
Adaptations in the Hippocampus during the Fetal to Neonatal Transition in Guinea Pigs

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Immunohistochemistry
2.3. Real Time PCR
2.4. Statistical Analyses
3. Results
3.1. Body and Organ Weights
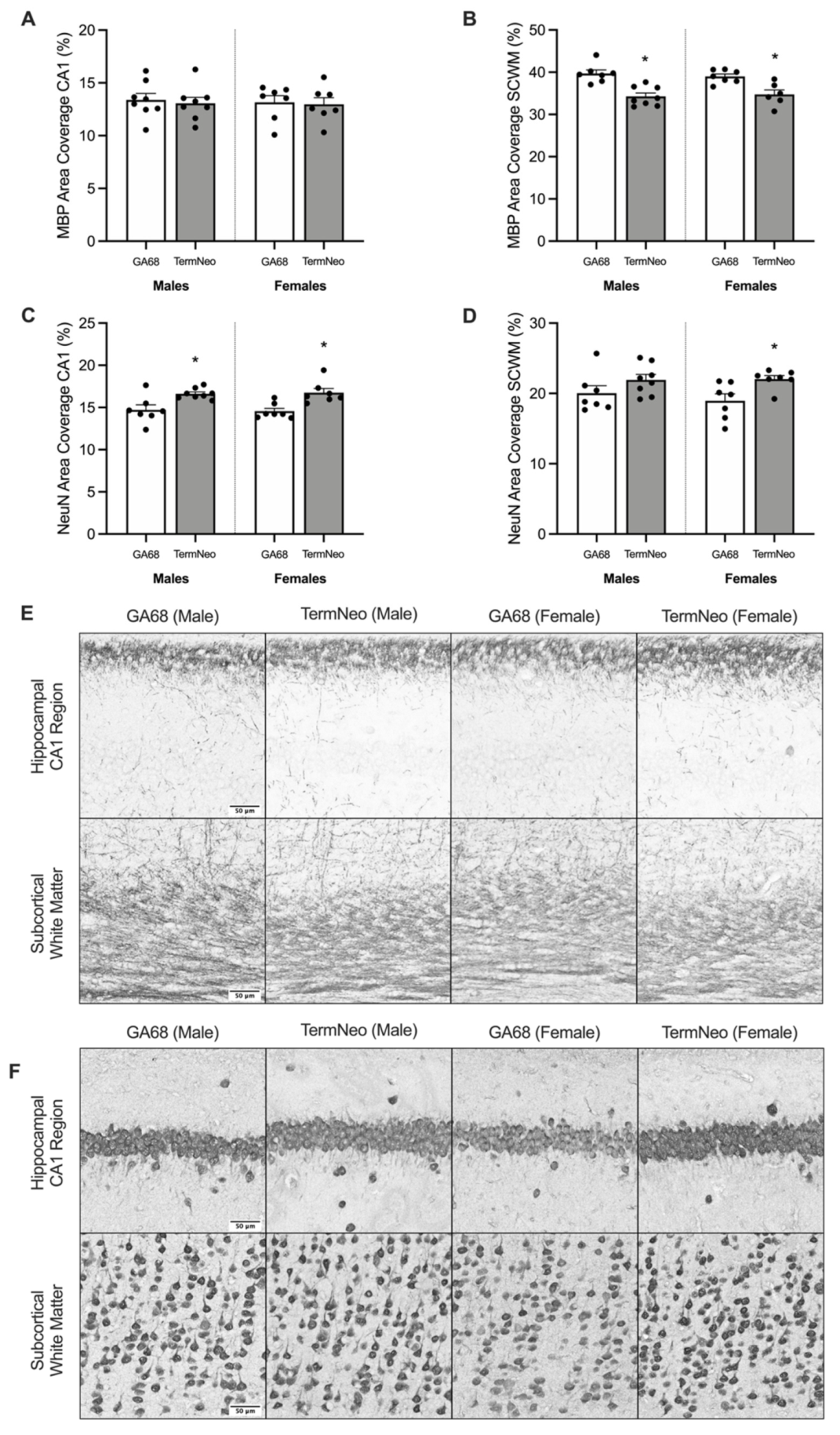
3.2. Myelination and Neuronal Nuclei Expression in the Hippocampus and Subcortical White Matter
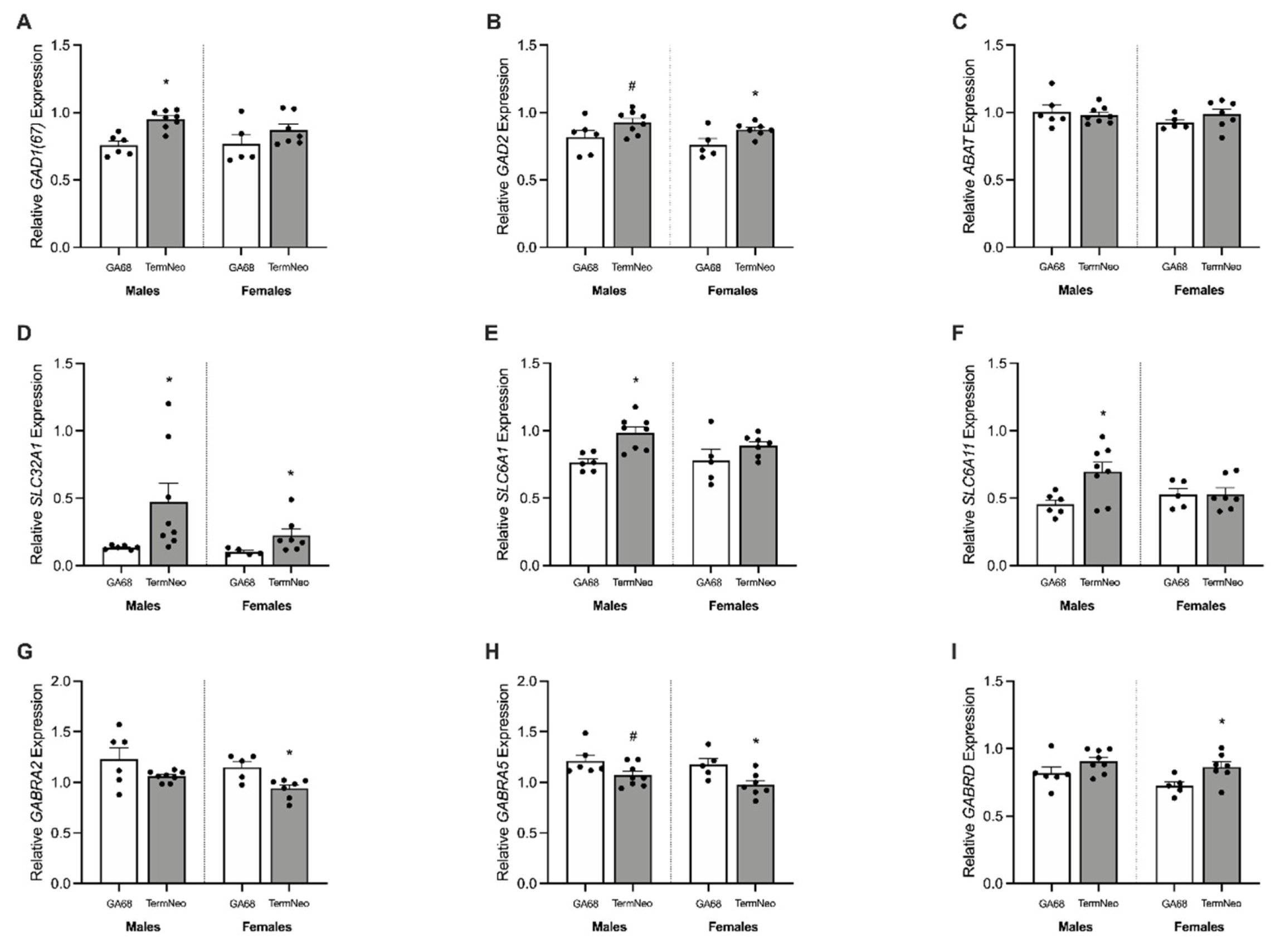
3.3. Relative mRNA Expression of Oligodendrocyte Lineage and Neuron Markers
3.4. Expression of GABAergic Synthetic Enzymes and Transporters
3.5. GABAA Receptor Subunit Expression
3.6. Changes to the Glutamatergic Pathway
3.7. Expression of Steroidogenic Enzymes, Transporters, and Receptors
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Hirst, J.J.; Kelleher, M.A.; Walker, D.W.; Palliser, H.K. Neuroactive steroids in pregnancy: Key regulatory and protective roles in the foetal brain. J. Steroid Biochem. Mol. Biol. 2014, 139, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Hirst, J.J.; Palliser, H.K.; Yates, D.M.; Yawno, T.; Walker, D.W. Neurosteroids in the fetus and neonate: Potential protective role in compromised pregnancies. Neurochem Int. 2008, 52, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Belelli, D.; Lambert, J.J. Neurosteroids: Endogenous regulators of the GABA(A) receptor. Nat. Rev. Neurosci. 2005, 6, 565–575. [Google Scholar] [CrossRef] [PubMed]
- Nicol, M.B.; Hirst, J.J.; Walker, D.W. Effect of finasteride on behavioural arousal and somatosensory evoked potentials in fetal sheep. Neurosci. Lett. 2001, 306, 13–16. [Google Scholar] [CrossRef]
- Yawno, T.; Yan, E.B.; Hirst, J.J.; Walker, D.W. Neuroactive steroids induce changes in fetal sheep behavior during normoxic and asphyxic states. Stress 2011, 14, 13–22. [Google Scholar] [CrossRef]
- Yawno, T.; Miller, S.L.; Bennet, L.; Wong, F.; Hirst, J.J.; Fahey, M.; Walker, D.W. Ganaxolone: A New Treatment for Neonatal Seizures. Front. Cell. Neurosci. 2017, 11, 246. [Google Scholar] [CrossRef] [Green Version]
- Shaw, J.C.; Dyson, R.M.; Palliser, H.K.; Gray, C.; Berry, M.J.; Hirst, J.J. Neurosteroid replacement therapy using the allopregnanolone-analogue ganaxolone following preterm birth in male guinea pigs. Pediatr. Res. 2019, 85, 86–96. [Google Scholar] [CrossRef]
