The Development and Decay of the Circadian Clock in Drosophila melanogaster


Abstract
:1. Introduction
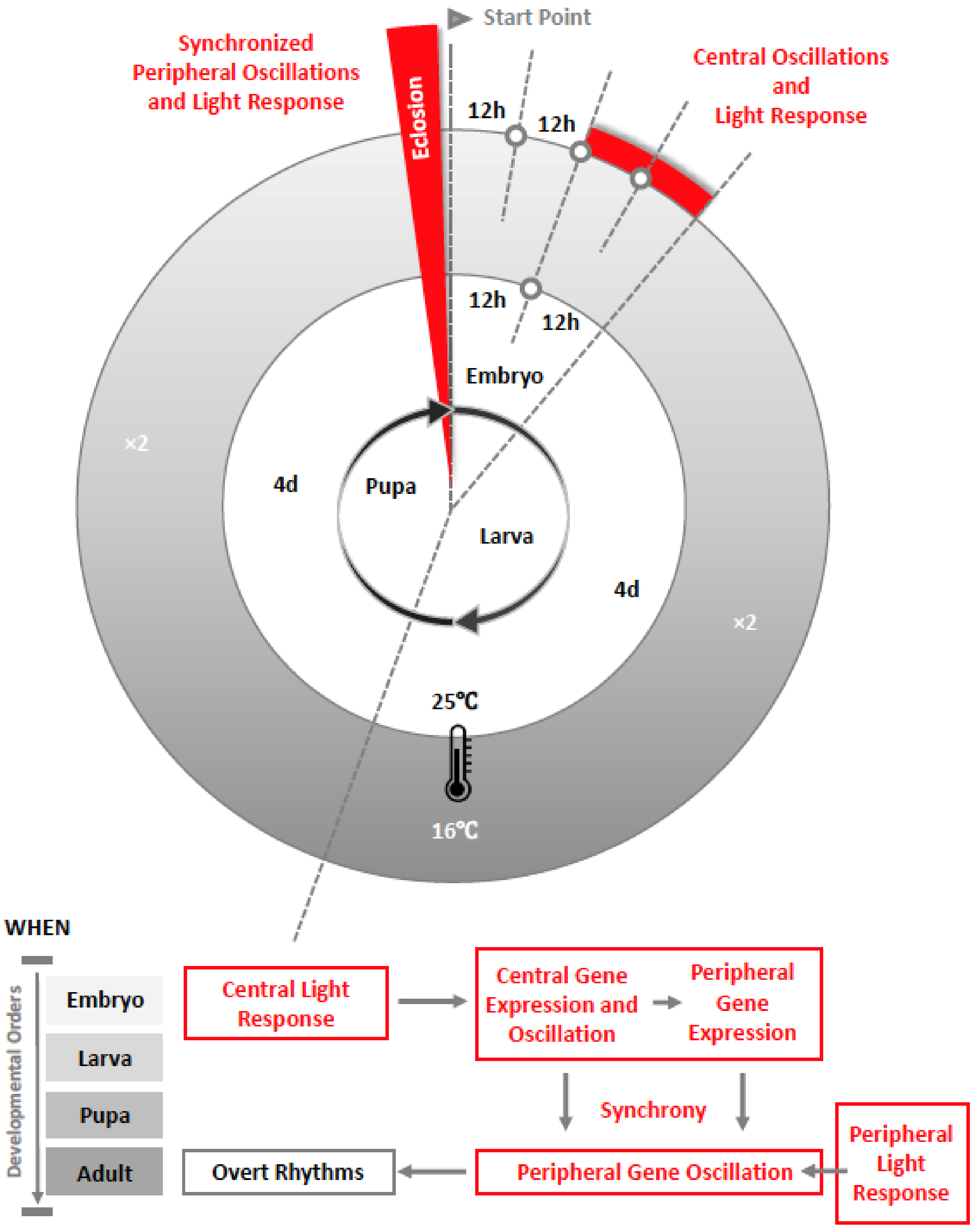
2. The Development of the Circadian Clock in Drosophila
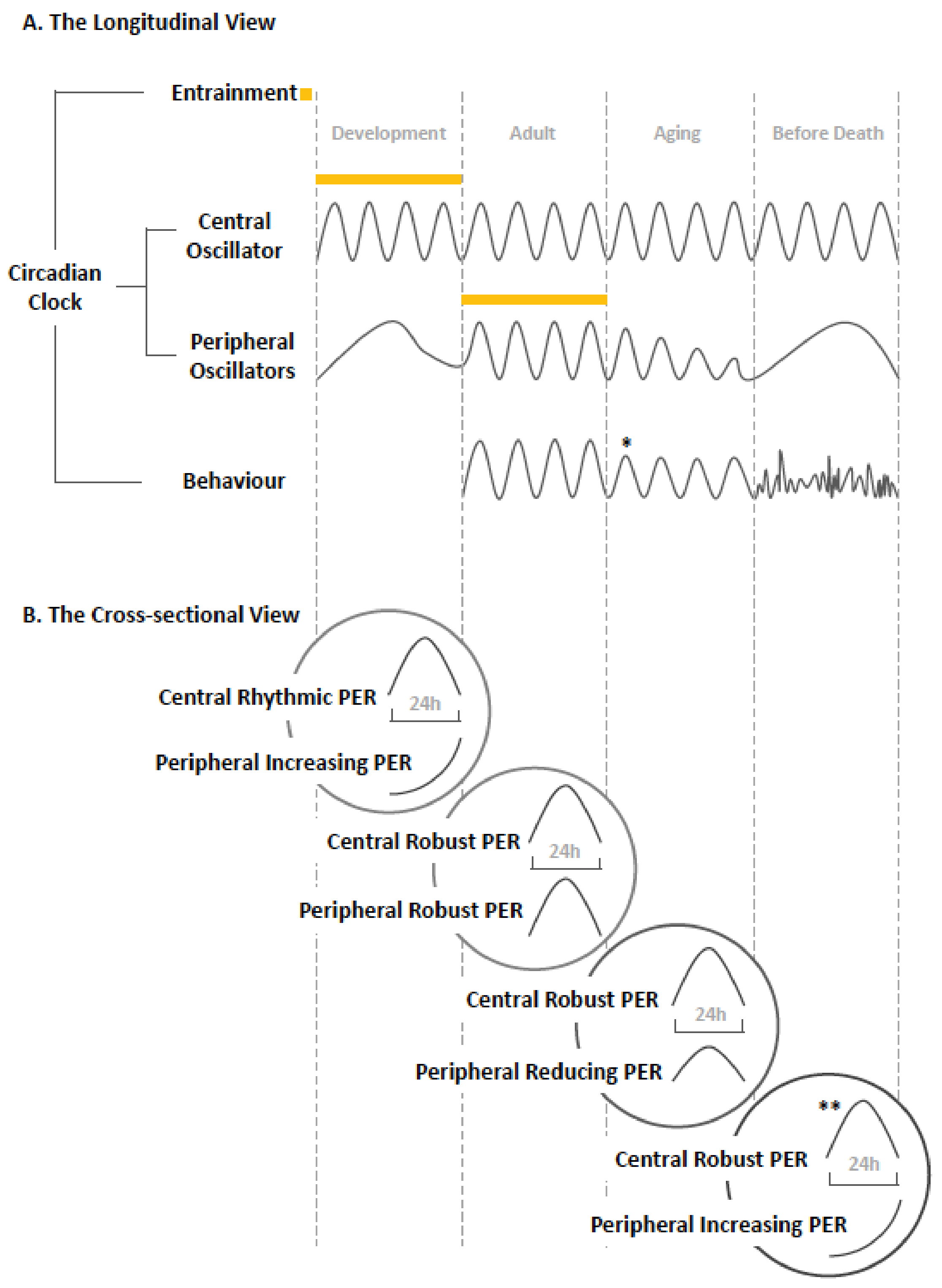
3. The Aging of the Circadian Clock in Drosophila
4. Discussion
5. Conclusions: The Dynamic Network
Author Contributions
Funding
Conflicts of Interest
References
- Nakamura, T.J.; Nakamura, W.; Yamazaki, S.; Kudo, T.; Cutler, T.; Colwell, C.S.; Block, G.D. Age-related decline in circadian output. J. Neurosci. 2011. [Google Scholar] [CrossRef] [PubMed]
- The 2017 Nobel Prize in Physiology or Medicine—Press Release. Available online: https://www.nobelprize.org/prizes/medicine/2017/press-release/ (accessed on 9 October 2019).
