
Unravelling the Combined Use of Soil and Microbial Technologies to Optimize Cultivation of Halophyte Limonium algarvense (Plumbaginaceae) Using Saline Soils and Water

 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microcosm Assay
2.2. Soils and Estuarine Water Analyses
2.3. Microbial Inocula
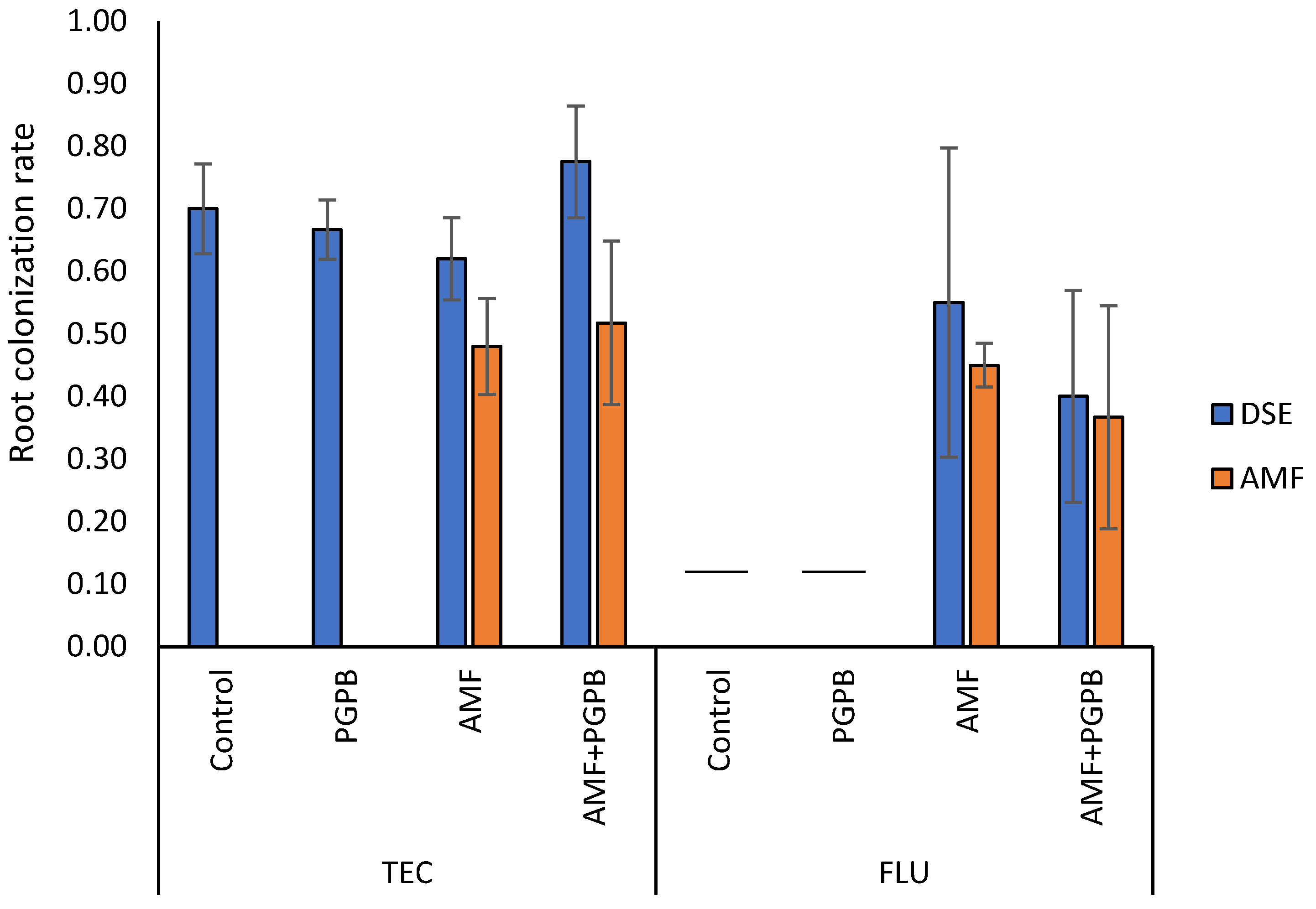
2.4. Plant Performance and Root Colonization Evaluation
2.5. Pollen Fertility
2.6. Statistical Analyses
3. Results
3.1. Characterisation of Irrigation Water and Soils
3.2. Evolution of Physiological Parameters of Plants with Microbial Inoculations Grown in Fluvisols (FLU) and Amended Fluvisols (TEC)
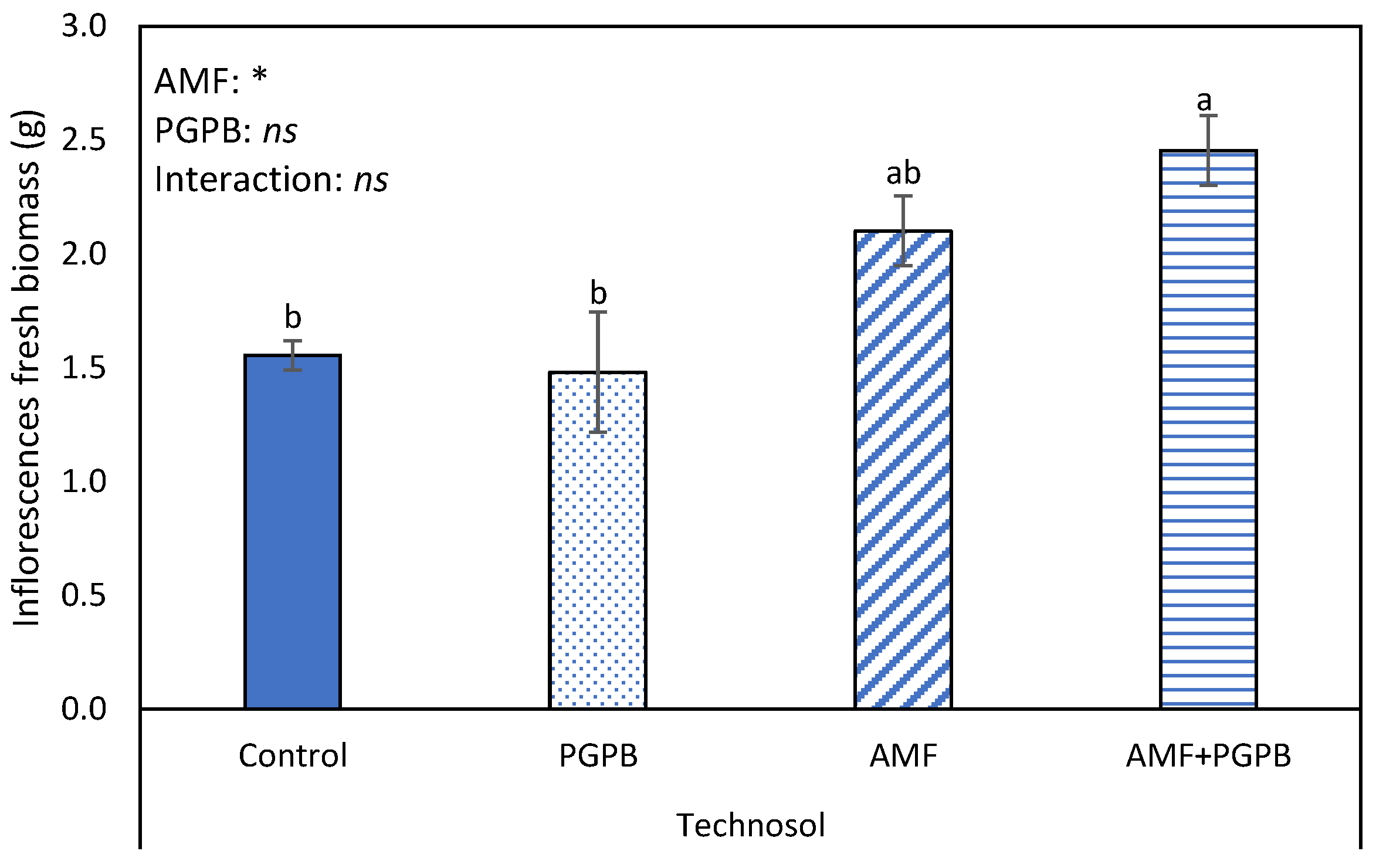
3.3. Effects of Microbial Inoculations and Technosols in L. algarvense Vegetative and Reproductive Growth
4. Discussion
4.1. Soil Technologies Improve Soil Properties
4.2. Microbial and Soil Technologies Improve Limonium algarvense Development
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stavi, I.; Thevs, N.; Priori, S. Soil Salinity and Sodicity in Drylands: A Review of Causes, Effects, Monitoring, and Restoration Measures. Front. Environ. Sci. 2021, 9, 330. [Google Scholar] [CrossRef]
- Omuto, C.T.; Vargas, R.R.; El Mobarak, A.M.; Mohamed, N.; Viatkin, K.; Yigini, Y. Mapping of Salt-Affected Soils—Technical Manual; Food and Agriculture Organization of the United Nations (FAO): Rome, Italy, 2020; Available online: https://www.fao.org/documents/card/fr/c/CA9215EN (accessed on 3 June 2023).