- Yawno, T.; Aridas, J.; Sutherland, A.; Malhotra, A.; Bennet, L.; Jenkin, G.; Walker, D.; Miller, S.; Fahey, M. The effects of ganaxolone in hypoxic ischaemic term lambs. In Proceedings of the 10th Hershey Conference on Developmental Brain Injury, Ecquevilly, France, 8–11 June 2016. [Google Scholar]
- Shaw, J.C.; Palliser, H.K.; Dyson, R.M.; Hirst, J.J.; Berry, M.J. Long-term effects of preterm birth on behavior and neurosteroid sensitivity in the guinea pig. Pediatr Res. 2016, 80, 275–283. [Google Scholar] [CrossRef]
- Shaw, J.C.; Palliser, H.K.; Dyson, R.M.; Berry, M.J.; Hirst, J.J. Disruptions to the cerebellar GABAergic system in juvenile guinea pigs following preterm birth. Int. J. Dev. Neurosci. 2018, 65, 1–10. [Google Scholar] [CrossRef]
- Palliser, H.K.; Kelleher, M.A.; Tolcos, M.; Walker, D.W.; Hirst, J.J. Effect of postnatal progesterone therapy following preterm birth on neurosteroid concentrations and cerebellar myelination in guinea pigs. J. Dev. Orig. Health Dis. 2015, 6, 350–361. [Google Scholar] [CrossRef]
- Crombie, G.K.; Palliser, H.K.; Shaw, J.C.; Hodgson, D.M.; Walker, D.W.; Hirst, J.J. Effects of prenatal stress on behavioural and neurodevelopmental outcomes are altered by maternal separation in the neonatal period. Psychoneuroendocrinology 2021, 124, 105060. [Google Scholar] [CrossRef] [PubMed]
- van Tilborg, E.; de Theije, C.G.M.; van Hal, M.; Wagenaar, N.; de Vries, L.S.; Benders, M.J.; Rowitch, D.H.; Nijboer, C.H. Origin and dynamics of oligodendrocytes in the developing brain: Implications for perinatal white matter injury. Glia 2018, 66, 221–238. [Google Scholar] [CrossRef] [PubMed]
- McLaurin, J.A.; Yong, V.W. Oligodendrocytes and myelin. Neurol. Clin. 1995, 13, 23–49. [Google Scholar] [CrossRef]
- Volpe, J.J.; Kinney, H.C.; Jensen, F.E.; Rosenberg, P.A. The developing oligodendrocyte: Key cellular target in brain injury in the premature infant. Int J. Dev. Neurosci. 2011, 29, 423–440. [Google Scholar] [CrossRef] [Green Version]
- Alexandrou, G.; Martensson, G.; Skiold, B.; Blennow, M.; Aden, U.; Vollmer, B. White matter microstructure is influenced by extremely preterm birth and neonatal respiratory factors. Acta Paediatr. 2014, 103, 48–56. [Google Scholar] [CrossRef]
- Rutherford, M.A.; Supramaniam, V.; Ederies, A.; Chew, A.; Bassi, L.; Groppo, M.; Anjari, M.; Counsell, S.; Ramenghi, L.A. Magnetic resonance imaging of white matter diseases of prematurity. Neuroradiology 2010, 52, 505–521. [Google Scholar] [CrossRef]
