Characterization of Magnetic Nanoparticles from the Shells of Freshwater Mussel L. fortunei and Marine Mussel P. perna

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
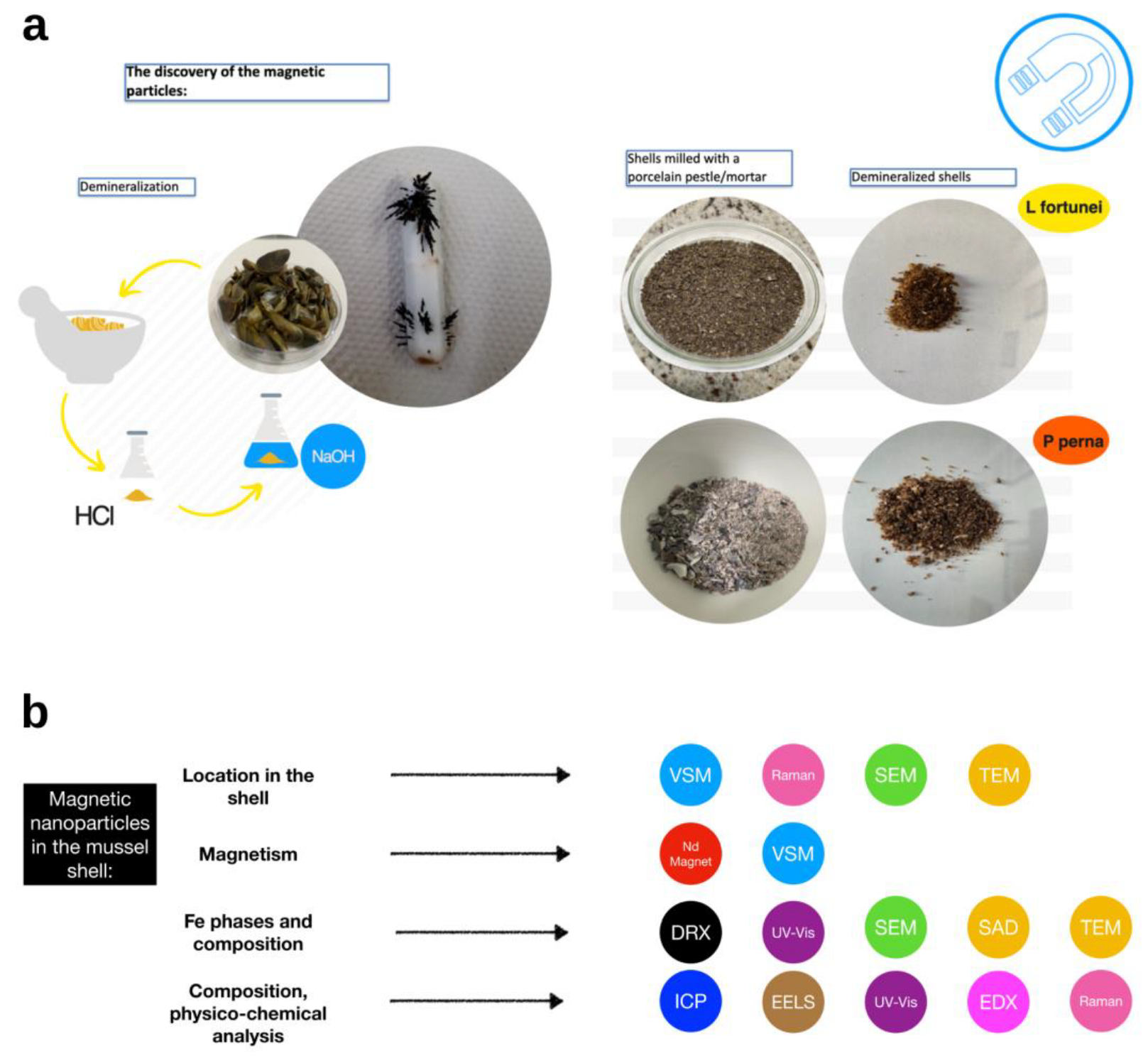
2.1. Specimen Collection and Preparation
2.2. Cleaning, Crushing, Sanding, and Powdering Shell Samples
2.3. Demineralization with Hydrochloric Acid
2.4. Testing Magnetite Particles Dissolved in 1 M HCl Solution
2.5. Demineralization with Ethylenediaminetetraacetic Acid (EDTA)
2.6. Direct Obtention of Magnetic Particles without Demineralization
2.7. Characterization of the Magnetic Powder
3. Results
3.1. Testing Dissolution of Magnetite Particles in 1 M HCl Solution
3.2. Direct Obtention of Magnetic Particles without Demineralization
3.3. Chemical Shell Composition of Limnoperna fortunei Using ICP-OES and EDX
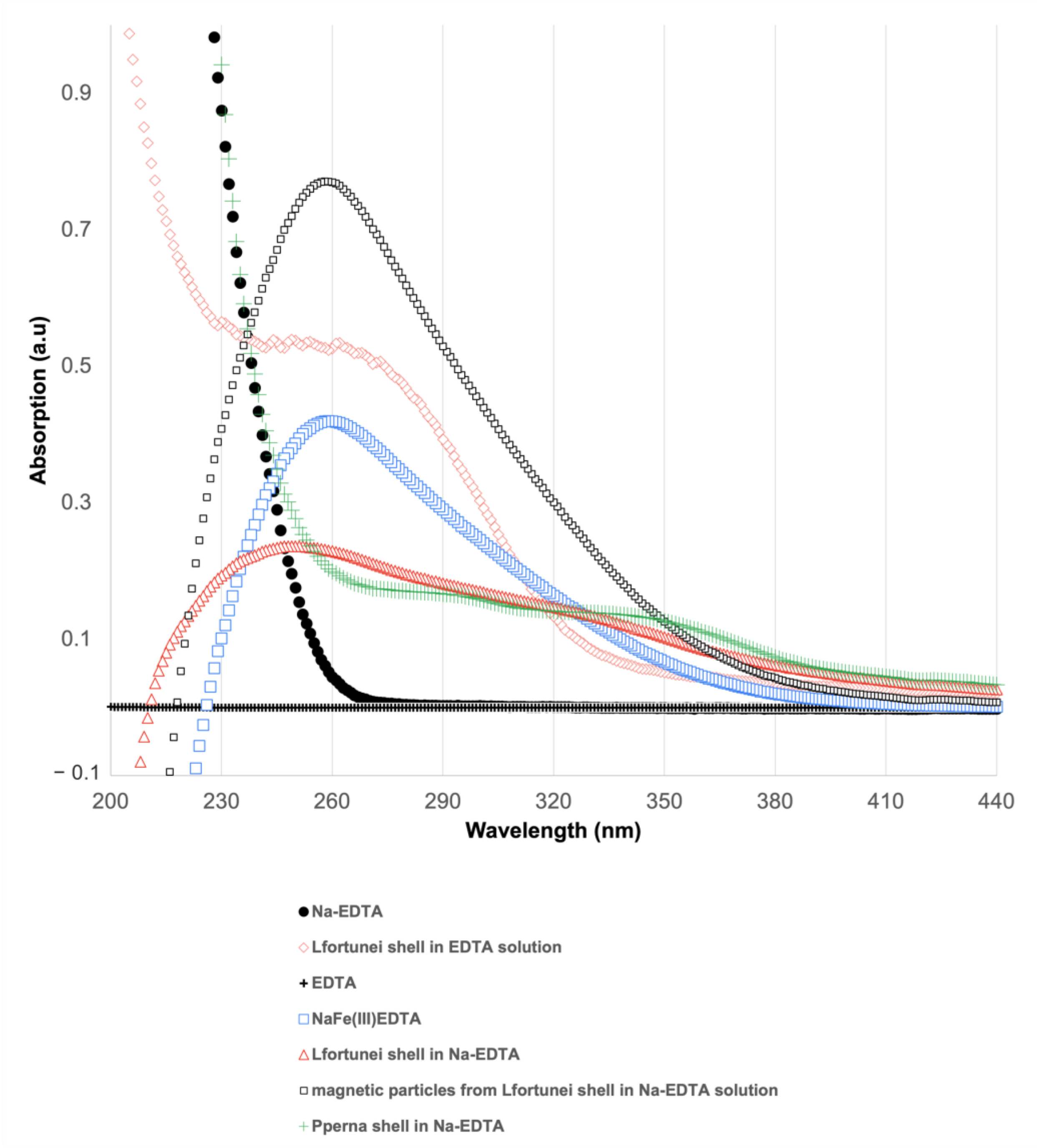
3.4. UV–Vis Spectroscopy
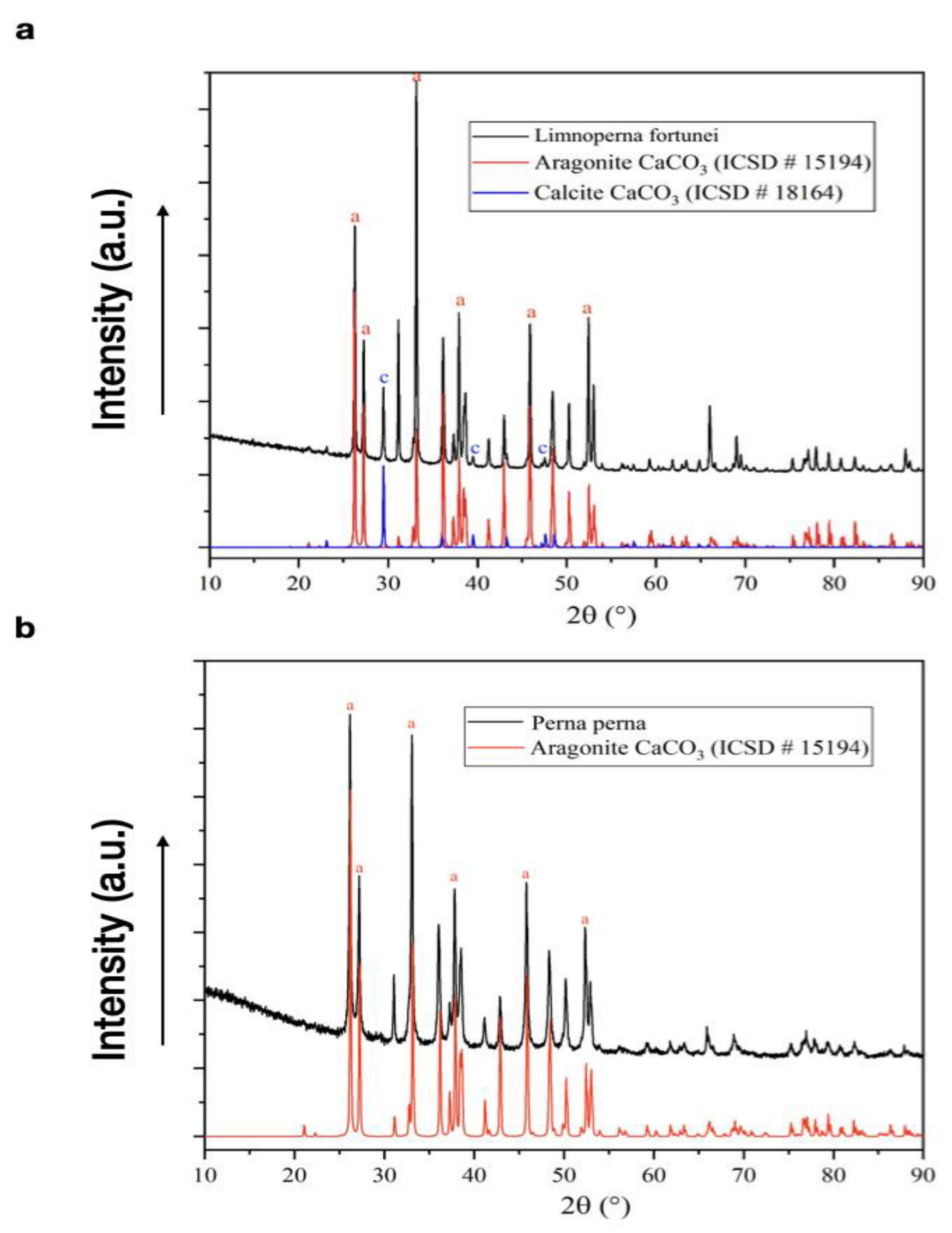
3.5. X-ray Diffraction
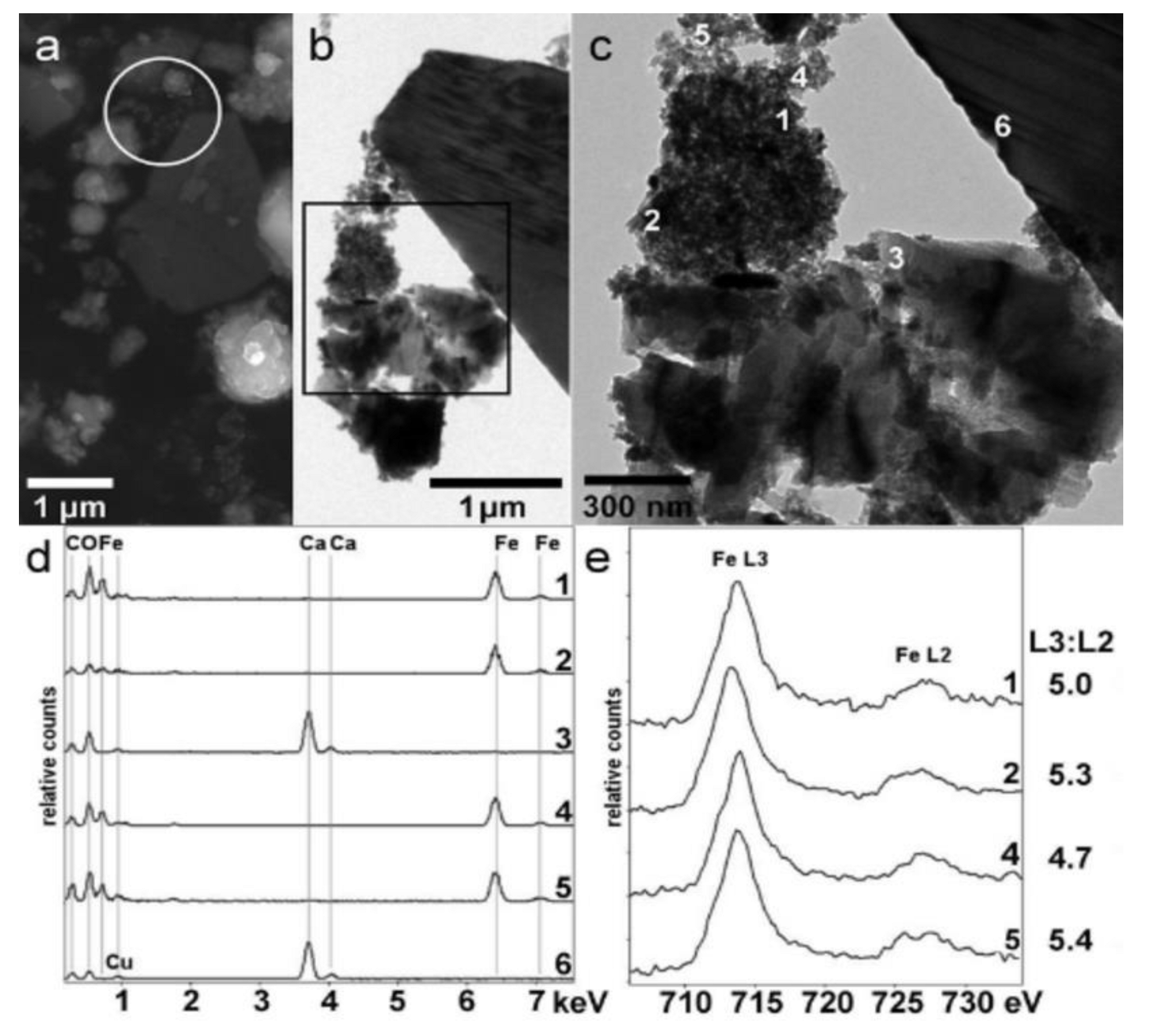
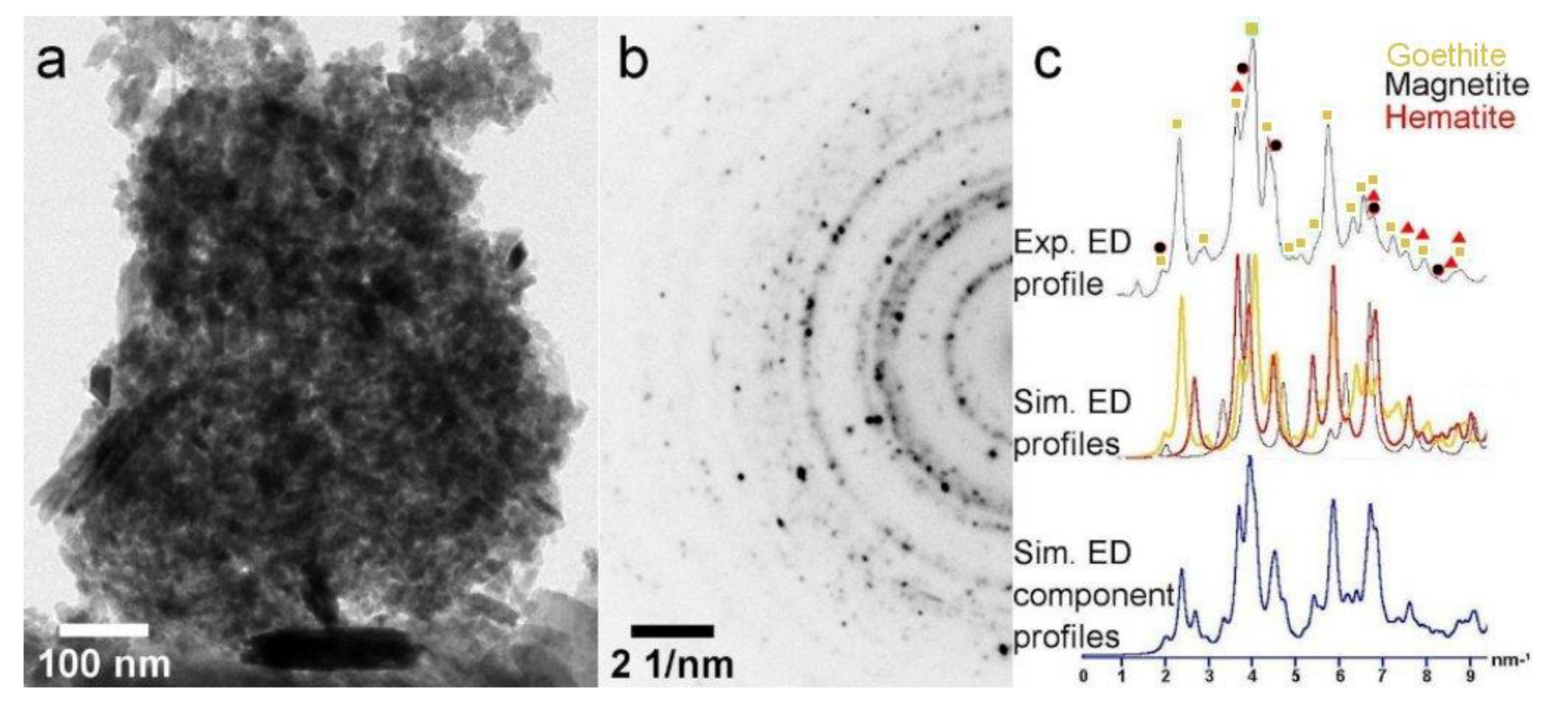
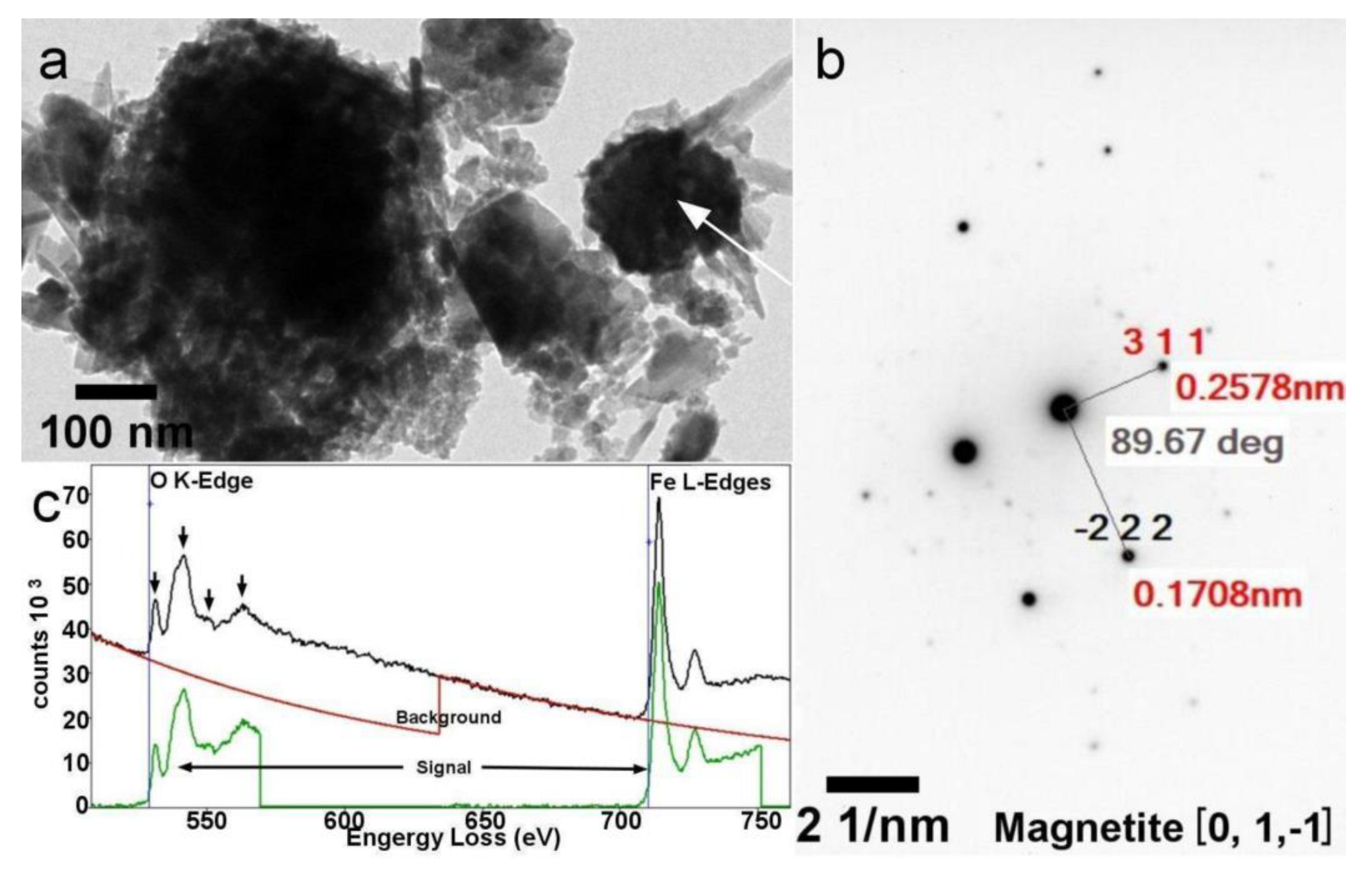
3.6. Electron Microscopy
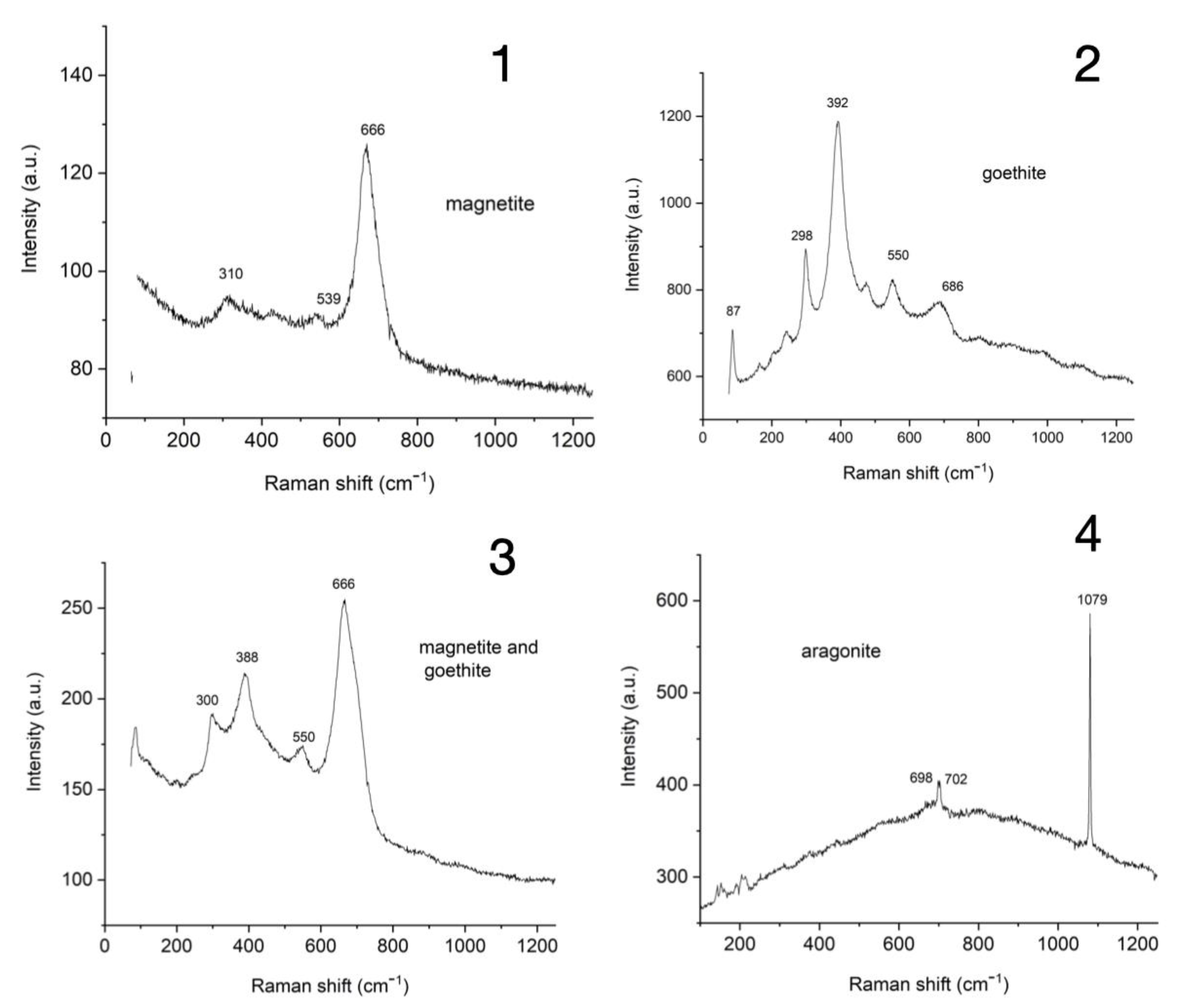
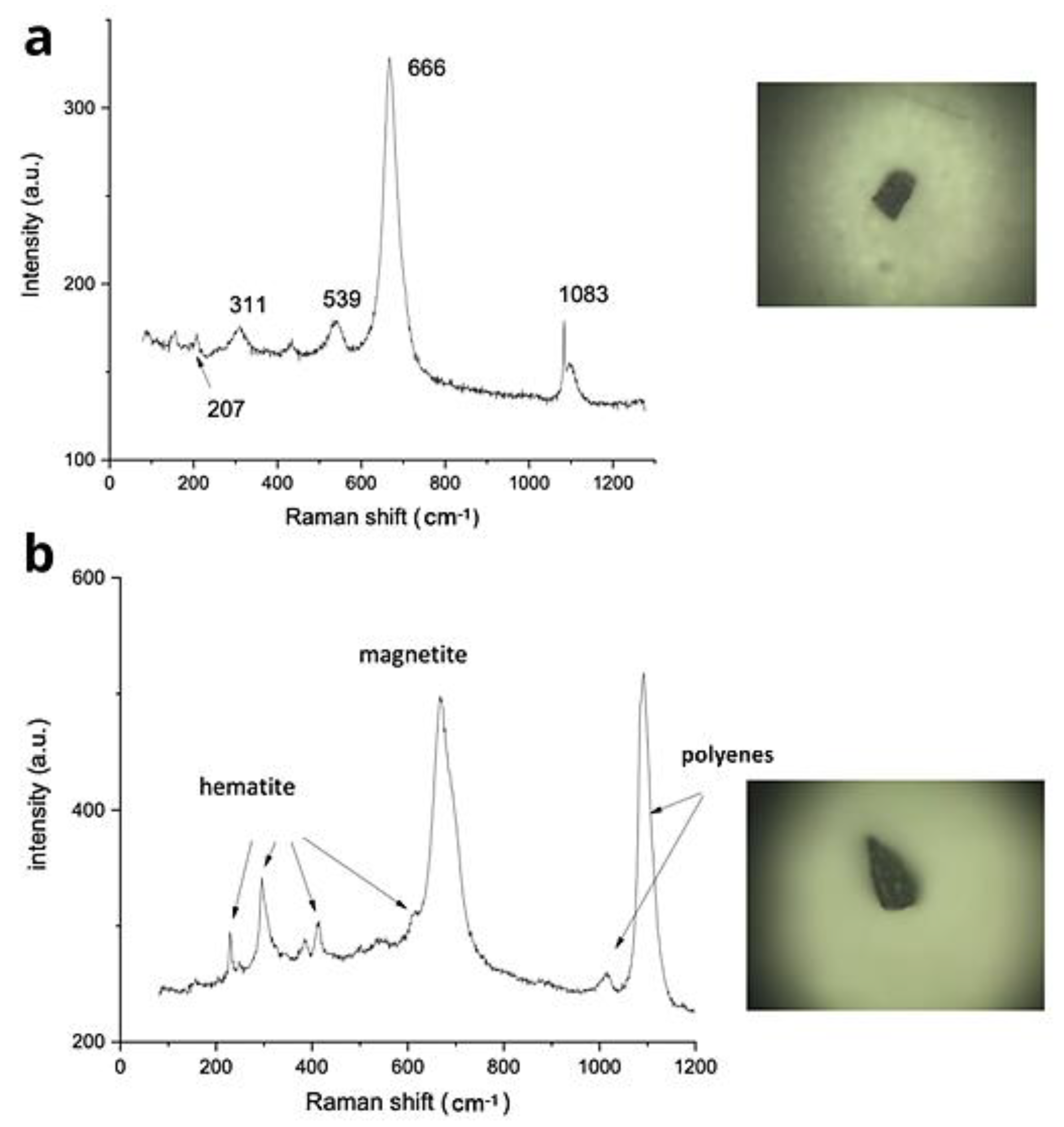
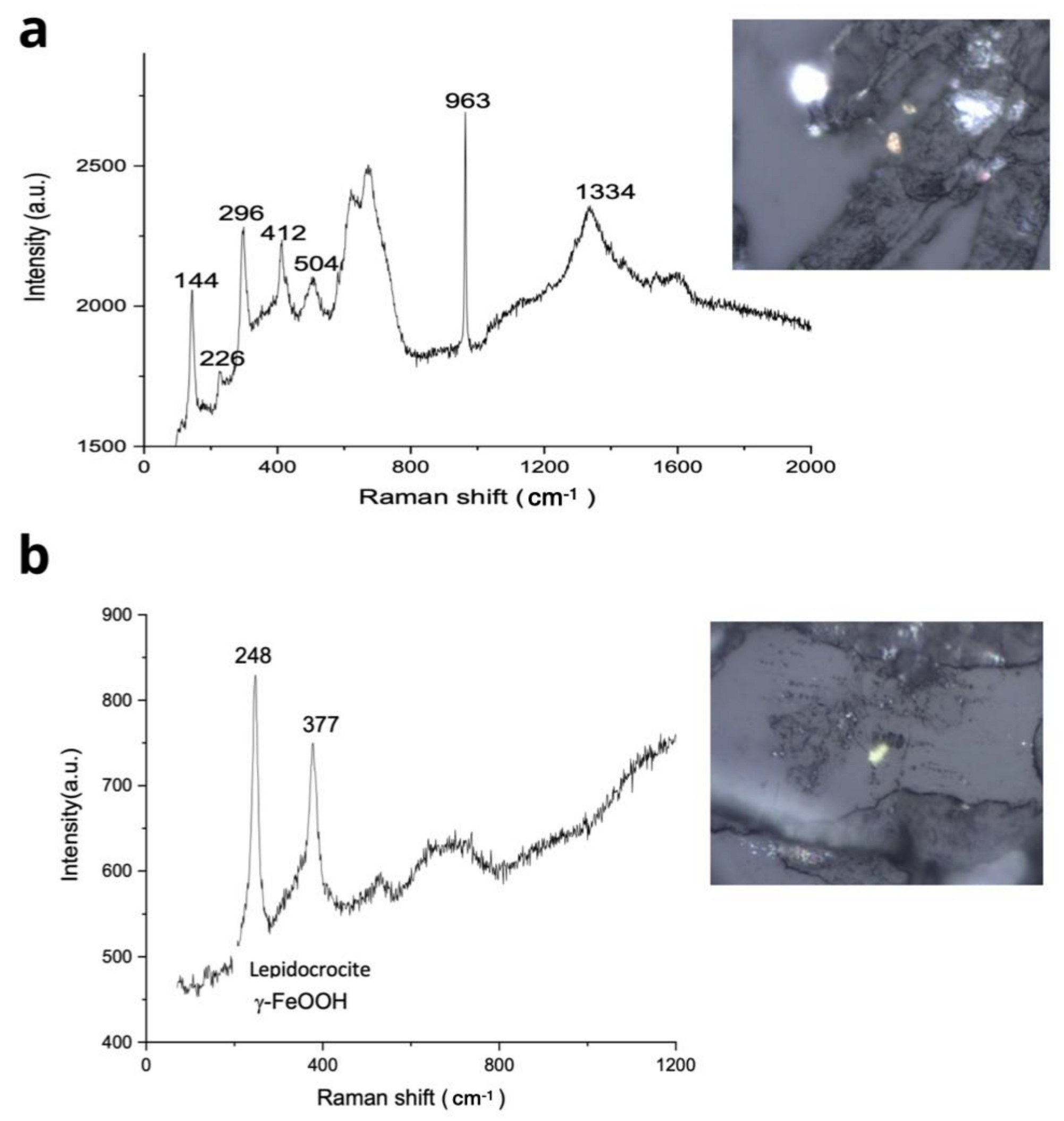
3.7. Raman Spectroscopy
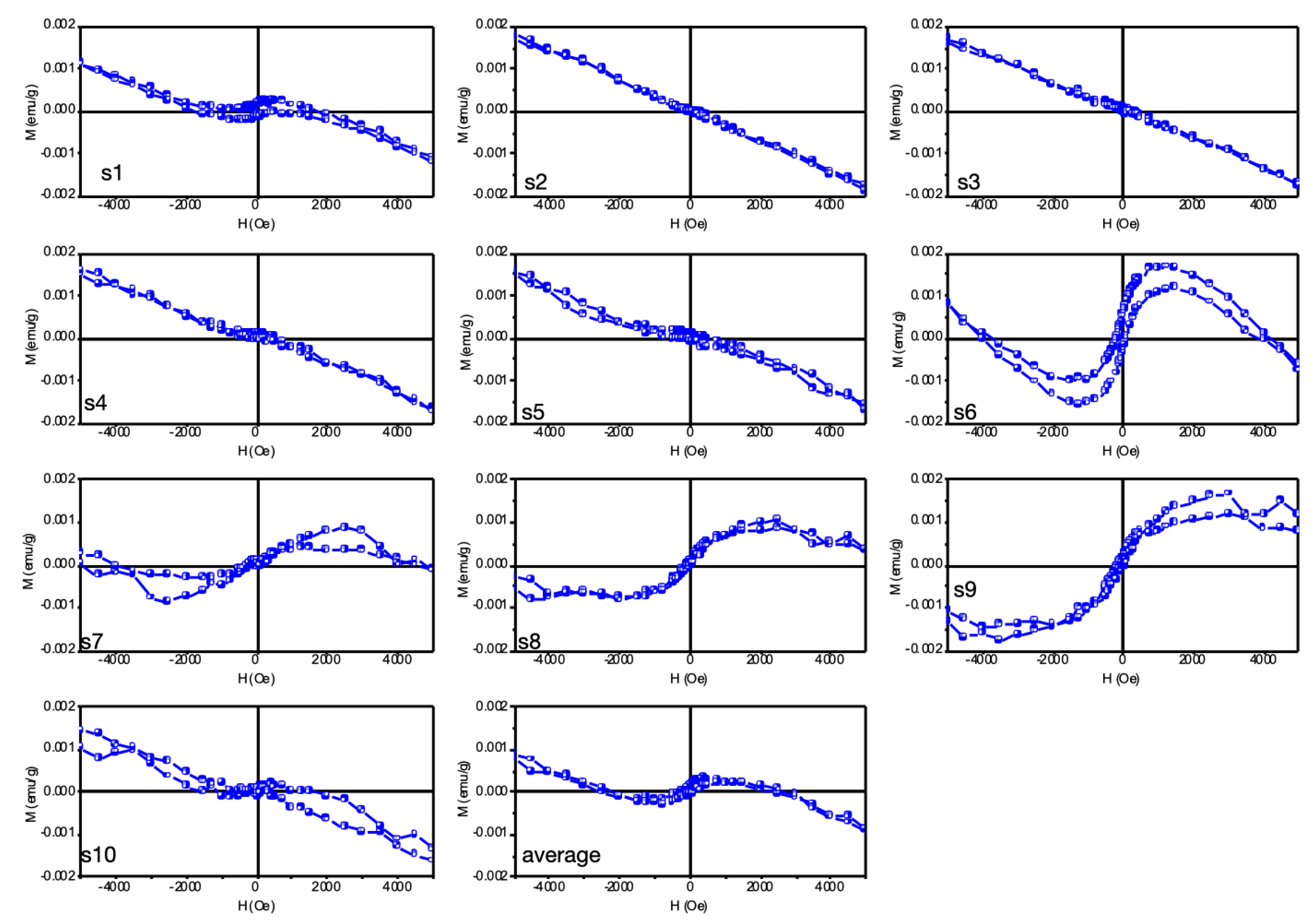
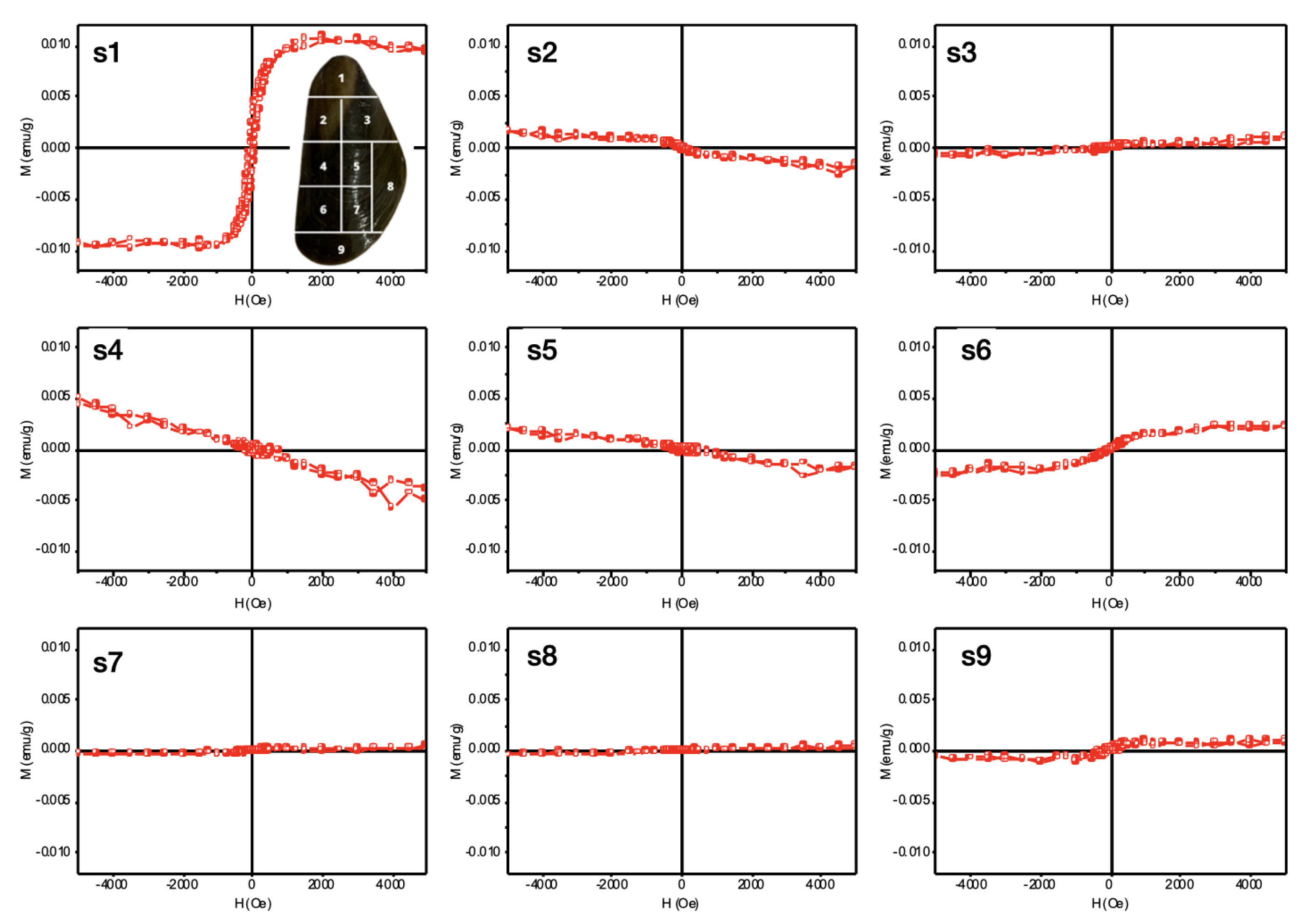
3.8. Vibrating Sample Magnetometer (VSM)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lowenstam, H.A. Magnetite in denticle capping in recent chitons (Polyplacophora). Geol. Soc. Am. Bull. 1962, 73, 435–438. [Google Scholar] [CrossRef]
- Bellinger, M.R.; Wei, J.; Hartmann, U.; Cadiou, H.; Winklhofer, M.; Banks, M.A. Conservation of magnetite biomineralization genes in all domains of life and implications for magnetic sensing. Proc. Natl. Acad. Sci. USA 2022, 119, e2108655119. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Chen, C.; Miyamoto, N.; Li, R.; Sigwart, J.D.; Xu, T.; Sun, Y.; Wong, W.C.; Ip, J.C.; Zhang, W.; et al. The Scaly-foot Snail genome and implications for the origins of biomineralized armour. Nat. Commun. 2020, 11, 1657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamester, M.R.; Balzer, P.S.; Becker, D. Characterization of calcium carbonate obtained from oyster and mussel shells and incorporation in polypropylene. Mater. Res. 2012, 15, 204–208. [Google Scholar] [CrossRef] [Green Version]