- Hulisz, P.; Pindral, S.; Kobierski, M.; Charzyński, P. Technogenic Layers in Organic Soils as a Result of the Impact of the Soda Industry. Eurasian Soil Sci. 2018, 51, 1133–1141. [Google Scholar] [CrossRef]
- WRB, (IUSS Working Group). World Reference Base for Soil Resources (2015) International Soil Classification System for Naming Soils and Creating Legends for Soil Maps, 4th ed.; WRB (IUSS Working Group): Vienna, Austria, 2019. [Google Scholar]
- Almeida, D. Ecology and Dynamics of Mediterranean Saltmarshes in a Perspective of Habitat Management and Restoration Policies: The Cases of Alvor and Arade in Portugal; Universidade de Lisboa: Lisbon, Portugal, 2016. [Google Scholar]
- Food and Agriculture Organization of the United Nations (FAO). Global Symposium on Salt-Affected Soils: Outcome Document; FAO: Rome, Italy, 2022; Available online: https://www.fao.org/documents/card/es/c/cb9929en/ (accessed on 3 June 2023).
- Elliott, M.; McLusky, D.S. The Need for Definitions in Understanding Estuaries. Estuar. Coast. Shelf. Sci. 2002, 55, 815–827. [Google Scholar] [CrossRef]
- Flowers, T.J.; Colmer, T.D. Salinity Tolerance in Halophytes. New Phytol. 2008, 179, 945–963. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Guo, J.; Shabala, S.; Wang, B. Reproductive Physiology of Halophytes: Current Standing. Front. Plant Sci. 2018, 9, 1954. [Google Scholar] [CrossRef] [PubMed]
- Caperta, A.D.; Róis, A.S.; Teixeira, G.; Garcia-Caparros, P.; Flowers, T.J. Secretory Structures in Plants: Lessons from the Plumbaginaceae on Their Origin, Evolution and Roles in Stress Tolerance. Plant Cell Environ. 2020, 43, 2912–2931. [Google Scholar] [CrossRef]
- Rahman, M.M.; Mostofa, M.G.; Keya, S.S.; Siddiqui, M.N.; Ansary, M.M.U.; Das, A.K.; Rahman, M.A.; Tran, L.S.P. Adaptive Mechanisms of Halophytes and Their Potential in Improving Salinity Tolerance in Plants. Int. J. Mol. Sci. 2021, 22, 10733. [Google Scholar] [CrossRef]
- Saeed, S.; Gul, B.; Ajmal Khan, M. Comparative Effects of NaCl and Sea Salt on Seed Germination of Arthrocnemum indicum. Pak. J. Bot. 2011, 43, 1091–1103. [Google Scholar]
- Evelin, H.; Devi, T.S.; Gupta, S.; Kapoor, R. Mitigation of Salinity Stress in Plants by Arbuscular Mycorrhizal Symbiosis: Current Understanding and New Challenges. Front. Plant Sci. 2019, 10, 470. [Google Scholar] [CrossRef]
- Dastogeer, K.M.G.; Zahan, M.I.; Tahjib-Ul-Arif, M.; Akter, M.A.; Okazaki, S. Plant Salinity Tolerance Conferred by Arbuscular Mycorrhizal Fungi and Associated Mechanisms: A Meta-Analysis. Front. Plant Sci. 2020, 11, 588550. [Google Scholar] [CrossRef]
- Tian, C.Y.; Feng, G.; Li, X.L.; Zhang, F.S. Different Effects of Arbuscular Mycorrhizal Fungal Isolates from Saline or Non-Saline Soil on Salinity Tolerance of Plants. Appl. Soil Ecol. 2004, 26, 143–148. [Google Scholar] [CrossRef]
- Wani, P.A.; Khan, M.S.; Zaidi, A. Synergistic Effects of the Inoculation with Nitrogen-Fixing and Phosphate-Solubilizing Rhizobacteria on the Performance of Field-Grown Chickpea. J. Plant Nutr. Soil Sci. 2007, 170, 283–287. [Google Scholar] [CrossRef]
- Porcel, R.; Aroca, R.; Ruiz-Lozano, J.M. Salinity Stress Alleviation Using Arbuscular Mycorrhizal Fungi. A Review. Agron. Sustain. Dev. 2012, 32, 181–200. [Google Scholar] [CrossRef]
- Navarro-Torre, S.; Carro, L.; Rodríguez-Llorente, I.D.; Pajuelo, E.; Caviedes, M.Á.; Igual, J.M.; Klenk, H.P.; Montero-Calasanz, M.D.C. Halomonas radicis sp. Nov., Isolated from Arthrocnemum macrostachyum Growing in the Odiel Marshes (Spain) and Emended Descriptions of Halomonas xinjiangensis and Halomonas zincidurans. Int. J. Syst. Evol. Microbiol. 2020, 70, 220–227. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–Microbiome Interactions: From Community Assembly to Plant Health. Nat. Rev. Microbiol. 2020, 18, 607–621. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.A.; Sahile, A.A.; Jan, R.; Asaf, S.; Hamayun, M.; Imran, M.; Adhikari, A.; Kang, S.M.; Kim, K.M.; Lee, I.J. Halotolerant Bacteria Mitigate the Effects of Salinity Stress on Soybean Growth by Regulating Secondary Metabolites and Molecular Responses. BMC Plant Biol. 2021, 21, 176. [Google Scholar] [CrossRef] [PubMed]