- Kelleher, M.; Hirst, J.J.; Palliser, H.K. Changes in neuroactive steroid concentrations after preterm delivery in the guinea pig. Reprod. Sci. 2013, 20, 1365–1375. [Google Scholar] [CrossRef] [Green Version]
- Yawno, T.; Hirst, J.J.; Castillo-Melendez, M.; Walker, D.W. Role of neurosteroids in regulating cell death and proliferation in the late gestation fetal brain. Neuroscience 2009, 163, 838–847. [Google Scholar] [CrossRef]
- Back, S.A. White matter injury in the preterm infant: Pathology and mechanisms. Acta Neuropathol. 2017, 134, 331–349. [Google Scholar] [CrossRef]
- Back, S.A.; Han, B.H.; Luo, N.L.; Chricton, C.A.; Xanthoudakis, S.; Tam, J.; Arvin, K.L.; Holtzman, D.M. Selective vulnerability of late oligodendrocyte progenitors to hypoxia–ischemia. J. Neurosci. 2002, 22, 455–463. [Google Scholar] [CrossRef] [Green Version]
- Morrison, J.L.; Botting, K.J.; Darby, J.R.T.; David, A.L.; Dyson, R.M.; Gatford, K.L.; Gray, C.; Herrera, E.A.; Hirst, J.J.; Kim, B.; et al. Guinea pig models for translation of the developmental origins of health and disease hypothesis into the clinic. J. Physiol. 2018, 596, 5535–5569. [Google Scholar] [CrossRef] [PubMed]
- Berry, M.; Gray, C.; Wright, K.; Dyson, R.; Wright, I. Premature guinea pigs: A new paradigm to investigate the late-effects of preterm birth. J. Dev. Orig. Health Dis. 2015, 6, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Nicol, M.; Hirst, J.; Walker, D. Effect of pregnane steroids on electrocortical activity and somatosensory evoked potentials in fetal sheep. Neurosci. Lett. 1998, 253, 111–114. [Google Scholar] [CrossRef]
- Belelli, D.; Herd, M.B.; Mitchell, E.A.; Peden, D.R.; Vardy, A.W.; Gentet, L.; Lambert, J.J. Neuroactive steroids and inhibitory neurotransmission: Mechanisms of action and physiological relevance. Neuroscience 2006, 138, 821–829. [Google Scholar] [CrossRef] [PubMed]
- Bennett, G.A.; Palliser, H.K.; Shaw, J.C.; Palazzi, K.L.; Walker, D.W.; Hirst, J.J. Maternal stress in pregnancy affects myelination and neurosteroid regulatory pathways in the guinea pig cerebellum. Stress 2017, 20, 580–588. [Google Scholar] [CrossRef] [PubMed]
- Bradl, M.; Lassmann, H. Oligodendrocytes: Biology and pathology. Acta Neuropathol. 2010, 119, 37–53. [Google Scholar] [CrossRef] [Green Version]
