
Delipidation of Chicken Feathers by Lipolytic Bacillus Species Isolated from River-Borne Sediments

Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection and Processing of Samples
2.2. Bacterial Isolation
2.3. Evaluation of Potential Lipase Activity
2.4. Enzyme Production and Assay for Lipase Activity
2.5. Lipolytic Bacteria Identification
2.6. Fermentation Condition Optimization to Enhance Lipase Production
2.7. Influence of Chicken Feather Concentration on the Medium Free Fatty Acid
2.8. Quantitation of the Crude Fat Content of Untreated and Biotreated Chicken Feathers
2.9. Statistical Analysis
3. Results
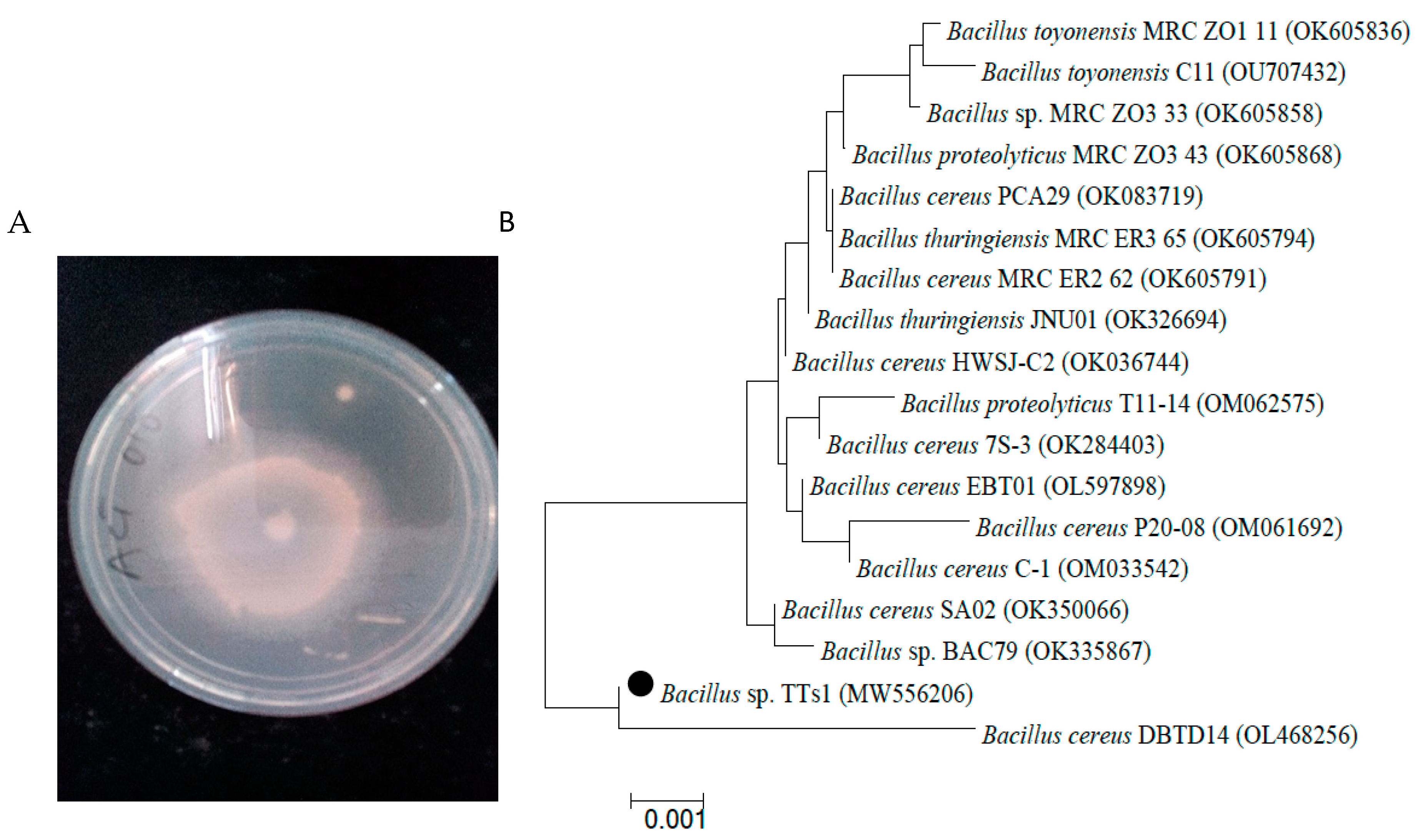
3.1. Lipolytic Bacterial Isolation and Identification
3.2. Optimization of the Fermentation Process Conditions
3.3. Influence of Chicken Feather Concentration on the Medium Free Fatty Acid
3.4. Quantitation of Crude Lipid Content of Untreated and TTs1-Pretreated Chicken Feathers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Laba, W.; Rodziewicz, A. Biodegradation of hard keratins by two Bacillus strains. Jundishapur J. Microbiol. 2014, 7, e8896. [Google Scholar] [CrossRef]
- Chilakamarry, C.R.; Mahmood, S.; Saffe, S.N.B.M.; Arifin, M.A.B.; Gupta, A.; Sikkandar, M.Y.; Begum, S.S.; Narasaiah, B. Extraction and application of keratin from natural resources: A review. 3 Biotech 2021, 11, 220. [Google Scholar] [CrossRef]
- Kokwe, L.; Nnolim, N.E.; Ezeogu, L.I.; Sithole, B.; Nwodo, U.U. Thermoactive metallo-keratinase from Bacillus sp. NFH5: Characterization, structural elucidation, and potential application as detergent additive. Heliyon 2023, 9, e13635. [Google Scholar] [CrossRef] [PubMed]
- Khumalo, M.; Tesfaye, T.; Sithole, B.B.; Ramjugernath, D. Possible beneficiation of waste chicken feathers via conversion into biomedical applications. Int. J. Chem. Sci. 2019, 17, 298. [Google Scholar] [CrossRef]
