, 764–776. [Google Scholar] [CrossRef]
- Fernandez-Varo, G.; Oro, D.; Cable, E.E.; Reichenbach, V.; Carvajal, S.; de la Presa, B.G.; Wisniewski, K.; Gines, P.; Harris, G.; Jimenez, W. Vasopressin 1a receptor partial agonism increases sodium excretion and reduces portal hypertension and ascites in cirrhotic rats. Hepatology 2016, 63, 207–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fahrner, R.; Patsenker, E.; de Gottardi, A.; Stickel, F.; Montani, M.; Stroka, D.; Candinas, D.; Beldi, G. Elevated liver regeneration in response to pharmacological reduction of elevated portal venous pressure by terlipressin after partial hepatectomy. Transplantation 2014, 97, 892–900. [Google Scholar] [CrossRef] [Green Version]
- Munoz-Ortega, M.; Macias-Segura, N.; Ventura-Juarez, J.; Avila-Blanco, M.E.; Ponce-Damian, L.D.; Gonzalez-Blas, D.; Sanchez-Aleman, E.; Quintanar-Stephano, A. Recovery from Liver Failure and Fibrosis in a Rat Portacaval Anastomosis Model after Neurointermediate Pituitary Lobectomy. J. Immunol. Res. 2021, 2021, 5529784. [Google Scholar] [CrossRef]
- Feher, E.; Pongor, E.; Altdorfer, K.; Kobori, L.; Lengyel, G. Neuroimmunomodulation in human autoimmune liver disease. Cell Tissue Res. 2013, 354, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, F.; Li, Y.; Wang, X.; Lu, Q.; Wang, D.; Qi, C.; Li, C.; Li, Z.; Lian, B.; et al. Combined anti-hepatocellular carcinoma therapy inhibit drug-resistance and metastasis via targeting “substance P-hepatic stellate cells-hepatocellular carcinoma” axis. Biomaterials 2021, 276, 121003. [Google Scholar] [CrossRef]
- Kim, S.; Hong, H.S. Substance-P prevents the cholestatic liver injury by regulating inflammatory responses. Peptides 2021, 137, 170494. [Google Scholar] [CrossRef] [PubMed]
- Hessheimer, A.J.; Martinez de la Maza, L.; Adel Al Shwely, F.; Espinoza, A.S.; Ausania, F.; Fondevila, C. Somatostatin and the “Small-For-Size” Liver. Int. J. Mol. Sci. 2019, 20, 2512. [Google Scholar] [CrossRef] [Green Version]
- Xue, Z.; Zhang, Y.; Liu, Y.; Zhang, C.; Shen, X.D.; Gao, F.; Busuttil, R.W.; Zheng, S.; Kupiec-Weglinski, J.W.; Ji, H. PACAP neuropeptide promotes Hepatocellular Protection via CREB-KLF4 dependent autophagy in mouse liver Ischemia Reperfusion Injury. Theranostics 2020, 10, 4453–4465. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, T.; Zhang, C.; Xue, Z.; Xu, J.; Busuttil, R.W.; Xia, Q.; Xu, N.; Kupiec-Weglinski, J.W.; Ji, H. Pituitary Adenylate Cyclase-activating Polypeptides Prevent Hepatocyte Damage by Promoting Yes-associated Protein in Liver Ischemia-Reperfusion Injury. Transplantation 2019, 103, 1639–1648. [Google Scholar] [CrossRef]
- Sato, A.; Kakinuma, S.; Miyoshi, M.; Kamiya, A.; Tsunoda, T.; Kaneko, S.; Tsuchiya, J.; Shimizu, T.; Takeichi, E.; Nitta, S.; et al. Vasoactive Intestinal Peptide Derived From Liver Mesenchymal Cells Mediates Tight Junction Assembly in Mouse Intrahepatic Bile Ducts. Hepatol. Commun. 2020, 4, 235–254. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Feng, F.; Liu, X.; Sun, C.; Yang, X.; Fang, Y.; Li, S. Exogenous Secretin Improves Parenteral Nutrition-associated Liver Disease in Rats. J. Pediatr. Gastroenterol. Nutr. 2020, 70, 430–435. [Google Scholar] [CrossRef]
- Baiocchi, L.; Lenci, I.; Milana, M.; Kennedy, L.; Sato, K.; Zhang, W.; Ekser, B.; Ceci, L.; Meadows, V.; Glaser, S.; et al. Cyclic AMP Signaling in Biliary Proliferation: A Possible Target for Cholangiocarcinoma Treatment? Cells 2021, 10, 1692. [Google Scholar] [CrossRef]