- Linggawati, A. Preparation and characterization of calcium oxide heterogeneous catalyst derived from Anadara granosa shell for biodiesel synthesis. KnE Eng. 2016, 1. [Google Scholar] [CrossRef] [Green Version]
- Santana, P.; Aranda, D.A. Nacre morphology and chemical composition in Atlantic winged oyster Pteria colymbus (Röding, 1798). PeerJ 2021, 9, e11527. [Google Scholar] [CrossRef]
- Kelton, K.F.; Greer, A.L. Nucleation in Condensed Matter: Applications in Materials and Biology; Elsevier: Amsterdam, The Netherlands, 2010. [Google Scholar]
- Blundell, S. Magnetism in Condensed Matter; Oxford University Press: Oxford, UK, 2001. [Google Scholar]
- Chen, C.W. Magnetism and Metallurgy of Soft Magnetic Materials; Elsevier: Amsterdam, The Netherlands, 1977. [Google Scholar]
- Petit, H.; Davis, W.L.; Jones, R.G.; Hagler, H.K. Morphological studies on the calcification process in the fresh-water mussel Amblema. Tissue Cell 1980, 12, 13–28. [Google Scholar] [CrossRef]
- Di Lorenzo, F.; Burgos-Cara, A.; Ruiz-Agudo, E.; Putnis, C.V.; Prieto, M. Effect of ferrous iron on the nucleation and growth of CaCO3in slightly basic aqueous solutions. CrystEngComm 2016, 19, 447–460. [Google Scholar] [CrossRef]
- Hirata, A.A. Studies on shell formation. II A mantle-shell preparation for in vitro studies. Biol. Bull. 1953, 104, 394–397. [Google Scholar] [CrossRef]
- Jodrey, L.H. Studies on shell formation. III. Measurement of calcium deposition in shell and calcium turnover in mantle tissue using the mantle-shell preparation and Ca45. Biol. Bull. 1953, 104, 398–407. [Google Scholar] [CrossRef]
- Mount, A.S.; Wheeler, A.P.; Paradkar, R.P.; Snider, D. Hemocyte-mediated shell mineralization in the eastern oyster. Science 2004, 304, 297–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasti, H.; Parivar, K.; Baharara, J.; Iranshahi, M.; Namvar, F. Chitin from the mollusc chiton: Extraction, characterization, and chitosan preparation. Iran. J. Pharm. Res. 2017, 16, 366–379. [Google Scholar] [PubMed]