- Metwally, E.; Zhao, G.; Zhang, Y.Q. The calcium-dependent protease calpain in neuronal remodeling and neurodegeneration. Trends Neurosci. 2021, 44, 741–752. [Google Scholar] [CrossRef]
- Boulanger, J.J.; Messier, C. Oligodendrocyte progenitor cells are paired with GABA neurons in the mouse dorsal cortex: Unbiased stereological analysis. Neuroscience 2017, 362, 127–140. [Google Scholar] [CrossRef]
- Micheva, K.D.; Chang, E.F.; Nana, A.L.; Seeley, W.W.; Ting, J.T.; Cobbs, C.; Lein, E.; Smith, S.J.; Weinberg, R.J.; Madison, D.V. Distinctive Structural and Molecular Features of Myelinated Inhibitory Axons in Human Neocortex. eNeuro 2018, 5. [Google Scholar] [CrossRef] [Green Version]
- Ben-Ari, Y. Excitatory actions of GABA during development: The nature of the nurture. Nat. Rev. Neurosci. 2002, 3, 728–739. [Google Scholar] [CrossRef]
- Coleman, H.; Hirst, J.J.; Parkington, H.C. The GABAA excitatory-to-inhibitory switch in the hippocampus of perinatal guinea-pigs. In Proceedings of the 40th Annual Meeting Fetal and Neonatal Physiological Society, Peurto Varas, Chile, 1–4 September 2014. [Google Scholar]
- Follett, P.L.; Deng, W.; Dai, W.; Talos, D.M.; Massillon, L.J.; Rosenberg, P.A.; Volpe, J.J.; Jensen, F.E. Glutamate receptor-mediated oligodendrocyte toxicity in periventricular leukomalacia: A protective role for topiramate. J. Neurosci. 2004, 24, 4412–4420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, Y.; Sun, W.; Shi, Y.; Shi, R.; Cheng, J.X. Glutamate excitotoxicity inflicts paranodal myelin splitting and retraction. PLoS ONE 2009, 4, e6705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, J.R.; Mukherjee, B.; Rogers, B.C.; Nafar, F.; Gosse, M.; Parsons, M.P. The Relationship Between Glutamate Dynamics and Activity-Dependent Synaptic Plasticity. J. Neurosci. 2020, 40, 2793–2807. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Protein | Forward Primer | Reverse Primer | Amplicon Size (bp) |
|---|---|---|---|---|
| ABAT | GABA aminotransferase (GABA-T) | GGATGTCCCAGCTTGTCACTA | TTGCTCCGGTACCACATGAA | 85 |
| ACTB | Beta actin (housekeeper) | TGCGTTACACCCTTTCTTGACA | ACAAAGCCATGCCAATCTCAT | 72 |
| AIF1 | Allograft inflammatory factor 1 | GGCTCTGGGGAGACATTCA | AGATCTCTTGCCCAGCATCA | 60 |
| BDNF | Brain derived neurotrophic factor | AATCGGCTGGCGGTTCATAA | AGCCACTATCTGCCCCTCTTA | 75 |