- Muraki, K.; Nishi, Y.; Arai, M.; Kubo, N.; Ueda, K.; Shikata, H.; Nakata, Y.; Segawa, T.; Yanaihara, N.; Yajima, H. Neurotensin receptors on the rat liver plasma membranes. Biochem. Biophys. Res. Commun. 1987, 145, 1071–1079. [Google Scholar] [CrossRef]
- Mitra, S.P.; Carraway, R.E. High affinity binding of 125I-neurotensin to dispersed cells from chicken liver and brain. Peptides 1997, 18, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.P.; Mazella, J.; Kitabgi, P. Neurotensin and neurotensin receptors. Trends Pharmacol. Sci. 1999, 20, 302–309. [Google Scholar] [CrossRef]
- Nikolaou, S.; Qiu, S.; Fiorentino, F.; Simillis, C.; Rasheed, S.; Tekkis, P.; Kontovounisios, C. The role of Neurotensin and its receptors in non-gastrointestinal cancers: A review. Cell Commun. Signal. 2020, 18, 68. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, M.; Nakao, N.; Yamamoto, I.; Tsushima, N.; Ohta, Y. Changes in expression levels of neurotensin precursor and receptor mRNA in chicken intestinal tissues and liver during late embryonic and early posthatching development. Poult. Sci. 2013, 92, 2765–2771. [Google Scholar] [CrossRef] [PubMed]
- Evers, B.M.; Rajaraman, S.; Chung, D.H.; Townsend, C.M.J.; Wang, X.; Graves, K.; Thompson, J.C. Developmental expression of the neurotensin gene in the rat liver. Ann. Surg. 1993, 218, 183–188. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, K.; Carr, B.I. Neurotensin-amplification of DNA synthesis stimulated by EGF or TGF alpha in primary cultures of adult rat hepatocytes. Cell Struct. Funct. 1993, 18, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Nakaizumi, A.; Uehara, H.; Baba, M.; Iishi, H.; Tatsuta, M. Enhancement by neurotensin of hepatocarcinogenesis by N-nitrosomorpholine in Sprague-Dawley rats. Cancer Lett. 1996, 110, 57–61. [Google Scholar] [CrossRef]
- Xiao, P.; Long, X.; Zhang, L.; Ye, Y.; Guo, J.; Liu, P.; Zhang, R.; Ning, J.; Yu, W.; Wei, F.; et al. Neurotensin/IL-8 pathway orchestrates local inflammatory response and tumor invasion by inducing M2 polarization of Tumor-Associated macrophages and epithelial-mesenchymal transition of hepatocellular carcinoma cells. Oncoimmunology 2018, 7, e1440166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Song, J.; Yan, B.; Weiss, H.L.; Weiss, L.T.; Gao, T.; Evers, B.M. Neurotensin differentially regulates bile acid metabolism and intestinal FXR-bile acid transporter axis in response to nutrient abundance. FASEB J. 2021, 35, e21371. [Google Scholar] [CrossRef] [PubMed]
- Assimakopoulos, S.F.; Vagianos, C.E.; Zervoudakis, G.; Filos, K.S.; Georgiou, C.; Nikolopoulou, V.; Scopa, C.D. Gut regulatory peptides bombesin and neurotensin reduce hepatic oxidative stress and histological alterations in bile duct ligated rats. Regul. Pept. 2004, 120, 185–193. [Google Scholar] [CrossRef]
- Assimakopoulos, S.F.; Tsamandas, A.C.; Georgiou, C.D.; Vagianos, C.E.; Scopa, C.D. Bombesin and neurotensin exert antiproliferative effects on oval cells and augment the regenerative response of the cholestatic rat liver. Peptides 2010, 31, 2294–2303. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Yuan, W.; Liu, T.; Huang, D.; Xiang, L. Bioinformatics analysis of hepatic gene expression profiles in type 2 diabetes mellitus. Exp. Ther. Med. 2019, 18, 4303–4312. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Song, J.; Zaytseva, Y.Y.; Liu, Y.; Rychahou, P.; Jiang, K.; Starr, M.E.; Kim, J.T.; Harris, J.W.; Yiannikouris, F.B.; et al. An obligatory role for neurotensin in high-fat-diet-induced obesity. Nature 2016, 533, 411–415. [Google Scholar] [CrossRef] [Green Version]