- Ehrlich, H.; Martinović, R.; Joksimović, D.; Petrenko, I.; Schiaparelli, S.; Wysokowski, M.; Tsurkan, D.; Stelling, A.L.; Springer, A.; Gelinsky, M.; et al. Conchixes: Organic scaffolds which resemble the size and shapes of mollusks shells, their isolation and potential multifunctional applications. Appl. Phys. A 2020, 126, 562. [Google Scholar] [CrossRef]
- Di Palma, L.; Ferrantelli, P.; Merli, C.; Biancifiori, F. Recovery of EDTA and metal precipitation from soil flushing solutions. J. Hazard. Mater. 2003, 103, 153–168. [Google Scholar] [CrossRef]
- Chen, X.; Su, Y.; Liu, H.; Jiang, T.; Yang, J. Mineral composition variation in the shells of freshwater mussel Anodonta woodiana at different growth stages. Invertebr. Reprod. Dev. 2017, 61, 274–282. [Google Scholar] [CrossRef]
- Gawad, S.S. Concentrations of heavy metals in water, sediment, and mollusk gastropod, Lanistes carinatus from Lake Manzala, Egypt. Egypt. J. Aquat. Res. 2018, 44, 77–82. [Google Scholar] [CrossRef]
- Rončević, S.; Pitarević Svedružić, L.; Smetiško, J.; Medaković, D. ICP-AES analysis of metal content in shell of mussel Mytilus galloprovincialis from Croatian coastal waters. Int. J. Environ. Anal. Chem. 2010, 90, 620–632. [Google Scholar] [CrossRef]
- Blesa, M.A.; Borghi, E.B.; Maroto, A.J.; Regazzoni, A.E. Adsorption of EDTA and iron—EDTA complexes on magnetite and the mechanism of dissolution of magnetite by EDTA. J. Colloid Interface Sci. 1984, 98, 295–305. [Google Scholar]
- Graf, D.L. Crystallographic tables for the rhombohedral carbonates. Am. Miner. J. Earth Planet. Mater. 1961, 46, 1283–1316. [Google Scholar]
- De Villiers, J.P. Crystal structures of aragonite, strontianite, and witherite. Am. Miner. J. Earth Planet. Mater. 1971, 56, 758–767. [Google Scholar]
- Nakamura Filho, A.; Almeida, A.C.; Riera, H.E.; Araújo, J.L.; Gouveia, V.J.; Carvalho, M.D.; Cardoso, A.V. Polymorphism of CaCO3 and microstructure of the shell of a Brazilian invasive mollusc (Limnoperna fortunei). Mater. Res. 2014, 17, 15–22. [Google Scholar] [CrossRef]