- Nnolim, N.E.; Udenigwe, C.C.; Okoh, A.I.; Nwodo, U.U. Microbial keratinase: Next generation green catalyst and prospective applications. Front. Microbiol. 2020, 11, 580164. [Google Scholar] [CrossRef]
- Bokveld, A.; Nnolim, N.E.; Nwodo, U.U. Chryseobacterium aquifrigidense FANN1 produced detergent-stable metallokeratinase and amino acids through the abasement of chicken feathers. Front. Bioeng. Biotechnol. 2021, 9, 720176. [Google Scholar] [CrossRef] [PubMed]
- Gurav, R.G.; Jadhav, J.P. A novel source of biofertilizer from feather biomass for banana cultivation. Environ. Sci. Pollut. Res. 2013, 20, 4532–4539. [Google Scholar] [CrossRef]
- European Union. Regulation 2019/1009, 2019. (EU) of the European Parliament and of the Council of 5 June 2019 Laying Down Rules on the Making Available on the Market of EU Fertilising Products and Amending Regulations (EC) No 1069/2009 and (EC) No 1107/2009 and Repealing Regulation (EC) No 2003/2003. Off. J. Eur. Union 2019, 170, 1–114. Available online: http://data.europa.eu/eli/reg/2019/1009/oj (accessed on 23 June 2023).
- Ningthoujam, D.S.; Tamereihao, K.; Mukhererjee, S.; Khunjamayum, R.; Devi, L.J.; Asem, R.S. Keratinaceous Wastes and Their Valorization through Keratinolytic Microorganisms. In Keratin; Blumenberg, M., Ed.; IntechOpen: London, UK, 2018. [Google Scholar] [CrossRef]
- Kakonke, G.; Tesfaye, T.; Sithole, B.; Ntunka, M. A novel method for rapid extraction of biofibres from waste chicken feathers. J. Solid Waste Technol. Manag. 2021, 47, 31–45. [Google Scholar] [CrossRef]
- European Union. Code of Good Practice for Poultry Farmers. Legal Obligations and Good Practice Guidelines for Poultry Farmers in Relation to Dead Birds and Poultry Litter. 2022. Available online: https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&ved=2ahUKEwifk7_csfKBAxXoMjQIHS9-C8oQFnoECBMQAQ&url=https%3A%2F%2Fassets.gov.ie%2F120140%2Fa247d156-9404-4c8a-aa99-605ffc3f66cf.pdf&usg=AOvVaw2DD4cfZraILeMsD-2ph0Qg&opi=89978449 (accessed on 23 June 2023).