- Dongiovanni, P.; Meroni, M.; Petta, S.; Longo, M.; Alisi, A.; Soardo, G.; Valenti, L.; Miele, L.; Grimaudo, S.; Pennisi, G.; et al. Neurotensin up-regulation is associated with advanced fibrosis and hepatocellular carcinoma in patients with MAFLD. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2020, 1865, 158765. [Google Scholar] [CrossRef]
- Villar, B.; Bertran, L.; Aguilar, C.; Binetti, J.; Martinez, S.; Sabench, F.; Real, M.; Riesco, D.; Paris, M.; Del Castillo, D.; et al. Circulating Levels of Pro-Neurotensin and Its Relationship with Nonalcoholic Steatohepatitis and Hepatic Lipid Metabolism. Metabolites 2021, 11, 373. [Google Scholar] [CrossRef]
- Auguet, T.; Aragones, G.; Berlanga, A.; Martinez, S.; Sabench, F.; Aguilar, C.; Villar, B.; Sirvent, J.J.; Del Castillo, D.; Richart, C. Low Circulating Levels of Neurotensin in Women with Nonalcoholic Fatty Liver Disease Associated with Severe Obesity. Obesity 2018, 26, 274–278. [Google Scholar] [CrossRef] [Green Version]
- Mohamed, A.A.; Abo-Elmatty, D.M.; Ezzat, O.; Mesbah, N.M.; Ali, N.S.; Abd El Fatah, A.S.; Alsayed, E.; Hamada, M.; Hassnine, A.A.; Abd-Elsalam, S.; et al. Pro-Neurotensin as a Potential Novel Diagnostic Biomarker for Detection of Nonalcoholic Fatty Liver Disease. Diabetes Metab. Syndr. Obes. 2022, 15, 1935–1943. [Google Scholar] [CrossRef]
- Vaudry, D.; Gonzalez, B.J.; Basille, M.; Yon, L.; Fournier, A.; Vaudry, H. Pituitary adenylate cyclase-activating polypeptide and its receptors: From structure to functions. Pharmacol. Rev. 2000, 52, 269–324. [Google Scholar] [PubMed]
- Cheng, C.Y.; Chu, J.Y.; Chow, B.K. Central and peripheral administration of secretin inhibits food intake in mice through the activation of the melanocortin system. Neuropsychopharmacology 2011, 36, 459–471. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uriz, M.; Saez, E.; Prieto, J.; Medina, J.F.; Banales, J.M. Ursodeoxycholic acid is conjugated with taurine to promote secretin-stimulated biliary hydrocholeresis in the normal rat. PLoS ONE 2011, 6, e28717. [Google Scholar] [CrossRef] [Green Version]
- Banales, J.M.; Saez, E.; Uriz, M.; Sarvide, S.; Urribarri, A.D.; Splinter, P.; Tietz Bogert, P.S.; Bujanda, L.; Prieto, J.; Medina, J.F.; et al. Up-regulation of microRNA 506 leads to decreased Cl−/HCO3− anion exchanger 2 expression in biliary epithelium of patients with primary biliary cirrhosis. Hepatology 2012, 56, 687–697. [Google Scholar] [CrossRef] [Green Version]
- Farouk, M.; Vigna, S.R.; Haebig, J.E.; Gettys, T.W.; McVey, D.C.; Chari, R.; Pruthi, R.S.; Meyers, W.C. Secretin receptors in a new preparation of plasma membranes from intrahepatic biliary epithelium. J. Surg. Res. 1993, 54, 1–6. [Google Scholar] [CrossRef]
- Guerrier, M.; Attili, F.; Alpini, G.; Glaser, S. Prolonged administration of secretin to normal rats increases biliary proliferation and secretin-induced ductal secretory activity. Hepatobiliary Surg. Nutr. 2014, 3, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Wu, N.; Meng, F.; Venter, J.; Giang, T.K.; Francis, H.; Kyritsi, K.; Wu, C.; Franchitto, A.; Alvaro, D.; et al. Knockout of secretin receptor reduces biliary damage and liver fibrosis in Mdr2(−/−) mice by diminishing senescence of cholangiocytes. Lab. Investig. 2018, 98, 1449–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, L.; Francis, H.; Invernizzi, P.; Venter, J.; Wu, N.; Carbone, M.; Gershwin, M.E.; Bernuzzi, F.; Franchitto, A.; Alvaro, D.; et al. Secretin/secretin receptor signaling mediates biliary damage and liver fibrosis in early-stage primary biliary cholangitis. FASEB J. 2019, 33, 10269–10279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feliu, J.E.; Mojena, M.; Silvestre, R.A.; Monge, L.; Marco, J. Stimulatory effect of vasoactive intestinal peptide on glycogenolysis and gluconeogenesis in isolated rat hepatocytes: Antagonism by insulin. Endocrinology 1983, 112, 2120–2127. [Google Scholar] [CrossRef] [PubMed]