- Colliex, C.; Manoubi, T.; Ortiz, C. Electron-energy-loss-spectroscopy near-edge fine structures in the iron-oxygen system. Phys. Rev. B 1991, 44, 11402–11411. [Google Scholar] [CrossRef] [PubMed]
- Frankel, R.B.; Papaefthymiou, G.C.; Blakemore, R.P.; O’Brien, W. Fe3O4 precipitation in magnetotactic bacteria. Biochim. Biophys. Acta Mol. Cell Res. 1983, 763, 147–159. [Google Scholar] [CrossRef] [Green Version]
- Danan, H.; Herr, A.; Meyer, A.J.P. New Determinations of the Saturation Magnetization of Nickel and Iron. J. Appl. Phys. 1968, 39, 669–670. [Google Scholar] [CrossRef]
- Park, B.C.; Cho, J.; Kim, M.S.; Ko, M.J.; Pan, L.; Na, J.Y.; Kim, Y.K. Strategy to control magnetic coercivity by elucidating crystallization pathway-dependent microstructural evolution of magnetite mesocrystals. Nat. Commun. 2020, 11, 298. [Google Scholar] [CrossRef] [Green Version]
- Kemp, S.J.; Ferguson, R.M.; Khandhar, A.P.; Krishnan, K.M. Monodisperse magnetite nanoparticles with nearly ideal saturation magnetization. RSC Adv. 2016, 6, 77452–77464. [Google Scholar] [CrossRef]
- De Araujo, F.F.T.; Pires, M.A.; Frankel, R.B.; Bicudo, C.E.M. Magnetite and Magnetotaxis in Algae. Biophys. J. 1986, 50, 375–378. [Google Scholar] [CrossRef] [Green Version]
- Machado, J.; Lopes-Lima, M. Calcification mechanism in freshwater mussels: Potential targets for cadmium. Toxicol. Environ. Chem. 2011, 93, 1778–1787. [Google Scholar] [CrossRef]
- Cravo, A.; Foster, P.; Bebianno, M. Minor and trace elements in the shell of Patella aspera (Röding 1798). Environ. Int. 2002, 28, 295–302. [Google Scholar] [CrossRef]
- Waller, T.R. Scanning Electron Microscopy of Shell and Mantle in the Order Arcoida (Mollusca: Bivalvia); Smithsonian Institution Scholarly Press: Washington, DC, USA, 1980; pp. 1–58. [Google Scholar] [CrossRef]
- Simkiss, K.; Wilbur, K.M. Biomineralization. In Cell Biology and Mineral Deposition; Academic Press: San Diego, CA, USA, 1989. [Google Scholar]