- Mazzoni, E.O.; Desplan, C.; Blau, J. Circadian pacemaker neurons transmit and modulate visual information to control a rapid behavioral response. Neuron 2005, 45, 293–300. [Google Scholar] [CrossRef] [PubMed]
- Skopik, S.D.; Pittendrigh, C.S. Circadian systems, II. The oscillation in the individual Drosophila pupa; its independence of developmental stage. Proc. Natl. Acad. Sci. USA 1967, 58, 1862–1869. [Google Scholar] [CrossRef] [PubMed]
- Brett, W.J. Persistent Diurnal Rhythmicity in Drosophila Emergence1. Ann. Entomol. Soc. Am. 1955, 48, 119–131. [Google Scholar] [CrossRef]
- Sehgal, A.; Price, J.; Young, M.W. Ontogeny of a biological clock in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1992, 89, 1423–1427. [Google Scholar] [CrossRef]
- Tomioka, K.; Uwozumi, K.; Matsumoto, N. Light Cycles Given During Development Affect Freerunning Period of Circadian Locomotor Rhythm of. J. Insect Physiol. 1997, 43, 1–9. [Google Scholar] [CrossRef]
- Kaneko, M.; Hamblen, M.J.; Hall, J.C. Involvement of the period gene in developmental time-memory: Effect of the perShort mutation on phase shifts induced by light pulses delivered to Drosophila larvae. J. Biol. Rhythms 2000, 15, 13–30. [Google Scholar] [CrossRef]
- Sheeba, V.; Chandrashekaran, M.; Joshi, A.; Sharma, V. Developmental plasticity of the locomotor activity rhythm of Drosophila melanogaster. J. Insect Physiol. 2002, 48, 25–32. [Google Scholar] [CrossRef]
- Seggio, J.A.; Possidente, B.; Ahmad, S.T. Larval Ethanol Exposure Alters Adult Circadian Free-Running Locomotor Activity Rhythm in Drosophila melanogaster. Chronobiol. Int. 2012, 29, 75–81. [Google Scholar] [CrossRef]
- Ahmad, S.T.; Steinmetz, S.B.; Bussey, H.M.; Possidente, B.; Seggio, J.A. Larval ethanol exposure alters free-running circadian rhythm and per Locus transcription in adult D. melanogaster period mutants. Behav. Brain Res. 2013, 241, 50–55. [Google Scholar] [CrossRef]
- Keene, A.C.; Sprecher, S.G. Seeing the light: Photobehavior in fruit fly larvae. Trends Neurosci. 2012, 35, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Keene, A.C.; Mazzoni, E.O.; Zhen, J.; Younger, M.A.; Yamaguchi, S.; Blau, J.; Desplan, C.; Sprecher, S.G. Distinct Visual Pathways Mediate Drosophila Larval Light Avoidance and Circadian Clock Entrainment. J. Neurosci. 2011, 31, 6527–6534. [Google Scholar] [CrossRef] [PubMed]
- Klarsfeld, A.; Picot, M.; Vias, C.; Chelot, E.; Rouyer, F.; Chélot, E.; Rouyer, F. Identifying specific light inputs for each subgroup of brain clock neurons in Drosophila larvae. J. Neurosci. 2011, 31, 17406–17415. [Google Scholar] [CrossRef] [PubMed]
- Helfrich-Förster, C.; Edwards, T.; Yasuyama, K.; Wisotzki, B.; Schneuwly, S.; Stanewsky, R.; Meinertzhagen, I.A.; Hofbauer, A. The extraretinal eyelet of Drosophila: Development, ultrastructure, and putative circadian function. J. Neurosci. 2002, 22, 9255–9266. [Google Scholar] [CrossRef] [PubMed]
- Malpel, S.; Klarsfeld, A.; Rouyer, F. Circadian Synchronization and Rhythmicity in Larval Photoperception-Defective Mutants of Drosophila. J. Biol. Rhythms 2004, 19, 10–21. [Google Scholar] [CrossRef]