- Robberecht, P.; Coy, D.H.; De Neef, P.; Camus, J.C.; Waelbroeck, M.; Christophe, J. Specific labelling of high-affinity vasoactive intestinal peptide receptors in rat liver membranes by a growth hormone-releasing factor analog. Neuroendocrinology 1986, 44, 108–111. [Google Scholar] [CrossRef]
- Nguyen, T.D.; Williams, J.A.; Gray, G.M. Vasoactive intestinal peptide receptor on liver plasma membranes: Characterization as a glycoprotein. Biochemistry 1986, 25, 361–368. [Google Scholar] [CrossRef]
- Gagnon, A.W.; Aiyar, N.; Elshourbagy, N.A. Molecular cloning and functional characterization of a human liver vasoactive intestinal peptide receptor. Cell Signal. 1994, 6, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Knodell, R.G.; Steele, N.M.; Stanley, L.N. Hepatic bile formation in the rat. Addition of vasoactive intestinal peptide to the equation. Dig. Dis. Sci. 1987, 32, 1290–1296. [Google Scholar] [CrossRef]
- Nyberg, B.; Einarsson, K.; Sonnenfeld, T. Evidence that vasoactive intestinal peptide induces ductular secretion of bile in humans. Gastroenterology 1989, 96, 920–924. [Google Scholar] [CrossRef]
- Rodriguez-Henche, N.; Guijarro, L.G.; Bajo, A.M.; Arilla, E.; Prieto, J.C. VIP receptor/effector system in liver membranes from cholestatic rats. Peptides 1994, 15, 353–357. [Google Scholar] [CrossRef]
- Rodriguez-Henche, N.; Guijarro, L.G.; Couvineau, A.; Carrero, I.; Arilla, E.; Laburthe, M.; Prieto, J.C. G proteins in rat liver proliferation during cholestasis. Hepatology 1994, 20, 1041–1047. [Google Scholar] [CrossRef] [PubMed]
- Cho, W.K.; Boyer, J.L. Vasoactive intestinal polypeptide is a potent regulator of bile secretion from rat cholangiocytes. Gastroenterology 1999, 117, 420–428. [Google Scholar] [CrossRef]
- Absood, A.; Hu, B.; Bassily, N.; Colletti, L. VIP inhibits human HepG2 cell proliferation in vitro. Regul. Pept. 2008, 146, 285–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, M.; Takeba, Y.; Iiri, T.; Ohta, Y.; Ootaki, M.; Watanabe, M.; Watanabe, D.; Koizumi, S.; Otsubo, T.; Matsumoto, N. Vasoactive intestinal peptide increases apoptosis of hepatocellular carcinoma by inhibiting the cAMP/Bcl-xL pathway. Cancer Sci. 2019, 110, 235–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Q.; Wang, Y.; Feng, D.; Xu, Y.; Xu, L. Vasoactive intestinal peptide attenuates concanavalin A-mediated liver injury. Eur. J. Pharmacol. 2009, 607, 226–233. [Google Scholar] [CrossRef]
- Jiang, W.; Tang, W.; Geng, Q.; Xu, X. Inhibition of Toll-like receptor 4 with vasoactive intestinal peptide attenuates liver ischemia-reperfusion injury. Transplant. Proc. 2011, 43, 1462–1467. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Zhang, Y.; Liu, Y.; Shen, X.D.; Gao, F.; Nguyen, T.T.; Busuttil, R.W.; Waschek, J.A.; Kupiec-Weglinski, J.W. Vasoactive intestinal peptide attenuates liver ischemia/reperfusion injury in mice via the cyclic adenosine monophosphate-protein kinase a pathway. Liver Transpl. 2013, 19, 945–956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottschall, P.E.; Tatsuno, I.; Miyata, A.; Arimura, A. Characterization and distribution of binding sites for the hypothalamic peptide, pituitary adenylate cyclase-activating polypeptide. Endocrinology 1990, 127, 272–277. [Google Scholar] [CrossRef] [PubMed]