- Alimbekova, S.R.; Ishmuratov, F.G.; Nosov, V.V.; Voloshin, A.I.; Alimbekov, R.I.; Dokichev, V.A. Electromagnetic field influence on the calcium carbonate crystallization in the iron ions Fe2+ presence in mineralized aqueous media. J. Phys. Conf. Ser. 2021, 1889, 022096. [Google Scholar] [CrossRef]
- Korchef, A. Effect of Operating Parameters and Foreign Ions on the Crystal Growth of Calcium Carbonate during Scale Formation: An Overview. In Crystallization and Applications; IntechOpen: London, UK, 2020. [Google Scholar]
- Herzog, R.E.; Shi, Q.; Patil, J.N.; Katz, J.L. Magnetic water treatment: The effect of iron on calcium carbonate nucleation and growth. Langmuir 1989, 5, 861–867. [Google Scholar] [CrossRef]
- De Leeuw, N.H. Molecular dynamics simulations of the growth inhibiting effect of Fe2+, Mg2+, Cd2+, and Sr2+ on calcite crystal growth. J. Phys. Chem. B 2002, 106, 5241–5249. [Google Scholar] [CrossRef]
- Langer, G.; Nehrke, G.; Baggini, C.; Rodolfo-Metalpa, R.; Hall-Spencer, J.M.; Bijma, J. Limpets counteract ocean acidification induced shell corrosion by thickening of aragonitic shell layers. Biogeosciences 2014, 11, 7363–7368. [Google Scholar] [CrossRef] [Green Version]
- Johnstone, M.B.; Gohad, N.V.; Falwell, E.P.; Hansen, D.C.; Hansen, K.M.; Mount, A.S. Cellular orchestrated biomineralization of crystalline composites on implant surfaces by the eastern oyster, Crassostrea virginica (Gmelin, 1791). J. Exp. Mar. Biol. Ecol. 2015, 463, 8–16. [Google Scholar] [CrossRef]
- Kirschvink, J.L.; Kobayashi-Kirschvink, A.; Woodford, B.J. Magnetite biomineralization in the human brain. Proc. Natl. Acad. Sci. USA 1992, 89, 7683–7687. [Google Scholar] [CrossRef]
- Greenebaum, B.; Barnes, F. (Eds.) . Bioengineering and Biophysical Aspects of Electromagnetic Fields; CRC Press: Boca Raton, FL, USA, 2007. [Google Scholar]
- Maher, B.A.; Ahmed, I.A.; Karloukovski, V.; MacLaren, D.A.; Foulds, P.G.; Allsop, D.; Mann, D.M.; Torres-Jardón, R.; Calderon-Garciduenas, L. Magnetite pollution nanoparticles in the human brain. Proc. Natl. Acad. Sci. USA 2016, 113, 10797–10801. [Google Scholar] [CrossRef]