- Picot, M.; Klarsfeld, A.; Chelot, E.; Malpel, S.; Rouyer, F.; Chélot, E.; Malpel, S.; Rouyer, F. A Role for Blind DN2 Clock Neurons in Temperature Entrainment of the Drosophila Larval Brain. J. Neurosci. 2009, 29, 8312–8320. [Google Scholar] [CrossRef]
- Malpel, S.; Klarsfeld, A.; Rouyer, F. Larval optic nerve and adult extra-retinal photoreceptors sequentially associated with clock neurons during Drosophila brain development. Development 2002, 129, 1443–1453. [Google Scholar]
- Yan, Y.; Xu, Y.; Deng, S.; Huang, N.; Yang, Y.; Qiu, J.; Liu, J.; Wang, X.; Yang, G.; Gu, H. A pair of identified giant visual projection neurons demonstrates rhythmic activities before eclosion. Neurosci. Lett. 2013, 550, 156–161. [Google Scholar] [CrossRef]
- Helfrich-Förster, C. Development of pigment-dispersing hormone-immunoreactive neurons in the nervous system of Drosophila melanogaster. J. Comp. Neurol. 1997. [Google Scholar] [CrossRef]
- Kaneko, M.; Helfrich-Förster, C.; Hall, J.C. Spatial and temporal expression of the period and timeless genes in the developing nervous system of drosophila: Newly identified pacemaker candidates and novel features of clock gene product cycling. J. Neurosci. 1997, 17, 6745–6760. [Google Scholar] [CrossRef]
- Frisch, B.; Hardin, P.E.; Hamblen-Coyle, M.J.; Rosbash, M.; Hall, J.C. A promoterless period gene mediates behavioral rhythmicity and cyclical per expression in a restricted subset of the drosophila nervous system. Neuron 1994. [Google Scholar] [CrossRef]
- Helfrich-Förster, C. Robust circadian rhythmicity of Drosophila melanogaster requires the presence of lateral neurons: A brain-behavioral study of disconnected mutants. J. Comp. Physiol. A Sens. Neural Behav. Physiol. 1998. [Google Scholar] [CrossRef] [PubMed]
- Renn, S.C.P.; Park, J.H.; Rosbash, M.; Hall, J.C.; Taghert, P.H. A pdf neuropeptide gene mutation and ablation of PDF neurons each cause severe abnormalities of behavioral circadian rhythms in Drosophila. Cell 1999. [Google Scholar] [CrossRef]
- Helfrich-Förster, C.; Shafer, O.T.; Wülbeck, C.; Grieshaber, E.; Rieger, D.; Taghert, P. Development and morphology of the clock-gene-expressing lateral neurons of Drosophila melanogaster. J. Comp. Neurol. 2007, 500, 47–70. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, M.; Hall, J.C. Neuroanatomy of cells expressing clock genes in Drosophila: Transgenic manipulation of the period and timeless genes to mark the perikarya of circadian pacemaker neurons and their projections. J. Comp. Neurol. 2000. [Google Scholar] [CrossRef]
- Nitabach, M.N.; Holmes, T.C.; Blau, J. Membranes, ions, and clocks: Testing the Njus-Sulzman-Hastings model of the circadian oscillator. Methods Enzymol. 2005, 393, 682–693. [Google Scholar]
- James, A.A.; Ewer, J.; Reddy, P.; Hall, J.C.; Rosbash, M. Embryonic expression of the period clock gene in the central nervous system of Drosophila melanogaster. EMBO J. 1986, 5, 2313–2320. [Google Scholar] [CrossRef]
- Ruiz, S.; Rickert, C.; Berger, C.; Technau, G.M.; Cantera, R. Spatio-temporal pattern of cells expressing the clock genes period and timeless and the lineages of period expressing neurons in the embryonic CNS of Drosophila melanogaster. Gene Expr. Patterns 2010, 10, 274–282. [Google Scholar] [CrossRef]