- Mozhiarasi, V.; Natarajan, T.S. Slaughterhouse and poultry wastes: Management practices, feedstocks for renewable energy production, and recovery of value added products. Biomass Conv. Bioref. 2022. [Google Scholar] [CrossRef]
- South African Poultry Association (SAPA). Broiler Production Annual Report 2021. Available online: https://www.sapoultry.co.za/wp-content/uploads/2022/06/SAPA-ANNUAL-REPORT-2021.pdf (accessed on 24 June 2023).
- Chen, H.; Gao, S.; Li, Y.; Xu, H.J.; Li, W.; Wang, J.; Zhang, Y. Valorization of Livestock Keratin Waste: Application in Agricultural Fields. Int. J. Environ. Res. Public Health 2022, 19, 6681. [Google Scholar] [CrossRef]
- Conway, A. World Poultry Production at Nearly 123 Million Tons in 2018. 2019. Available online: https://www.wattagnet.com/articles/38018-world-poultry-production-at-nearly-123-million-tons-in-2018 (accessed on 21 June 2022).
- Godfrey, L.; Oelofse, S. Historical review of waste management and recycling in South Africa. Resources 2017, 6, 57. [Google Scholar] [CrossRef]
- Fagbemi, O.D.; Sithole, B.; Tesfaye, T. Optimization of keratin protein extraction from waste chicken feathers using hybrid pretreatment techniques. Sustain. Chem. Pharm. 2020, 17, 100267. [Google Scholar] [CrossRef]
- Kondamudi, N.; Strull, J.; Misra, M.; Mohapatra, S.K. A green process for producing biodiesel from feather meal. J. Agric. Food Chem. 2009, 57, 6163–6166. [Google Scholar] [CrossRef]
- Khumalo, M.; Sithole, B.; Tesfaye, T. Valorization of waste chicken feathers: Optimization of keratin extraction from waste chicken feathers by sodium bisulphite, sodium dodecyl sulphate and urea. J. Environ. Manag. 2020, 262, 110329. [Google Scholar] [CrossRef] [PubMed]
- Prajapati, S.; Koirala, S.; Anal, A.K. Bioutilization of chicken feather waste by newly isolated keratinolytic bacteria and conversion into protein hydrolysates with improved functionalities. Appl. Biochem. Biotechnol. 2021, 193, 2497–2515. [Google Scholar] [CrossRef]
- Fagbemi, O.D.; Sithole, B. Evaluation of waste chicken feather protein hydrolysate as a bio-based binder for particleboard production. Curr. Res. Green Sustain. Chem. 2021, 4, 100168. [Google Scholar] [CrossRef]
- Li, Z.W.; Liang, S.; Ke, Y.; Deng, J.J.; Zhang, M.S.; Lu, D.L.; Li, J.Z.; Luo, X.C. The feather degradation mechanisms of a new Streptomyces sp. isolate SCUT-3. Commun. Biol. 2020, 3, 191. [Google Scholar] [CrossRef]
- Adelina, A.; Feliatra, F.; Siregar, Y.I.; Suharman, I.; Pamukas, N.A. Fermented chicken feathers using Bacillus subtilis to improve the quality of nutrition as a fish feed material. IOP Conf. Ser. Earth Environ. Sci. 2019, 348, 012008. [Google Scholar] [CrossRef]
- Nealson, K.H. Sediment bacteria: Who’s there, what are they doing, and what’s new? Annu. Rev. Earth Planet. Sci. 1997, 25, 403–434. [Google Scholar] [CrossRef]
- Shrivastava, P.; Yandigeri, M.S.; Kumar, R.; Arora, D.K. Isolation and characterization of Streptomycetes with plant growth promoting potential from Mangrove ecosystem. Pol. J. Microbiol. 2015, 64, 339–349. [Google Scholar] [CrossRef]