- Monteil, C.L.; Benzerara, K.; Menguy, N.; Bidaud, C.C.; Michot-Achdjian, E.; Bolzoni, R.; Mathon, F.P.; Coutaud, M.; Alonso, B.; Garau, C.; et al. Intracellular amorphous Ca-carbonate and magnetite biomineralization by a magnetotactic bacterium affiliated to the Alphaproteobacteria. ISME J. 2021, 15, 1–18. [Google Scholar] [CrossRef]
- Du, H.; Amstad, E. Water: How does it influence the CaCO3 formation? Angew. Chem. Int. Ed. 2020, 59, 1798–1816. [Google Scholar] [CrossRef]
- Du, H.; Steinacher, M.; Borca, C.; Huthwelker, T.; Murello, A.; Stellacci, F.; Amstad, E. Amorphous CaCO3: Influence of the formation time on its degree of hydration and stability. J. Am. Chem. Soc. 2018, 140, 14289–14299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirschvink, J.L.; Hagadorn, J.W. 10 A Grand Unified theory of Biomineralization. In The Biomineralization of Nano- and Micro-Structures; Bäuerlein, E., Ed.; Wiley-VCH Verlag GmbH: Weinheim, Germany, 2000; pp. 139–150. [Google Scholar]
- Spadiut, O.; Pekarsky, A. Intrinsically magnetic cells: A review on their natural occurrence and synthetic generation. Front. Bioeng. Biotechnol. 2020, 8, 1234. [Google Scholar]
- Lin, W.; Kirschvink, J.L.; Paterson, G.A.; Bazylinski, D.A.; Pan, Y. On the origin of microbial magnetoreception. Natl. Sci. Rev. 2019, 7, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Gorobets, S.; Gorobets, O.; Gorobets, Y.; Bulaievska, M. Chain-Like Structures of Biogenic and Nonbiogenic Magnetic Nanoparticles in Vascular Tissues. Bioelectromagnetics 2022, 43, 119–143. [Google Scholar] [CrossRef] [PubMed]
- Espinosa, H.D.; Juster, A.L.; Latourte, F.J.; Loh, O.Y.; Gregoire, D.; Zavattieri, P.D. Tablet-level origin of toughening in abalone shells and translation to synthetic composite materials. Nat. Commun. 2011, 2, 173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, A.; Parveen, S.; Chanda, D.K.; Aditya, G. An insight into the structure, composition and hardness of a biological material: The shell of freshwater mussels. RSC Adv. 2020, 10, 29543–29554. [Google Scholar] [CrossRef] [PubMed]