- Shukla, P.S.; Agarwal, P.K.; Jha, B. Improved Salinity Tolerance of Arachis hypogaea (L.) by the Interaction of Halotolerant Plant-Growth-Promoting Rhizobacteria. J. Plant Growth Regul. 2012, 31, 195–206. [Google Scholar] [CrossRef]
- Sharma, S.; Kulkarni, J.; Jha, B. Halotolerant Rhizobacteria Promote Growth and Enhance Salinity Tolerance in Peanut. Front. Microbiol. 2016, 7, 1600. [Google Scholar] [CrossRef]
- Santos, J.; Al-Azzawi, M.; Aronson, J.; Flowers, T.J. EHALOPH a Database of Salt-Tolerant Plants: Helping Put Halophytes to Work. Plant Cell Physiol. 2016, 57, e10. [Google Scholar] [CrossRef]
- Lledó, M.D.; Karis, P.O.; Crespo, M.B.; Fay, M.F.; Chase, M.W. Endemism and Evolution in Macaronesian and Mediterranean Limonium Taxa. In The Biology of Island Floras; Cambridge University Press: Cambridge, UK, 2011; pp. 325–337. [Google Scholar] [CrossRef]
- Morgan, E.; Funnell, K. Limonium. In Handbook of Plant Breeding; Springer: Cham, Switzerland, 2018; pp. 513–527. [Google Scholar] [CrossRef]
- Caperta, A.D.; Castro, S.; Loureiro, J.; Róis, A.S.; Conceição, S.; Costa, J.; Rhazi, L.; Espírito Santo, D.; Arsénio, P. Biogeographical, Ecological and Ploidy Variation in Related Asexual and Sexual Limonium taxa (Plumbaginaceae). Bot. J. Linn. Soc. 2017, 183, 75–93. [Google Scholar] [CrossRef]
- Cortinhas, A.; Caperta, A.D.; Teixeira, G.; Carvalho, L.; Abreu, M.M. Harnessing Sediments of Coastal Aquaculture Ponds through Technosols Construction for Halophyte Cultivation Using Saline Water Irrigation. J. Environ. Manag. 2020, 261, 109907. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, M.J.; Monteiro, I.; Castañeda-Loaiza, V.; Placines, C.; Conceição Oliveira, M.; Reis, C.; Caperta, A.D.; Soares, F.; Pousão-Ferreira, P.; Pereira, C.; et al. Growth Performance, in Vitro Antioxidant Properties and Chemical Composition of the Halophyte Limonium algarvense Erben Are Strongly Influenced by the Irrigation Salinity. Ind. Crops Prod. 2020, 143, 111930. [Google Scholar] [CrossRef]
- Caperta, A.; Carapeto, A. Limonium daveaui: Ficha de Avaliação Do Risco de Extinção. In Lista Vermelha da Flora Vascular de Portugal Continental; Sociedade Portuguesa de Botânica, Associação Portuguesa de Ciência da Vegetação—PHYTOS and Instituto da Conservação da Natureza e das Florestas: Lisboa, Portugal, 2020; Available online: https://listavermelha-flora.pt/wp-content/uploads/2020/10/Lista_Vermelha_Flora_Vascular_Portugal_Continental_2020_versao_digital.pdf (accessed on 3 June 2023).
- Cortinhas, A.; Ferreira, T.C.; Abreu, M.M.; Caperta, A.D. Conservation of a Critically Endangered Endemic Halophyte of West Portugal: A Microcosm Assay to Assess the Potential of Soil Technology for Species Reintroduction. Front. Ecol. Evol. 2021, 9, 604509. [Google Scholar] [CrossRef]
- Negacz, K.; Malek, Ž.; de Vos, A.; Vellinga, P. Saline Soils Worldwide: Identifying the Most Promising Areas for Saline Agriculture. J. Arid. Environ. 2022, 203, 104775. [Google Scholar] [CrossRef]
- Rozema, J.; Schat, H. Salt Tolerance of Halophytes, Research Questions Reviewed in the Perspective of Saline Agriculture. Environ. Exp. Bot. 2013, 92, 83–95. [Google Scholar] [CrossRef]
- Rengasamy, P. Soil Chemistry Factors Confounding Crop Salinity Tolerance—A Review. Agronomy 2016, 6, 53. [Google Scholar] [CrossRef]
- Santos, E.S.; Abreu, M.M.; Macías, F.; de Varennes, A. Chemical Quality of Leachates and Enzymatic Activities in Technosols with Gossan and Sulfide Wastes from the São Domingos Mine. J. Soils Sediments 2016, 16, 1366–1382. [Google Scholar] [CrossRef]
- Macías, F. Recuperación de Suelos Degradados, Reutilización de Residuos y Secuestro de Carbono. Una Alternativa Integral de Mejora de La Calidad Ambiental. Recur. Rurais 2004, 1, 49–56. [Google Scholar]
- Macías García, F.; Marta Camps Arbestain, F.; Vázquez, F.M. Utilización de Tecnosoles Derivados de Residuos En Procesos de Restauración de Suelos en La Mina de Touro. In Proceedings of the Minería Sostenible: Conferencia internacional 09, Santiago de Compostela, Spain, 15–18 April 2009; Cámara Oficial Mineira de Galicia: La Coruña, Spain, 2009; pp. 651–662.