- Robberecht, P.; Gourlet, P.; Cauvin, A.; Buscail, L.; De Neef, P.; Arimura, A.; Christophe, J. PACAP and VIP receptors in rat liver membranes. Am. J. Physiol. 1991, 260, G97–G102. [Google Scholar] [CrossRef] [PubMed]
- Arimura, A. Receptors for pituitary adenylate cyclase-activating polypeptide: Comparison with vasoactive intestinal peptide receptors. Trends Endocrinol. Metab. 1992, 3, 288–294. [Google Scholar] [CrossRef]
- Hosoya, M.; Onda, H.; Ogi, K.; Masuda, Y.; Miyamoto, Y.; Ohtaki, T.; Okazaki, H.; Arimura, A.; Fujino, M. Molecular cloning and functional expression of rat cDNAs encoding the receptor for pituitary adenylate cyclase activating polypeptide (PACAP). Biochem. Biophys. Res. Commun. 1993, 194, 133–143. [Google Scholar] [CrossRef]
- Nguyen, T.D.; Heintz, G.G.; Wolfe, M.S. Structural characterization of PACAP receptors on rat liver plasma membranes. Am. J. Physiol. 1993, 265, G811–G818. [Google Scholar] [CrossRef]
- Yokota, C.; Kawai, K.; Ohashi, S.; Watanabe, Y.; Yamashita, K. PACAP stimulates glucose output from the perfused rat liver. Peptides 1995, 16, 55–60. [Google Scholar] [CrossRef]
- Bosch, J.; Abraldes, J.G.; Groszmann, R. Current management of portal hypertension. J. Hepatol. 2003, 38 (Suppl. S1), S54–S68. [Google Scholar] [CrossRef]
- Yu, M.; Das, J.M. Neuroanatomy, Nucleus Supraoptic. In StatPearls; StatPearls: Treasure Island, FL, USA, 2023. [Google Scholar]
- Papaluca, T.; Gow, P. Terlipressin: Current and emerging indications in chronic liver disease. J. Gastroenterol. Hepatol. 2018, 33, 591–598. [Google Scholar] [CrossRef]
- Israelsen, M.; Dahl, E.K.; Madsen, B.S.; Wiese, S.; Bendtsen, F.; Moller, S.; Fialla, A.D.; Jensen, B.L.; Krag, A. Dobutamine reverses the cardio-suppressive effects of terlipressin without improving renal function in cirrhosis and ascites: A randomized controlled trial. Am. J. Physiol. Gastrointest. Liver Physiol. 2020, 318, G313–G321. [Google Scholar] [CrossRef]
- Montero-Melendez, T.; Boesen, T.; Jonassen, T.E.N. Translational advances of melanocortin drugs: Integrating biology, chemistry and genetics. Semin. Immunol. 2022, 59, 101603. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, A.V.; Arab, J.P.; Premkumar, M.; Benitez, C.; Tirumalige Ravikumar, S.; Kumar, P.; Sharma, M.; Reddy, D.N.; Simonetto, D.A.; Rao, P.N. Terlipressin has stood the test of time: Clinical overview in 2020 and future perspectives. Liver Int. 2020, 40, 2888–2905. [Google Scholar] [CrossRef]
- Cavallin, M.; Piano, S.; Romano, A.; Fasolato, S.; Frigo, A.C.; Benetti, G.; Gola, E.; Morando, F.; Stanco, M.; Rosi, S.; et al. Terlipressin given by continuous intravenous infusion versus intravenous boluses in the treatment of hepatorenal syndrome: A randomized controlled study. Hepatology 2016, 63, 983–992. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, F.; Pappas, S.C.; Curry, M.P.; Reddy, K.R.; Rubin, R.A.; Porayko, M.K.; Gonzalez, S.A.; Mumtaz, K.; Lim, N.; Simonetto, D.A.; et al. Terlipressin plus Albumin for the Treatment of Type 1 Hepatorenal Syndrome. N. Engl. J. Med. 2021, 384, 818–828. [Google Scholar] [CrossRef] [PubMed]
- Audsley, J.; Sasadeusz, J.; Lewin, S.R. Tesamorelin, liver fat, and NAFLD in the setting of HIV. Lancet HIV 2019, 6, e808–e809. [Google Scholar] [CrossRef]