| CALB1 | Calbindin | CTGACTGAGATGGCCAGGTTA | CCCACACATTTTAACTCCCTGAAA | 75 |
| CAPN1 | Calpain1 | TTCCCTCACTCTGAACGACAC | AGTGGAAGATGCCAGCGTAA | 86 |
| CAPN2 | Calpain 2 | TGCAGAAACTGATCCGAGTCC | TCCAGTTTGGGCAGTTGTCA | 81 |
| CAPNS1 | Calpain small subunit 1 | CGACACCCTGACCTGAAAAC | TGTGGTGTCGCTATCCATCA | 81 |
| CAST | Calpastatin | AAGCCAAGGAACCACTTCCA | AGGTATCTGGAGGACTGGAGAA | 84 |
| CKAP5 | Cytoskeleton associated protein 5 | AGGACATGTTGTGCCAACCA | GCCTCTTGCAGGGCTTGTA | 75 |
| CSPG4 | Chondroitin sulfate proteoglycan 4 (NG2) | CTCCTCACCACCACCCTCAA | ACTCTTCAGCACAGCCCTCA | 79 |
| DLG4 | Postsynaptic density protein 95 (PSD-95) | TATTCCCAGCACCTGGACAA | TCATGGCTGTGGGGTAATCA | 70 |
| DNMT1 | DNA-methyltransferase 1 | ACGACGTGGACCTCAAGTAC | CGAAGATGGACAGTGCCTCA | 87 |
| DNMT3A | DNA-methyltransferase 3A | GGGGACAAGAATGCCACCAA | CCAGCTCCCCAATGCCAAA | 79 |
| ENO2 | Neuron specific enolase | TGACCTGACAGTGACCAACC | TTGACCTTGAGCAGCAGACA | 81 |
| FYN | Protein-tyrosine kinase | ACCGGTTACATTCCCAGCAA | GCGGCCAAGTTTTCCAAAGTA | 81 |
| GABRA1 | GABAA receptor subunit alpha 1 | CTCAAGCCCGCAATGAAGAAA | TCCAGTCAACGTGCTCAGAA | 81 |
| GABRA2 | GABAA receptor subunit alpha 2 | ACTAGGCCAATCAATTGGGAA | TCAAGTGGAAATGAGCTGTCA | 80 |
| GABRA3 | GABAA receptor subunit alpha 3 | TTGGCAGCTATGCCTACACA | ACCTCCACAGACTTGTTCTTCC | 73 |
| GABRA4 | GABAA receptor subunit alpha 4 | TGGACAAAGGGTCCTGAGAAA | CACTGTTTGCCCAATCAGATCA | 84 |
| GABRA5 | GABAA receptor subunit alpha 5 | TGGTTCATCGCTGTGTGCTA | CCCAGCCTCTCTTCGTGAAATA | 85 |
| GABRD | GABAA receptor subunit delta | ATGCTGGACCTGGAAAGCTA | GGATCTGCTCCTGGTTCTCA | 76 |
| GABRG2 | GABAA receptor subunit gamma 2 | AGGCAGATGCCCATTGGATA | TGTAGAGCACTCTGCCATCA | 72 |
| GAD1 (67) | Glutamate decarboxlase 1 (67 kDa) | AGCTCGCTACAAGTACTTCCC | TGTGTTCTGAGGTGAAGAGGAC | 83 |
| GAD2 | Glutamate decarboxlase 2 | GGCGCCATCTCCAACATGTA | TGCCCTTCTCCTTGACCTCA | 73 |
| GLS1 | Glutaminase | CACGTTGGTCTTCCTGCAAA | GCACATCATGCCCATGACA | 78 |
| GRIA1 | Glutamate ionotropic receptor AMPA 1 (GluA1) | TGAACGCAGGACTGTCAACA | AAGCTCGGTGTGATGAAGCA | 72 |
| GRIA2 | Glutamate ionotropic receptor AMPA 2 (GluA2) | GACACCTCACATCGACAACC | CGCCTCTTGAAAACTGGGAA | 80 |
| GRIA3 | Glutamate ionotropic receptor AMPA 3 (GluA3) | CATGGCCAGGCAGAAGAAAA | ATGACCCAAAAGCCCCAGTA | 76 |