- Houl, J.H.; Ng, F.; Taylor, P.; Hardin, P.E. CLOCK expression identifies developing circadian oscillator neurons in the brains of Drosophila embryos. BMC Neurosci. 2008, 9, 119. [Google Scholar] [CrossRef]
- Glossop, N.R.J.; Gummadova, J.O.; Ghangrekar, I.; Hardin, P.E.; Coutts, G.A. Effects of TWIN-OF-EYELESS on Clock Gene Expression and Central-Pacemaker Neuron Development in Drosophila. J. Biol. Rhythms 2014, 29, 151–166. [Google Scholar] [CrossRef]
- Zhao, J.; Stanewsky, R.; Warman, G.R.; Cheeseman, J.F. Development of the Molecular Circadian Clock and Its Light Sensitivity in Drosophila Melanogaster. J. Biol. Rhythms 2019, 34, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Lorenz, L.; Yu, Q.N.; Hall, J.C.; Rosbash, M. Spatial and temporal expression of the period gene in Drosophila melanogaster. Genes Dev. 1988, 2, 228–238. [Google Scholar] [CrossRef] [PubMed]
- Kuntz, S.G.; Eisen, M.B. Drosophila Embryogenesis Scales Uniformly across Temperature in Developmentally Diverse Species. PLoS Genet. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Economos, A.C.; Lints, F.A. Developmental temperature and life span in drosophila melanogaster: I. constant developmental temperature: Evidence for physiological adaptation in a wide temperature range. Gerontology 1986. [Google Scholar] [CrossRef]
- Ghosh, S.M.; Testa, N.D.; Shingleton, A.W. Temperature-size rule is mediated by thermal plasticity of critical size in Drosophila melanogaster. Proc. R. Soc. B Biol. Sci. 2013, 280, 20130174. [Google Scholar] [CrossRef] [Green Version]
- Ivanchenko, M.; Stanewsky, R.; Giebultowicz, J.M. Circadian photoreception in Drosophila: Functions of cryptochrome in peripheral and central clocks. J. Biol. Rhythms 2001, 16, 205–215. [Google Scholar] [CrossRef]
- Agrawal, P.; Houl, J.H.; Gunawardhana, K.L.; Liu, T.; Zhou, J.; Zoran, M.J.; Hardin, P.E. Drosophila CRY Entrains Clocks in Body Tissues to Light and Maintains Passive Membrane Properties in a Non-clock Body Tissue Independent of Light. Curr. Biol. 2017, 27, 2431–2441.e3. [Google Scholar] [CrossRef]
- Fonseca Costa, S.S.; Ripperger, J.A. Impact of the Circadian Clock on the Aging Process. Front. Neurol. 2015, 6, 6–43. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.J.; Nakamura, W.; Tokuda, I.T.; Ishikawa, T.; Kudo, T.; Colwell, C.S.; Block, G.D. Age-Related Changes in the Circadian System Unmasked by Constant Conditions. eNeuro 2015, 2. [Google Scholar] [CrossRef] [Green Version]
- Banks, G.; Nolan, P.M.; Peirson, S.N. Reciprocal interactions between circadian clocks and aging. Mamm. Genome 2016, 27, 332–340. [Google Scholar] [CrossRef]
- Kondratova, A.A.; Kondratov, R.V. The circadian clock and pathology of the ageing brain. Nat. Rev. Neurosci. 2012, 13, 325–335. [Google Scholar] [CrossRef] [PubMed]