- Praveen Kumar, P.; Preetam Raj, J.P.; Nimal Christhudas, I.V.S.; Sagaya Jansi, R.; Murugan, N.; Agastian, P.; Arunachalam, C.; Ali Alharbi, S. Screening of actinomycetes for enzyme and antimicrobial activities from the soil sediments of Northern Tamil Nadu, South India. J. Biol. Act. Prod. Nat. 2015, 5, 58–70. [Google Scholar] [CrossRef]
- Gopinath, S.C.; Hilda, A.; Anbu, P. Extracellular enzymatic activity profiles in fungi isolated from oil-rich environments. Mycoscience 2005, 46, 119–126. [Google Scholar] [CrossRef]
- Bharathi, D.; Rajalakshmi, G.; Komathi, S. Optimization and production of lipase enzyme from bacterial strains isolated from petrol spilled soil. J. King Saud Univ. Sci. 2019, 31, 898–901. [Google Scholar] [CrossRef]
- Kanjanavas, P.; Khuchareontaworn, S.; Khawsak, P.; Pakpitcharoen, A.; Pothivejkul, K.; Santiwatanakul, S.; Matsui, K.; Kajiwara, T.; Chansiri, K. Purification and characterization of organic solvent and detergent tolerant lipase from thermotolerant Bacillus sp. RN2. Int. J. Mol. Sci. 2010, 11, 3783–3792. [Google Scholar] [CrossRef] [PubMed]
- Turner, S.; Pryer, K.M.; Miao, V.P.; Palmer, J.D. Investigating deep phylogenetic relationships among cyanobacteria and plastids by small subunit rRNA sequence analysis 1. J. Eukaryot. Microbiol. 1999, 46, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Kralik, P.; Beran, V.; Pavlik, I. Enumeration of Mycobacterium avium subsp. paratuberculosis by quantitative real-time PCR, culture on solid media and optical densitometry. BMC Res. Notes 2012, 5, 114. [Google Scholar] [CrossRef] [PubMed]
- Vicentini-Polette, C.M.; Ramos, P.R.; Gonçalves, C.B.; De Oliveira, A.L. Determination of free fatty acids in crude vegetable oil samples obtained by high-pressure processes. Food Chem. X 2021, 12, 100166. [Google Scholar] [CrossRef]
- Suresh, K.S.; Suresh, P.V.; Kudre, T.G. Prospective ecofuel feedstocks for sustainable production. In Advances in Eco-Fuels for a Sustainable Environment; Woodhead Publishing: Cambridge, UK, 2019; pp. 89–117. [Google Scholar]
- Redfern, J.; Kinninmonth, M.; Burdass, D.; Verran, J. Using soxhlet ethanol extraction to produce and test plant material (essential oils) for their antimicrobial properties. J. Microbiol. Biol. Educ. 2014, 15, 45–46. [Google Scholar] [CrossRef]
- Laroche, M.; Perreault, V.; Marciniak, A.; Gravel, A.; Chamberland, J.; Doyen, A. Comparison of conventional and sustainable lipid extraction methods for the production of oil and protein isolate from edible insect meal. Foods 2019, 8, 572. [Google Scholar] [CrossRef] [PubMed]
- United States Department of Agriculture Food Safety and Inspection Service, Office of Public Health Science. 2009. Available online: https://www.fsis.usda.gov/wps/wcm/connect/dd881c92-c19b-4530-b6ee-931c368b8904/CLG_FAT_03.pdf?MOD=AJPERES (accessed on 6 March 2023).