| GRIA4 | Glutamate ionotropic receptor AMPA 4 (GluA4) | TGTCCTGGTGATGGCTGAAA | ATCCCCAGCATTTCCTCTCC | 79 |
| GRIN1 | Glutamate ionotropic receptor 1 (GluN1) | AGAGCATCCACTTGAGCTTCC | TACACGCGCATCATCTCGAA | 82 |
| GRIN2A | Glutamate ionotropic receptor 2A (GluN2A) | TCGAGGATGCGAAGACACAA | AGCCTCGTCTTTGGAGCAATA | 80 |
| GRIN2B | Glutamate ionotropic receptor 2B (GluN2B) | GCACTAAGGAAGAAGCCACCTA | ATCCATGTGTAGCCGTAGCC | 76 |
| GRIN2C | Glutamate ionotropic receptor 2C (GluN2C) | ATGCACACCCACATGGTCAA | CGTCCAGCTTCCCCATCTTAA | 79 |
| GRIN2D | Glutamate ionotropic receptor 2D (GluN2D) | ACCTGGGATAACCGGGACTA | TGTCTCTGGTGAGGGAAATGAC | 85 |
| GRIN3A | Glutamate ionotropic receptor 3A (GluN3A) | AAGGGTTGCTGCCCTACAA | GCAAAAGTGGCAGGTCTCC | 78 |
| GRM1 | Glutamate metabotropic receptor 1 (mGluR1) | GGGCAGACAGAGATGAAGTCA | CCTCTGGAGACTGCAGCTTTA | 81 |
| GRM2 | Glutamate metabotropic receptor 2 (mGluR2) | GACTTCGTGCTCAACGTCAA | GCCATCTCCAAAACGGTCAA | 87 |
| GRM3 | Glutamate metabotropic receptor 3 (mGluR3) | GCATACCCCAGGTGAGCTAC | TGCGCGCGAAGTAGTCATA | 75 |
| GRM4 | Glutamate metabotropic receptor 4 (mGluR4) | ACGTCAAGAAGTGCACCAAC | AATCACGAACTGCACCTTCC | 80 |
| GRM5 | Glutamate metabotropic receptor 5 (mGluR5) | AAGCTCCGGCCAGAAACAAA | CCTTCCAGCCGACACTGAAA | 80 |
| GRM7 | Glutamate metabotropic receptor 7 (mGluR7) | GATCCTTGCAGCAGCCAAAA | TTTTGGATCCCCAGCTGTCA | 77 |
| GRM8 | Glutamate metabotropic receptor 8 (mGluR8) | ACACTGGCTTCTGAGGGAAA | TGCAAACACCACCAATCTCC | 82 |
| HNRNPA2B1 | Heterogeneous nuclear ribonucleoprotein A2/B1 | TGGACCAGGACCAGGAAGTAA | CCCGTTATAGCCATCCCCAAA | 82 |
| IL1B | Interleukin 1 beta | ACGGCCCCAATAAGATGCA | TTTGCAGCTTGATCCCCTCA | 75 |
| INA | Alpha-internexin | ACAAGATCATCCGCACCAAC | GTGCACCTTTTCGATGAACAC | 80 |
| KIF1B | Kinesin family member 1B | AAGCATGACTCGGAGACCAA | GTTCACTTCCGGCTAGATCCA | 76 |
| MBP | Myelin basic protein | ACCTCCTCCGTCTCAAGGAAA | GCTCTGCCTCCATAGCCAAA | 66 |
| MOG | Myelin oligodendrocyte glycoprotein | GCAGCACAGACTGAGAGGAA | CCCTCAGAAAGTGGGGATCAA | 77 |
| NCAM1 | Neural cell adhesion molecule 1 | TTGTTCCCAGCCAAGGAGAA | TGTCTTTGGCATCTCCTGCTA | 78 |
| NEFH | Neurofilament heavy chain | CTCCGTATCGGCTTCACCAA | CCGTTGCTCAGCGTGTCTA | 81 |
| NEFL | Neurofilament light chain | TGATGCCCAGCTTGGAGAA | CTGCGTGCGGATCGATTTAA | 77 |
| NEFM | Neurofilament medium chain | GAGAGCAGCCTCGACTTCA | GGAGCGGGACAGCTTGTA | 78 |