- Koh, K.; Evans, J.M.; Hendricks, J.C.; Sehgal, A. A Drosophila model for age-associated changes in sleep:wake cycles. Proc. Natl. Acad. Sci. USA 2006, 103, 13843–13847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umezaki, Y.; Yoshii, T.; Kawaguchi, T.; Helfrich-Förster, C.; Tomioka, K. Pigment-dispersing factor is involved in age-dependent rhythm changes in Drosophila melanogaster. J. Biol. Rhythms 2012, 27, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Duffy, J.F.; Zitting, K.-M.M.; Chinoy, E.D. Aging and Circadian Rhythms. Sleep Med. Clin. 2015, 10, 423–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rakshit, K.; Krishnan, N.; Guzik, E.M.; Pyza, E.; Giebultowicz, J.M. Effects of aging on the molecular circadian oscillations in Drosophila. Chronobiol. Int. 2012, 29, 5–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giebultowicz, J.M.; Long, D.M. Aging and circadian rhythms. Curr. Opin. Insect Sci. 2015, 7, 82–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, W.; Chen, W.-F.F.; Yue, Z.; Chen, D.; Sowcik, M.; Sehgal, A.; Zheng, X. Old flies have a robust central oscillator but weaker behavioral rhythms that can be improved by genetic and environmental manipulations. Aging Cell 2012, 11, 428–438. [Google Scholar] [CrossRef]
- Zhao, J.; Warman, G.R.; Cheeseman, J.F. Clock gene expression and locomotor activity predict death in the last days of life in Drosophila melanogaster. Sci. Rep. 2018, 8, 11923. [Google Scholar] [CrossRef]
- Driver, C. The circadian clock in old Drosophila melanogaster. Biogerontology 2000, 1, 157–162. [Google Scholar] [CrossRef]
- Krishnan, N.; Kretzschmar, D.; Rakshit, K.; Chow, E.; Giebultowicz, J.M. The circadian clock gene period extends healthspan in aging Drosophila melanogaster. Aging (Albany NY) 2009, 1, 937–948. [Google Scholar] [CrossRef] [Green Version]
- Rakshit, K.; Giebultowicz, J.M. Cryptochrome restores dampened circadian rhythms and promotes healthspan in aging Drosophila. Aging Cell 2013, 12, 752–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuintzle, R.C.; Chow, E.S.; Westby, T.N.; Gvakharia, B.O.; Giebultowicz, J.M.; Hendrix, D.A. Circadian deep sequencing reveals stress-response genes that adopt robust rhythmic expression during aging. Nat. Commun. 2017, 8, 14529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pozhitkov, A.E.; Noble, P.A. Gene expression in the twilight of death: The increase of thousands of transcripts has implications to transplantation, cancer, and forensic research. Bioessays 2017. [Google Scholar] [CrossRef] [PubMed]
- Hood, S.; Amir, S. The aging clock: Circadian rhythms and later life. J. Clin. Investig. 2017, 127, 437–446. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Mohan, A.; Sharma, V.K. Circadian dysfunction reduces lifespan in Drosophila melanogaster. Chronobiol. Int. 2005, 22, 641–653. [Google Scholar] [CrossRef] [PubMed]
- Pittendrigh, C.S.; Minis, D.H. Circadian Systems: Longevity as a Function of Circadian Resonance in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1972, 69, 1537–1539. [Google Scholar] [CrossRef] [Green Version]
- Klarsfeld, A.; Rouyer, F. Effects of Circadian Mutations and LD Periodicity on the Life Span of Drosophila melanogaster. J. Biol. Rhythms 1998, 13, 471–478. [Google Scholar] [CrossRef]
- Vaccaro, A.; Issa, A.-R.R.; Seugnet, L.; Birman, S.; Klarsfeld, A. Drosophila Clock Is Required in Brain Pacemaker Neurons to Prevent Premature Locomotor Aging Independently of Its Circadian Function. PLoS Genet. 2017, 13, e1006507. [Google Scholar] [CrossRef] [Green Version]