- Moreno-Barriga, F.; Díaz, V.; Acosta, J.A.; Muñoz, M.Á.; Faz, Á.; Zornoza, R. Organic Matter Dynamics, Soil Aggregation and Microbial Biomass and Activity in Technosols Created with Metalliferous Mine Residues, Biochar and Marble Waste. Geoderma 2017, 301, 19–29. [Google Scholar] [CrossRef]
- Arán, D.; Santos, E.S.; Abreu, M.M.; Antelo, J.; Macías, F. Use of Combined Tools for Effectiveness Evaluation of Tailings Rehabilitated with Designed Technosol. Environ. Geochem. Health 2022, 44, 1857–1873. [Google Scholar] [CrossRef]
- Santos, E.S.; Abreu, M.M.; Macías, F.; de Varennes, A. Improvement of Chemical and Biological Properties of Gossan Mine Wastes Following Application of Amendments and Growth of Cistus ladanifer L. J. Geochem. Explor. 2014, 147, 173–181. [Google Scholar] [CrossRef]
- Hafeez, F.; Spor, A.; Breuil, M.C.; Schwartz, C.; Martin-Laurent, F.; Philippot, L. Distribution of Bacteria and Nitrogen-Cycling Microbial Communities along Constructed Technosol Depth-Profiles. J. Hazard. Mater. 2012, 231–232, 88–97. [Google Scholar] [CrossRef] [PubMed]
- Villenave, C.; Séré, G.; Schwartz, C.; Watteau, F.; Jimenez, A.; Cortet, J. Rapid Changes in Soil Nematodes in the First Years after Technosol Construction for the Remediation of an Industrial Wasteland. Eurasian Soil Sci. 2018, 51, 1266–1273. [Google Scholar] [CrossRef]
- Guerreiro, M.; Fortunato, A.B.; Freire, P.; Rilo, A.; Taborda, R.; Freitas, M.C.; Andrade, C.; Silva, T.; Rodrigues, M.; Bertin, X.; et al. Evolution of the Hydrodynamics of the Tagus Estuary (Portugal) in the 21st Century. J. Integr. Coast. Zone Manag. 2015, 15, 65–80. [Google Scholar] [CrossRef]
- Róis, A.S.; Teixeira, G.; Sharbel, T.F.; Fuchs, J.; Martins, S.; Espírito-Santo, D.; Caperta, A.D. Male Fertility versus Sterility, Cytotype, and DNA Quantitative Variation in Seed Production in Diploid and Tetraploid Sea Lavenders (Limonium sp., Plumbaginaceae) Reveal Diversity in Reproduction Modes. Sex. Plant Reprod. 2012, 25, 305–318. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The Water-Culture Method for Growing Plants without Soil. In Circular; California Agricultural Experiment Station: Berkeley, CA, USA, 1950; Volume 347. [Google Scholar]
- Springer, U.; Klee, J. Prüfung der Leistungsfähigkeit von Einigen Wichtigeren Verfahren zur Bestimmung des Kohlenstoffs Mittels Chromschwefelsäure Sowie Vorschlag Einer Neuen Schnellmethode. Z. Pflanz. Düngung Bodenkd. 1954, 64, 1–26. [Google Scholar] [CrossRef]
- Lakanen, E.; Ervio, R. A Comparison of Eight Extractants for the Determination of Plant Available Micronutrients in Soils. Acta Agral. Fenn. 1971, 123, 223–232. [Google Scholar]
- Tabatabai, M.A. Soil Enzymes. In Methods of Soil Analysis. Part 2. Microbiological and Biochemical Properties; Weaver, R.W., Angle, J.S., Bottomley, P.S., Eds.; Soil Science Society of America: Madison, WI, USA, 1994; pp. 775–833. ISBN 9780891188650. [Google Scholar]
- Eivazi, F.; Tabatabai, M.A. Glucosidases and Galactosidases in Soils. Soil Biol. Biochem. 1988, 20, 601–606. [Google Scholar] [CrossRef]
- Kandeler, E.; Gerber, H. Short-Term Assay of Soil Urease Activity Using Colorimetric Determination of Ammonium. Biol. Fertil. Soils 1988, 6, 68–72. [Google Scholar] [CrossRef]
- Hope, C.F.A.; Burns, R.G. Activity, Origins and Location of Cellulases in a Silt Loam Soil. Biol. Fertil. Soils 1987, 5, 164–170. [Google Scholar] [CrossRef]
- Murphy, J.; Riley, J. A modified single Solution for the Determination of Phosphate in Natural Waters. Anal. Chim. Acta 1962, 27, 31. [Google Scholar] [CrossRef]