- Ivanina, A.V.; Falfushynska, H.I.; Beniash, E.; Piontkivska, H.; Sokolova, I.M. Biomineralization-related specialization of hemocytes and mantle tissues of the Pacific oysters Crassostrea gigas. J. Exp. Biol. 2017, 220, 3209–3221. [Google Scholar] [CrossRef] [Green Version]
- Ivanina, A.V.; Borah, B.M.; Vogts, A.; Malik, I.; Wu, J.; Chin, A.R.; Almarza, A.J.; Kumta, P.; Piontkivska, H.; Beniash, E.; et al. Potential trade-offs between biomineralization and immunity revealed by shell properties and gene expression profiles of two closely related Crassostrea species. J. Exp. Biol. 2018, 221, jeb183236. [Google Scholar] [CrossRef] [Green Version]
- Ramesh, K.; Melzner, F.; Griffith, A.W.; Gobler, C.J.; Rouger, C.; Tasdemir, D.; Nehrke, G. In vivo characterization of bivalve larval shells: A confocal Raman microscopy study. J. R. Soc. Interface 2018, 15, 20170723. [Google Scholar] [CrossRef] [Green Version]
- Segovia-Campos, I.; Martignier, A.; Filella, M.; Jaquet, J.; Ariztegui, D. Micropearls and other intracellular inclusions of amorphous calcium carbonate: An unsuspected biomineralization capacity shared by diverse microorganisms. Environ. Microbiol. 2021, 24, 537–550. [Google Scholar] [CrossRef]
- Ajili, W.; Tovani, C.B.; Fouassier, J.; de Frutos, M.; Laurent, G.P.; Bertani, P.; Djediat, C.; Marin, F.; Auzoux-Bordenave, S.; Azaïs, T.; et al. Inorganic phosphate in growing calcium carbonate abalone shell suggests a shared mineral ancestral precursor. Nat. Commun. 2022, 13, 1496. [Google Scholar] [CrossRef] [PubMed]
- De Frutos, M.; Rodríguez-Navarro, A.B.; Li, X.; Checa, A.G. Nanoscale Analysis of the Structure and Composition of Biogenic Calcite Reveals the Biomineral Growth Pattern. ACS Nano 2023, 17, 2829–2839. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Sample Weight | Year | Al | Ba | Be | Ca | Cd | Co | Cr | Cu | Fe | K | Li |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (g) | μg/g | % | μg/g | ||||||||||
| Limnoperna fortunei Shell | 0.5 | 2016 | 51.0 | 159.05 | <0.25 | 35.08 | <0.5 | <1 | 2.37 | 5.2 | 395.0 | 31.8 | <0.2 |