- Dias, A.C.; Andreote, F.D.; Dini-Andreote, F.; Lacava, P.T.; Sá, A.L.; Melo, I.S.; Azevedo, J.L.; Araújo, W.L. Diversity and biotechnological potential of culturable bacteria from Brazilian mangrove sediment. World J. Microbiol. Biotechnol. 2009, 25, 1305–1311. [Google Scholar] [CrossRef]
- Jankoski, P.R.; Correa, A.P.F.; Brandelli, A.; Motta, A.S. Biological activity of bacteria isolated from wetland sediments collected from a conservation unit in the southern region of Brazil. An. Acad. Bras. Ciênc. 2021, 93, e20191269. [Google Scholar] [CrossRef] [PubMed]
- Lee, L.P.; Karbul, H.M.; Citartan, M.; Gopinath, S.C.; Lakshmipriya, T.; Tang, T.H. Lipase-secreting Bacillus species in an oil-contaminated habitat: Promising strains to alleviate oil pollution. BioMed Res. Int. 2015, 2015, 820575. [Google Scholar] [CrossRef]
- Lanka, S.; Latha, J.N.L. A short review on various screening methods to isolate potential lipase producers: Lipases-the present and future enzymes of biotech industry. Int. J. Biol. Chem. 2015, 9, 207–219. [Google Scholar] [CrossRef]
- Kanchana, R.; Muraleedharan, U.D.; Raghukumar, S. Alkaline lipase activity from the marine protists, thraustochytrids. World J. Microbiol. Biotechnol. 2011, 27, 2125–2131. [Google Scholar] [CrossRef]
- Pham, V.H.T.; Kim, J.; Chang, S.; Chung, W. Investigation of lipolytic-secreting bacteria from an artificially polluted soil using a modified culture method and optimization of their lipase production. Microorganisms 2021, 9, 2590. [Google Scholar] [CrossRef]
- Ghori, M.I.; Iqbal, M.J.; Hameed, A. Characterization of a novel lipase from Bacillus sp. isolated from tannery wastes. Braz. J. Microbiol. 2011, 42, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Sharma, N.; Pathania, S.; Handa, S. Purification and characterization of lipase by Bacillus methylotrophicus PS3 under submerged fermentation and its application in detergent industry. J. Genet. Eng. Biotechnol. 2017, 15, 369–377. [Google Scholar] [CrossRef]
- Suci, M.; Arbianti, R.; Hermansyah, H. Lipase production from Bacillus subtilis with submerged fermentation using waste cooking oil. IOP Conf. Ser. Earth Environ. Sci. 2018, 105, 012126. [Google Scholar] [CrossRef]
- Shart, A.O.; Elkhalil, E.A. Biochemical Characterisation of Lipase Produced by Bacillus spp. Isolated from Soil and Oil Effluent. Adv. Enzym. Res. 2020, 8, 39–48. [Google Scholar] [CrossRef]
- Bora, L.; Bora, M. Optimization of extracellular thermophilic highly alkaline lipase from thermophilic Bacillus sp. isolated from hotspring of Arunachal Pradesh, India. Braz. J. Microbiol. 2012, 43, 30–42. [Google Scholar] [CrossRef]
- Riaz, M.; Shah, A.A.; Hameed, A.; Hasan, F. Characterization of lipase produced by Bacillus sp. FH5 in immobilized and free state. Ann. Microbiol. 2010, 60, 169–175. [Google Scholar] [CrossRef]
- Saranya, P.; Kumari, H.S.; Jothieswari, M.; Rao, B.P.; Sekaran, G. Novel extremely acidic lipases produced from Bacillus species using oil substrates. J. Ind. Microbiol. Biotechnol. 2014, 41, 9–15. [Google Scholar] [CrossRef]
- Lv, P.J.; Qiang, S.; Liu, L.; Hu, C.Y.; Meng, Y.H. Dissolved-oxygen feedback control fermentation for enhancing β-carotene in engineered Yarrowia lipolytica. Sci. Rep. 2020, 10, 17114. [Google Scholar] [CrossRef]
- Singh, V.; Haque, S.; Niwas, R.; Srivastava, A.; Pasupuleti, M.; Tripathi, C. Strategies for fermentation medium optimization: An in-depth review. Front. Microbiol. 2017, 7, 2087. [Google Scholar] [CrossRef]
- Potumarthi, R.; Subhakar, C.; Vanajakshi, J.; Jetty, A. Effect of aeration and agitation regimes on lipase production by newly isolated Rhodotorula mucilaginosa–MTCC 8737 in stirred tank reactor using molasses as sole production medium. Appl. Biochem. Biotechnol. 2008, 151, 700–710. [Google Scholar] [CrossRef] [PubMed]