- Porter, W. The “most Probable Number” Method for Enumerating Infective Propagules of Vesicular Arbuscular Mycorrhizal Fungi in Soil. Aust. J. Soil Res. 1979, 17, 515. [Google Scholar] [CrossRef]
- Powell, C.L. Mycorrhizal Infectivity of Eroded Soils. Soil Biol. Biochem. 1980, 12, 247–250. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved Procedures for Clearing Roots and Staining Parasitic and Vesicular-Arbuscular Mycorrhizal Fungi for Rapid Assessment of Infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Koske, R.E.; Gemma, J.N. A Modified Procedure for Staining Roots to Detect VA Mycorrhizas. Mycol. Res. 1989, 92, 486–488. [Google Scholar] [CrossRef]
- Jarvis, B.; Wilrich, C.; Wilrich, P.-T. Reconsideration of the Derivation of Most Probable Numbers, Their Standard Deviations, Confidence Bounds and Rarity Values. J. Appl. Microbiol. 2010, 109, 1660–1667. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Torre, S.; Barcia-Piedras, J.M.; Mateos-Naranjo, E.; Redondo-Gómez, S.; Camacho, M.; Caviedes, M.A.; Pajuelo, E.; Rodríguez-Llorente, I.D. Assessing the Role of Endophytic Bacteria in the Halophyte Arthrocnemum Macrostachyum Salt Tolerance. Plant Biol. 2016, 19, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Torre, S.; Carro, L.; Rodríguez-Llorente, I.D.; Pajuelo, E.; Caviedes, M.Á.; Igual, J.M.; Klenk, H.P.; Montero-Calasanz, M.D.C. Pseudoalteromonas rhizosphaerae sp. Nov., a Novel Plant Growthpromoting Bacterium with Potential Use in Phytoremediation. Int. J. Syst. Evol. Microbiol. 2020, 70, 3287–3294. [Google Scholar] [CrossRef]
- Navarro-Torre, S.; Barcia-Piedras, J.M.; Caviedes, M.A.; Pajuelo, E.; Redondo-Gómez, S.; Rodríguez-Llorente, I.D.; Mateos-Naranjo, E. Bioaugmentation with Bacteria Selected from the Microbiome Enhances Arthrocnemum Macrostachyum Metal Accumulation and Tolerance. Mar. Pollut. Bull. 2017, 117, 340–347. [Google Scholar] [CrossRef]
- Zhang, C.; Fountain, D.W.; Morgan, E.R. In Vitro Germination of the Trinucleate Pollen of Limonium perezii. Grana 1997, 36, 284–288. [Google Scholar] [CrossRef]
- Leogrande, R.; Vitti, C. Use of Organic Amendments to Reclaim Saline and Sodic Soils: A Review. Arid. Land Res. Manag. 2018, 33, 1498038. [Google Scholar] [CrossRef]
- Carvalho, L.C.; Santos, E.S.; Saraiva, J.A.; Magalhães, M.C.F.; Macías, F.; Abreu, M.M. The Potential of Cistus salviifolius L. to Phytostabilize Gossan Mine Wastes Amended with Ash and Organic Residues. Plants 2022, 11, 588. [Google Scholar] [CrossRef] [PubMed]
- Betancur-Corredor, B.; Loaiza-Usuga, J.C.; Denich, M.; Borgemeister, C. Changes of Technosol Properties and Vegetation Structure along a Chronosequence of Dredged Sediment Deposition in Areas with Alluvial Gold Mining in Colombia. J. Soils Sediments 2020, 20, 2377–2394. [Google Scholar] [CrossRef]
- Adetunji, A.T.; Lewu, F.B.; Mulidzi, R.; Ncube, B. The Biological Activities of β-Glucosidase, Phosphatase and Urease as Soil Quality Indicators: A Review. J. Soil Sci. Plant Nutr. 2017, 17, 794–807. [Google Scholar] [CrossRef]
- Wolińska, A.; Stępniewska, Z.; Wolińska, A.; Stępniewska, Z. Dehydrogenase Activity in the Soil Environment. In Dehydrogenases; IntechOpen: London, UK, 2012. [Google Scholar] [CrossRef]
- Santos, E.S.; Arán, D.; Abreu, M.M.; de Varennes, A. Engineered Soils Using Amendments for In Situ Rehabilitation of Mine Lands. In Bio-Geotechnologies for Mine Site Rehabilitation; Elsevier: Amsterdam, The Netherlands, 2016; pp. 131–146. [Google Scholar] [CrossRef]
- Tejada, M.; Garcia, C.; Gonzalez, J.L.; Hernandez, M.T. Use of Organic Amendment as a Strategy for Saline Soil Remediation: Influence on the Physical, Chemical and Biological Properties of Soil. Soil Biol. Biochem. 2006, 38, 1413–1421. [Google Scholar] [CrossRef]