- Fourman, L.T.; Billingsley, J.M.; Agyapong, G.; Ho Sui, S.J.; Feldpausch, M.N.; Purdy, J.; Zheng, I.; Pan, C.S.; Corey, K.E.; Torriani, M.; et al. Effects of tesamorelin on hepatic transcriptomic signatures in HIV-associated NAFLD. JCI Insight 2020, 5, e140134. [Google Scholar] [CrossRef]
- Huang, Z.; Lu, X.; Huang, L.; Zhang, C.; Veldhuis, J.D.; Cowley, M.A.; Chen, C. Stimulation of endogenous pulsatile growth hormone secretion by activation of growth hormone secretagogue receptor reduces the fat accumulation and improves the insulin sensitivity in obese mice. FASEB J. 2021, 35, e21269. [Google Scholar] [CrossRef]
- Peng, S.; Zhou, Y.L.; Song, Z.Y.; Lin, S. Effects of Neuropeptide Y on Stem Cells and Their Potential Applications in Disease Therapy. Stem Cells Int. 2017, 2017, 6823917. [Google Scholar] [CrossRef] [Green Version]
- Bu, F.T.; Jia, P.C.; Zhu, Y.; Yang, Y.R.; Meng, H.W.; Bi, Y.H.; Huang, C.; Li, J. Emerging therapeutic potential of adeno-associated virus-mediated gene therapy in liver fibrosis. Mol. Ther. Methods Clin. Dev. 2022, 26, 191–206. [Google Scholar] [CrossRef]
- Amor, C.; Feucht, J.; Leibold, J.; Ho, Y.J.; Zhu, C.; Alonso-Curbelo, D.; Mansilla-Soto, J.; Boyer, J.A.; Li, X.; Giavridis, T.; et al. Senolytic CAR T cells reverse senescence-associated pathologies. Nature 2020, 583, 127–132. [Google Scholar] [CrossRef] [PubMed]


| Neuropeptide/Receptor/Drug | Disease/Therapeutical Application | Assays/Tests/Measurements | Clinical Trial Identifier/Phase/Country | Start Year/End Year |
|---|---|---|---|---|
| Agouti-related protein/melanocortin receptor inhibitor/TTP435 | Obesity Weight loss | Change in body weight, BMI, serum baseline glucose, insulin, free fatty acids, leptin, after 8 weeks | NCT00779519/ Phase 2/ TransTech Pharma/Canada | 2008/2011 |
| Setmelanotide | Hypothalamic obesity | Change in body weight, BMI, changes in waist circumference, change in hunger after 16 weeks | NCT04725240 | 2021–2022 |
| Corticotropin-releasing hormone (CRH)/CRH receptor/Corticorelin | Craving Mood Addiction Substance-related disorders | Stress hormone level (plasma ACTH, cortisol). Subjective measures of stress, craving, mood. | NCT01984177/ Phase 1/Center for Addiction and Mental Health/Canada | 2013/2016 |
| Neurotensin + Glucagon-like peptide 1 (GLP-1) | Obesity | Ad libitum food intake. VAS for hunger. Plasma glucose, NT, GLP-1, insulin, BA, ghrelin, leptin | NCT04186026/ Phase 1/University of Copenhagen/ Denmark | 2019/2024 |
| Neurotensin (NT)/ | Effect on food intake, appetite | Visual analog scales (VAS) for food appetite, plasma glucose, insulin, bile acids, NT, ghrelin | NCT03522792/ Phase 1/ University of Copenhagen/ Denmark | 2017/2019 |
| Somatostatin (SST)/SST receptor/SST | Post hepatectomy liver failure | Complication rate. Time of Recovery. In-hospital mortality. | NCT02882347 Phase 3 Korea | 2015/2017 |
| Somatostatin (SST)/SST receptor/SST | Liver failure. Impaired hepatic circulation in patients with liver resection. | Complication rate. Time of Recovery. In-hospital mortality. | NCT04010669/ Phase 3/Korea University Anam Hospital, Korea | 2015/2017 |
| Octreotide, an SST analog | Locally advanced, metastatic liver cancer | Median survival within 6 months. Toxicities. | NCT00257426/Phase 2/University of North Carolina, USA | 2005–2007 |
| Octreotide | Hepatectomy | Liver recovery rate. Area under the curve of bilirubin. Liver volume by CT scan. | NCT03179995/Phase 2/Fox Chase Cancer Center, PA, USA | 2017–2088 |
| Octreotide | Polycystic Liver Disease | Percent change in liver volume by MRI and CT scan. Mean score of health-related quality of life. | NCT00426153/Phase 3/ Mayo Clinic, USA | 2007–2008 |