| Limnoperna fortunei Shell | - | 2022 | 41.1 | 161.1 | <0.2 | 36.07 | <0.5 | <1 | <4 | 1.2 | 105.9 | 36.7 | <0.5 |
| Perna perna Shell | - | 2022 | 21.0 | 3.3 | <0.2 | 38.64 | <0.5 | <1 | <4 | <0.5 | 19.8 | 133.1 | 1.3 |
| Sample Weight | Year | Mg | Mn | Na | Ni | Pb | Sn | Sr | Ti | V | Zn | Hg | |
| (g) | μg/g | ||||||||||||
| Limnoperna fortunei Shell | 0.5 | 2016 | 235 | 180 | 2200 | 2.86 | 23.05 | <15.0 | 585 | 22.8 | 2.81 | 90.4 | <0.01 |
| Limnoperna fortunei Shell | - | 2022 | 180.2 | 30.9 | 5371.5 | <4 | <10 | <30 | 1200.9 | 3.2 | <1 | 2.7 | <0.02 |
| Perna perna Shell | - | 2022 | 159.0 | 12.6 | 6630.2 | <4 | <10 | <30 | 1054.6 | <1 | <1 | <2 | <0.02 |
| Phase | Chemistry Formula | Space Group | Crystalline System | Concentration (%) |
|---|---|---|---|---|
| Freshwater Limnoperna fortunei (golden mussel) | ||||
| Aragonite | CaCO3 | Pmcn | Orthorhombic | 97.48 |
| Calcite | CaCO3 | R3c | Trigonal | 2.52 |
| Marine Perna perna (brown mussel) | ||||
| Aragonite | CaCO3 | Pmcn | Orthorhombic | >99.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cardoso, A.V.; Souza, C.C.; Dantas, M.S.; Machado, C.S.; Freitas, E.T.; Krohling, A.; Rosario, V.M.; Nappi, G.U.; Heneine, L.D. Characterization of Magnetic Nanoparticles from the Shells of Freshwater Mussel L. fortunei and Marine Mussel P. perna. Appl. Nano 2023, 4, 191-212. https://doi.org/10.3390/applnano4030011
Cardoso AV, Souza CC, Dantas MS, Machado CS, Freitas ET, Krohling A, Rosario VM, Nappi GU, Heneine LD. Characterization of Magnetic Nanoparticles from the Shells of Freshwater Mussel L. fortunei and Marine Mussel P. perna. Applied Nano. 2023; 4(3):191-212. https://doi.org/10.3390/applnano4030011
Chicago/Turabian StyleCardoso, Antonio Valadão, Clara Carvalho Souza, Maria Sylvia Dantas, Camila Schults Machado, Erico Tadeu Freitas, Alisson Krohling, Veronica Martins Rosario, Giancarlo Ubaldo Nappi, and Luiz Dias Heneine. 2023. "Characterization of Magnetic Nanoparticles from the Shells of Freshwater Mussel L. fortunei and Marine Mussel P. perna" Applied Nano 4, no. 3: 191-212. https://doi.org/10.3390/applnano4030011