- Wichern, J.; Wichern, F.; Joergensen, R.G. Impact of Salinity on Soil Microbial Communities and the Decomposition of Maize in Acidic Soils. Geoderma 2006, 137, 100–108. [Google Scholar] [CrossRef]
- Xie, X.; Pu, L.; Wang, Q.; Zhu, M.; Xu, Y.; Zhang, M. Response of Soil Physicochemical Properties and Enzyme Activities to Long-Term Reclamation of Coastal Saline Soil, Eastern China. Sci. Total Environ. 2017, 607–608, 1419–1427. [Google Scholar] [CrossRef]
- Boyrahmadi, M.; Raiesi, F. Plant Roots and Species Moderate the Salinity Effect on Microbial Respiration, Biomass, and Enzyme Activities in a Sandy Clay Soil. Biol. Fertil. Soils 2018, 54, 509–521. [Google Scholar] [CrossRef]
- Sritongon, N.; Sarin, P.; Theerakulpisut, P.; Riddech, N. The Effect of Salinity on Soil Chemical Characteristics, Enzyme Activity and Bacterial Community Composition in Rice Rhizospheres in Northeastern Thailand. Sci. Rep. 2022, 12, 20360. [Google Scholar] [CrossRef]
- Kang, Y.H.; Liu, S.H.; Wan, S.Q.; Wang, R.S. Assessment of Soil Enzyme Activities of Saline-Sodic Soil under Drip Irrigation in the Songnen Plain. Paddy Water Environ. 2013, 11, 87–95. [Google Scholar] [CrossRef]
- Singh, K. Microbial and Enzyme Activities of Saline and Sodic Soils. Land Degrad. Dev. 2016, 27, 706–718. [Google Scholar] [CrossRef]
- Weil, R.R.; Brady, N.C. The Nature and Properties of Soils, 15th ed.; Weil, R.R., Brady, N.C., Eds.; Pearson Education Limited: Edinburgh, UK, 2017; ISBN 0133254488. [Google Scholar]
- de Souza, R.; Ambrosini, A.; Passaglia, L.M.P. Plant Growth-Promoting Bacteria as Inoculants in Agricultural. Genet. Mol. Biol. 2015, 38, 401. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Torre, S.; Garcia-Caparrós, P.; Nogales, A.; Abreu, M.M.; Santos, E.; Cortinhas, A.L.; Caperta, A.D. Sustainable Agricultural Management of Saline Soils in Arid and Semi-Arid Mediterranean Regions through Halophytes, Microbial and Soil-Based Technologies. Environ. Exp. Bot. 2023, 212, 105397. [Google Scholar] [CrossRef]
- Tang, A.; Haruna, A.O.; Majid, N.M.A.; Jalloh, M.B. Potential PGPR Properties of Cellulolytic, Nitrogen-Fixing, Phosphate-Solubilizing Bacteria in Rehabilitated Tropical Forest Soil. Microorganisms 2020, 8, 442. [Google Scholar] [CrossRef] [PubMed]
- Camprubi, A.; Abril, M.; Estaun, V.; Calvet, C. Contribution of Arbuscular Mycorrhizal Symbiosis to the Survival of Psammophilic Plants after Sea Water Flooding. Plant Soil 2012, 351, 97–105. [Google Scholar] [CrossRef]
- Navarro-Torre, S.; Mateos-Naranjo, E.; Caviedes, M.A.; Pajuelo, E.; Rodríguez-Llorente, I.D. Isolation of Plant-Growth-Promoting and Metal-Resistant Cultivable Bacteria from Arthrocnemum Macrostachyum in the Odiel Marshes with Potential Use in Phytoremediation. Mar. Pollut. Bull. 2016, 110, 133–142. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.Q.; Lu, X.B.; Li, Z.H.; Tian, C.Y.; Song, J. The Role of Root-Associated Microbes in Growth Stimulation of Plants under Saline Conditions. Land Degrad. Dev. 2021, 32, 3471–3486. [Google Scholar] [CrossRef]
- Ochoa-Hueso, R.; Pérez-Corona, M.E.; Manrique, E. Impacts of Simulated N Deposition on Plants and Mycorrhizae from Spanish Semiarid Mediterranean Shrublands. Ecosystems 2013, 16, 838–851. [Google Scholar] [CrossRef]
- Sheikh-Assadi, M.; Khandan-Mirkohi, A.; Alemardan, A.; Moreno-Jiménez, E. Mycorrhizal Limonium sinuatum (L.) Mill. Enhances Accumulation of Lead and Cadmium. Int. J. Phytoremed. 2015, 17, 556–562. [Google Scholar] [CrossRef]