- Popoola1, B.M.; Olanbiwoninu, A.A. Characterisation of Some Selected Bacterial Strains from Vegetable Oil Contaminated Soil. Microbiol. Res. J. Int. 2021, 31, 22–37. [Google Scholar] [CrossRef]
- Olusesan, A.T.; Azura, L.K.; Abubakar, F.; Mohamed, A.K.S.; Radu, S.; Manap, M.Y.A.; Saari, N. Enhancement of thermostable lipase production by a genotypically identified extremophilic Bacillus subtilis NS 8 in a continuous bioreactor. J. Mol. Microbiol. Biotechnol. 2011, 20, 105–115. [Google Scholar] [CrossRef]
- Isiaka Adetunji, A.; Olufolahan Olaniran, A. Optimization of culture conditions for enhanced lipase production by an indigenous Bacillus aryabhattai SE3-PB using response surface methodology. Biotechnol. Biotechnol. Equip. 2018, 32, 1514–1526. [Google Scholar] [CrossRef]
- Saraswat, R.; Bhushan, I.; Gupta, P.; Kumar, V.; Verma, V. Production and purification of an alkaline lipase from Bacillus sp. for enantioselective resolution of (±)-Ketoprofen butyl ester. 3 Biotech 2018, 8, 491. [Google Scholar] [CrossRef] [PubMed]
- Ramani, K.; Kennedy, L.J.; Ramakrishnan, M.; Sekaran, G. Purification, characterization and application of acidic lipase from Pseudomonas gessardii using beef tallow as a substrate for fats and oil hydrolysis. Process Biochem. 2010, 45, 1683–1691. [Google Scholar] [CrossRef]
- Al Mohaini, M.; Farid, A.; Muzammal, M.; Ghazanfar, S.; Dadrasnia, A.; Alsalman, A.J.; Al Hawaj, M.A.; Alhashem, Y.N.; Ismail, S. Enhancing lipase production of Bacillus salmalaya strain 139si using different carbon sources and surfactants. Appl. Microbiol. 2022, 2, 237–247. [Google Scholar] [CrossRef]
- Soleymani, S.; Alizadeh, H.; Mohammadian, H.; Rabbani, E.; Moazen, F.; Sadeghi, H.M.; Shariat, Z.S.; Etemadifar, Z.; Rabbani, M. Efficient media for high lipase production: One variable at a time approach. Avicenna J. Med. Biotechnol. 2017, 9, 82–86. [Google Scholar]
- Hasan, F.; Shah, A.A.; Hameed, A. Influence of culture conditions on lipase production by Bacillus sp. FH5. Ann. Microbiol. 2006, 56, 247–252. [Google Scholar] [CrossRef]
- Haniya, M.; Naaz, A.; Sakhawat, A.; Amir, S.; Zahid, H.; Syed, S.A. Optimized production of lipase from Bacillus subtilis PCSIRNL-39. Afr. J. Biotechnol. 2017, 16, 1106–1115. [Google Scholar] [CrossRef]
- Adetunji, A.I.; Olaniran, A.O. Production strategies and biotechnological relevance of microbial lipases: A review. Braz. J. Microbiol. 2021, 52, 1257–1269. [Google Scholar] [CrossRef]
- Tesfaye, T.; Sithole, B.; Ramjugernath, D. Valorisation of waste chicken feathers: Optimisation of decontamination and pre-treatment with bleaching agents using response surface methodology. Sustain. Chem. Pharm. 2018, 8, 21–37. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Feather Sample | Crude Fat (%) |
|---|---|
| Untreated | 2.1 ± 0.42 a |
| TTs1-pretreated | 0.92 ± 0.13 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shiri, T.; Nnolim, N.E.; Nwodo, U.U. Delipidation of Chicken Feathers by Lipolytic Bacillus Species Isolated from River-Borne Sediments. Clean Technol. 2023, 5, 1235-1247. https://doi.org/10.3390/cleantechnol5040062
Shiri T, Nnolim NE, Nwodo UU. Delipidation of Chicken Feathers by Lipolytic Bacillus Species Isolated from River-Borne Sediments. Clean Technologies. 2023; 5(4):1235-1247. https://doi.org/10.3390/cleantechnol5040062
Chicago/Turabian StyleShiri, Tariro, Nonso E. Nnolim, and Uchechukwu U. Nwodo. 2023. "Delipidation of Chicken Feathers by Lipolytic Bacillus Species Isolated from River-Borne Sediments" Clean Technologies 5, no. 4: 1235-1247. https://doi.org/10.3390/cleantechnol5040062