| Substance P (SP) or Neurokin-1/NK1-R antagonist/Serlopitant | Pruritus * | Severity of pruritus on a scale of 0–100 | NCT03836001/ Phase 2/ Stanford University, USA | 2019/2022 |
| Substance P (SP) or Neurokin-1/NK1-R antagonist/Aprepitant | Pruritus ** | Severity of pruritus on a scale of 0–100 | NCT01625455/ Phase 4/ Vanderbilt University Medical Center/USA | 2012/2016 |
| Terlipressin (glycine vasopressin) | Cirrhosis with hepatorenal syndrome type 1 (HRS1) | Double-blind study to confirm reversal of HRS1 | NCT01143246/Phase 2 Mallinckrodt/ USA | 2010–2013 |
| Terlipressin | Efficacy and safety in hepatorenal syndrome type 1 HRS), and cirrhosis | HRS reversal based on serum creatin assays | NCT02770716/Phase 3/Mallinckrodt/ USA | 2016–2019 |
| Terlipressin | Hepatectomy | Blood loss. Lactate level in serum. | NCT02588716/Phase 2 Assiut University Hospital, Egypt | 2015–2017 |
| Terlipressin | Hepatobiliary surgery | Portal pressure changes | NCT02718599/Phase2 Assiut University Hospital, Egypt | 2016–2017 |
| Terlipressin versus STT and/or Octreotide (Sandostatin) | Cirrhosis with acute variceal bleeding | Acute kidney incidence. In-hospital mortality. | NCT03846180/ General Hospital of Shenyang MR, China | 2019–2019 |
| Terlipressin plus albumin therapy | Hepatorenal syndrome in acute-on-chronic liver failure | Acute kidney injury reversal. Baseline organ failure(s). Mortality versus time course. | NCT04416282/ Institute of Liver and Biliary Sciences, India | 2020–2022 |
| Terlipressin | Portal vein pressure after liver tumor resection | Change in portal vein pressure. Incidence of hepatic dysfunction and acute renal failure within 8 months. Adverse effects of terlipressin. | NCT03352349/ Shanghai Zhongshan Hospital, China | 2017–2018 |
| Terlipressin with paracentesis albumin | Cirrhosis with recidivation of ascites | Mean number of paracentesis over 6 months. Total ascites retrieval, cirrhosis complications, liver transplantations, and mortality over 6 months. | NCT00986817/Phase 3 Saint Antoine Hospital, France | 2009–2015 |
| Terlipressin plus Octreotide | Cirrhosis and portal hypertension | Change in portal vein pressure | NCT04353193/phase 4 Hospital Clinic of Barcelona, Spain | 2020–2020 |
| Terlipressin versus Octreotide, STT | Cirrhosis with variceal bleeding | Failure to contain bleeding and rebleeding within 5 days of treatment. | NCT00966355/phase 4 Korea University, Korea | 2006–2010 |
| Tesamorelin (analogue of GHRH) | NAFLD in HIV-infected patients | Change in liver mass by MRI, histological quantification of NAFLD severity, serum transaminases | NCT02196831/ Massachusetts General Hospital, USA | 2015–2019 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrescu, A.D.; An, S.Y.; Venter, J.; McMillin, M.; DeMorrow, S. The Role of Hypothalamic Neuropeptides in Regulation of Liver Functions in Health and Disease. Endocrines 2023, 4, 457-487. https://doi.org/10.3390/endocrines4020034
Petrescu AD, An SY, Venter J, McMillin M, DeMorrow S. The Role of Hypothalamic Neuropeptides in Regulation of Liver Functions in Health and Disease. Endocrines. 2023; 4(2):457-487. https://doi.org/10.3390/endocrines4020034
Chicago/Turabian StylePetrescu, Anca D., Su Yeon An, Juliet Venter, Matthew McMillin, and Sharon DeMorrow. 2023. "The Role of Hypothalamic Neuropeptides in Regulation of Liver Functions in Health and Disease" Endocrines 4, no. 2: 457-487. https://doi.org/10.3390/endocrines4020034