- Hendricks, J.C.; Lu, S.; Kume, K.; Yin, J.C.-P.; Yang, Z.; Sehgal, A. Gender Dimorphism in the Role of cycle (BMAL1) in Rest, Rest Regulation, and Longevity in Drosophila melanogaster. J. Biol. Rhythms 2003, 18, 12–25. [Google Scholar] [CrossRef]
- Kyriacou, C.P.; Oldroyd, M.; Wood, J.; Sharp, M.; Hill, M. Clock mutations alter developmental timing in drosophila. Heredity (Edinb) 1990, 64, 395–401. [Google Scholar] [CrossRef]
- Hurd, M.W.; Ralph, M.R. The Significance of Circadian Organization for Longevity in the Golden Hamster. J. Biol. Rhythms 1998. [Google Scholar] [CrossRef] [PubMed]
- Oklejewicz, M.; Daan, S. Enhanced longevity in Tau mutant Syrian hamsters, Mesocricetus auratus. J. Biol. Rhythms 2002. [Google Scholar] [CrossRef] [Green Version]
- Paranjpe, D.A.; Anitha, D.; Chandrashekaran, M.K.; Joshi, A.; Sharma, V.K. Possible role of eclosion rhythm in mediating the effects of light-dark environments on pre-adult development in Drosophila melanogaster. BMC Dev. Biol. 2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vallone, D.; Lahiri, K.; Dickmeis, T.; Foulkes, N.S. Start the clock! Circadian rhythms and development. Dev. Dyn. 2007, 236, 142–155. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Vaze, K.M.; Kumar, D.; Sharma, V.K. Selection for early and late adult emergence alters the rate of pre-adult development in Drosophila melanogaster. BMC Dev. Biol. 2006, 6, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konopka, R.J.; Benzer, S. Clock Mutants of Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 1971, 68, 2112–2116. [Google Scholar] [CrossRef] [Green Version]
- Myers, E.M.; Yu, J.; Sehgal, A. Circadian control of eclosion: Interaction between a central and peripheral clock in Drosophila melanogaster. Curr. Biol. 2003, 13, 526–533. [Google Scholar] [CrossRef] [Green Version]
- Morioka, E.; Matsumoto, A.; Ikeda, M. Neuronal influence on peripheral circadian oscillators in pupal Drosophila prothoracic glands. Nat. Commun. 2012. [Google Scholar] [CrossRef] [Green Version]
- Selcho, M.; Millán, C.; Palacios-Muñoz, A.; Ruf, F.; Ubillo, L.; Chen, J.; Bergmann, G.; Ito, C.; Silva, V.; Wegener, C.; et al. Central and peripheral clocks are coupled by a neuropeptide pathway in Drosophila. Nat. Commun. 2017. [Google Scholar] [CrossRef]
- Girardot, F.; Lasbleiz, C.; Monnier, V.; Tricoire, H. Specific age related signatures in Drosophila body parts transcriptome. BMC Genomics 2006, 7, 69. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, J.; Warman, G.; Cheeseman, J. The Development and Decay of the Circadian Clock in Drosophila melanogaster. Clocks & Sleep 2019, 1, 489-500. https://doi.org/10.3390/clockssleep1040037
Zhao J, Warman G, Cheeseman J. The Development and Decay of the Circadian Clock in Drosophila melanogaster. Clocks & Sleep. 2019; 1(4):489-500. https://doi.org/10.3390/clockssleep1040037
Chicago/Turabian StyleZhao, Jia, Guy Warman, and James Cheeseman. 2019. "The Development and Decay of the Circadian Clock in Drosophila melanogaster" Clocks & Sleep 1, no. 4: 489-500. https://doi.org/10.3390/clockssleep1040037