| NFKB1 | Nuclear factor kappa B subunit 1 | AAGGCTCCCAATGCATCCAA | AATCTCCTCCCCGCCAGTTA | 78 |
| NR3C1 | Glucocorticoid receptor | ACCACAGCCCCAAGTGAAAA | GGCCCAGTTTCTCCTGCTTAA | 83 |
| NR3C2 | Mineralocorticoid receptor | GTGCCATTGTGAAAAGCCCTA | GCTGCAAACCGAAGATGTCA | 86 |
| NTRK2 | Neurotrophic receptor tyrosine kinase 2 | CTCTAATGGGAGCAACACTCCA | GACAGGGATCTTGGTCATCCC | 79 |
| OLIG2 | Oligodendrocyte transcription factor 2 | GCACTCATCCTGGGGACAA | CCGACGACGTGGATGATGAA | 78 |
| PCBP1 | Heterogeneous nuclear ribonucleoprotein E1 | CTGGACGCCTACTCGATTCAA | TTGTCTTGCCACCTGGTTCA | 78 |
| PLP1 | Proteolipid protein 1 | TGCAGCAACATTGGTTTCCC | CTCGGCCCATGAGTTTAAGAAC | 80 |
| PVALB | Parvalbumin | AAGGATGGGGACGGCAAA | GGGTCCATCAGCTCTGCTTA | 77 |
| QKI | KH domain RNA binding | CTGCTGCAATAGTCCCTCCA | AGCTGGTGCCAACGTGTA | 80 |
| RBFOX3 | RNA binding fox-1 homolog 3 (NeuN) | CACAGACAGACAGCCAACCA | CGGAAGGGGATGTTGGAGAC | 88 |
| SLC12A2 | Sodium/potassium/chloride transporter | GGGGAAGAAAGTACTCCAACCA | ACACCCTTGATCCAGCCAAA | 89 |
| SLC12A5 | Potassium-chloride cotransporter 2 | CCTGTATGGCCACTGTGGTA | AGGATGACACAACCCAGGAA | 79 |
| SLC17A6 | Vesicular glutamate transporter 2 (vGlut2) | TGCCATGCCTTTAGCTGGTA | CATGTACCAGACCATCCCAAAAC | 91 |
| SLC17A7 | Vesicular glutamate transporter 1 (vGlut1) | CAGCCTTTTGCGGTTCCTAC | AACAGAGCTCCATCCCGAATAC | 86 |
| SLC17A8 | Vesicular glutamate transporter 3 (vGlut3) | GGATGGGCTTCGGTCTTCTA | GCACTCATAGGCTTGCAACA | 81 |
| SLC1A2 | Excitatory amino acid transporter 2 (EAAT2) | CACAGTCGTCTCCCTGTTGAA | CAGGCCCTTCTTGAGAACCA | 76 |
| SLC1A3 | Excitatory amino acid transporter 1 (EAAT1) | TGACCAAAGGCAATGGGGAA | GCGGACTCCTTGCTGGAA | 68 |
| SLC32A1 | Vesicular inhibitory amino acid transporter (vGAT) | ACACGACAAGCCCAAGATCA | TAGCACGAACATGCCCTGAA | 76 |
| SLC6A1 | GABA transporter 1 (GAT1) | AGCGCTGCTTCTCCAACTAC | ATTGCGCTCCCAAAACTCCA | 77 |
| SLC6A11 | GABA transporter 3 (GAT3) | ATCATGCTCTGCTGCCTGAA | CATAAGCCATGAAGCCCAAGAC | 82 |
| SRD5A1 | Steroid 5 alpha-reductase 1 | GGCTATGTTCGTCGTCCACTA | AAGGCTGACACACAGGTGAA | 99 |
| SRD5A2 | Steroid 5 alpha-reductase 2 | CGGCGAGATCACTGAATGGA | CTCCCGAGGAAACAAAGCGT | 93 |
| SST | Somatostatin | AAGCAGGAACTGGCCAAGTA | TGGGACAAATCTTCAGGTTCCA | 92 |
| STAR | Steroidogenic acute regulatory protein (StAR) | GGACCTTGAGAGGTTCCAGAA | AGCATTGTGCCCAGAAGGTA | 74 |
| SYP | Synaptophysin | TTCAGGCTGCACCAAGTGTA | GTAGTCCCCAACGAGGAAGAC | 78 |
| TBP | TATA-binding protein (housekeeper) | CAAGCGGTTTGCTGCTGTAA | CACCATCTTCCCGGAACTGAA | 79 |
| TSPO | Translocator protein | CCCATCTGGGGTACGCTGTA | AATCCACCCAGCTCCTTCCA | 71 |
| UBE2D2 | Ubiquitin conjugating enzyme E2 D2 (housekeeper) | CAGTGCTGCGTGTTGTACATA | TGCTAGGAGGCAATGTTGGTA | 76 |
| VEGFA | Vascular endothelial growth factor A | GGAGAATGTCCCTCCCAGAA | GCCTCCCTAGAAGGGACAAA | 84 |
| YWHAZ | Tyrosine 3-monooxygenase (housekeeper) | GCTTCACAAGCAGAGAGCAA | CAGCAACTTCGGCCAAGTAA | 76 |
| Sex | Delivery | N | Body Wgt | Brain Wgt | Hippo. Wgt | Cereb. Wgt | Liver Wgt | Heart Wgt | Kidney Wgt | Adren-als Wgt | Sub. Cut. Fat Wgt | Visc. Fat Wgt |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Male | GA68 Fetus | 7 | 102.0 ± 5.6 | 2.68 ± 0.03 | 0.114 ± 0.005 | 0.156 ± 0.010 | 5.32 ± 0.42 | 0.551 ± 0.026 | 0.742 ± 0.050 | 0.034 ± 0.004 | 1.57 ± 0.10 | 0.968 ± 0.066 |
| Term 24 h Neonate | 8 | 97.2 ± 2.8 | 2.61 ± 0.05 | 0.112 ± 0.004 | 0.141 ± 0.005 | 3.97 ± 0.27 * | 0.395 ± 0.018 * | 0.834 ± 0.027 | 0.034 ± 0.002 | 1.40 ± 0.06 | 0.837 ± 0.037 | |
| Female | GA68 Fetus | 7 | 96.4 ± 5.0 | 2.68 ± 0.04 | 0.128 ± 0.014 | 0.174 ± 0.013 | 5.35 ± 0.36 | 0.546 ± 0.047 | 0.725 ± 0.035 | 0.043 ± 0.005 | 1.33 ± 0.09 | 0.863 ± 0.031 |
| Term 24 h Neonate | 7 | 98.6 ± 4.3 | 2.52 ± 0.04 * | 0.123 ± 0.007 | 0.158 ± 0.010 | 3.91 ± 0.29 * | 0.386 ± 0.024 * | 0.849 ± 0.045 * | 0.037 ± 0.005 | 1.22 ± 0.09 | 0.663 ± 0.050 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shaw, J.C.; Dyson, R.M.; Palliser, H.K.; Crombie, G.K.; Berry, M.J.; Hirst, J.J. Adaptations in the Hippocampus during the Fetal to Neonatal Transition in Guinea Pigs. Reprod. Med. 2022, 3, 85-100. https://doi.org/10.3390/reprodmed3020008
Shaw JC, Dyson RM, Palliser HK, Crombie GK, Berry MJ, Hirst JJ. Adaptations in the Hippocampus during the Fetal to Neonatal Transition in Guinea Pigs. Reproductive Medicine. 2022; 3(2):85-100. https://doi.org/10.3390/reprodmed3020008
Chicago/Turabian StyleShaw, Julia C., Rebecca M. Dyson, Hannah K. Palliser, Gabrielle K. Crombie, Mary J. Berry, and Jonathan J. Hirst. 2022. "Adaptations in the Hippocampus during the Fetal to Neonatal Transition in Guinea Pigs" Reproductive Medicine 3, no. 2: 85-100. https://doi.org/10.3390/reprodmed3020008