- Qin, S.; Zhang, Y.J.; Yuan, B.; Xu, P.Y.; Xing, K.; Wang, J.; Jiang, J.H. Isolation of ACC Deaminase-Producing Habitat-Adapted Symbiotic Bacteria Associated with Halophyte Limonium sinense (Girard) Kuntze and Evaluating Their Plant Growth-Promoting Activity under Salt Stress. Plant Soil 2014, 374, 753–766. [Google Scholar] [CrossRef]
- Qin, S.; Feng, W.W.; Wang, T.T.; Ding, P.; Xing, K.; Jiang, J.H. Plant Growth-Promoting Effect and Genomic Analysis of the Beneficial Endophyte streptomyces sp. KLBMP 5084 Isolated from Halophyte Limonium sinense. Plant Soil 2017, 416, 117–132. [Google Scholar] [CrossRef]
- Qin, S.; Feng, W.-W.; Zhang, Y.-J.; Wang, T.-T.; Xiong, Y.-W.; Xing, K. Diversity of Bacterial Microbiota of Coastal Halophyte Limonium sinense and Amelioration of Salinity Stressdamage. Appl. Environ. Microbiol. 2018, 84, e01533-18. [Google Scholar] [PubMed]
- Feng, W.W.; Wang, T.T.; Bai, J.L.; Ding, P.; Xing, K.; Jiang, J.H.; Peng, X.; Qin, S. Glutamicibacter halophytocola sp. Nov., an Endophytic Actinomycete Isolated from the Roots of a Coastal Halophyte, Limonium sinense. Int. J. Syst. Evol. Microbiol. 2017, 67, 1120–1125. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.T.; Ding, P.; Chen, P.; Xing, K.; Bai, J.L.; Wan, W.; Jiang, J.H.; Qin, S. Complete Genome Sequence of Endophyte Bacillus flexus KLBMP 4941 Reveals Its Plant Growth Promotion Mechanism and Genetic Basis for Salt Tolerance. J. Biotechnol. 2017, 260, 38–41. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Li, E.; Liu, C.; Jousset, A.; Salles, J.F. Functionality of Root-Associated Bacteria along a Salt Marsh Primary Succession. Front. Microbiol. 2017, 8, 2102. [Google Scholar] [CrossRef]
- Xiong, Y.W.; Li, X.W.; Wang, T.T.; Gong, Y.; Zhang, C.M.; Xing, K.; Qin, S. Root Exudates-Driven Rhizosphere Recruitment of the Plant Growth-Promoting Rhizobacterium Bacillus flexus KLBMP 4941 and Its Growth-Promoting Effect on the Coastal Halophyte Limonium sinense under Salt Stress. Ecotoxicol. Environ. Saf. 2020, 194, 110374. [Google Scholar] [CrossRef]
- Aly, A.H.; Debbab, A.; Clements, C.; Edrada-Ebel, R.; Orlikova, B.; Diederich, M.; Wray, V.; Lin, W.; Proksch, P. NF Kappa B Inhibitors and Antitrypanosomal Metabolites from Endophytic Fungus penicillium sp. Isolated from Limonium tubiflorum. Bioorg. Med. Chem. 2011, 19, 414–421. [Google Scholar] [CrossRef]
- Khalmuratova, I.; Kim, H.; Nam, Y.J.; Oh, Y.; Jeong, M.J.; Choi, H.R.; You, Y.H.; Choo, Y.S.; Lee, I.J.; Shin, J.H.; et al. Diversity and Plant Growth Promoting Capacity of Endophytic Fungi Associated with Halophytic Plants from the West Coast of Korea. Mycobiology 2015, 43, 373–383. [Google Scholar] [CrossRef]
- Khalmuratova, I.; Choi, D.H.; Woo, J.R.; Jeong, M.J.; Oh, Y.; Kim, Y.G.; Lee, I.J.; Choo, Y.S.; Kim, J.G. Diversity and Plant Growth-Promoting Effects of Fungal Endophytes Isolated from Salt-Tolerant Plants. J. Microbiol. Biotechnol. 2020, 30, 1680–1687. [Google Scholar] [CrossRef]
- Khalmuratova, I.; Choi, D.H.; Kim, J.G.; Lee, I.S. Endophytic Fungi of Salt-Tolerant Plants: Diversity and Ability to Promote Plant Growth. J. Microbiol. Biotechnol. 2021, 31, 1526–1532. [Google Scholar] [CrossRef]
- Mandyam, K.G.; Jumpponen, A. Mutualism-Parasitism Paradigm Synthesized from Results of Root-Endophyte Models. Front. Microbiol. 2014, 5, 776. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, R.J.; White, J.F.; Arnold, A.E.; Redman, R.S. Fungal Endophytes: Diversity and Functional Roles. New Phytol. 2009, 182, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Malicka, M.; Magurno, F.; Piotrowska-Seget, Z. Plant Association with Dark Septate Endophytes: When the Going Gets Tough (and Stressful), the Tough Fungi Get Going. Chemosphere 2022, 302, 134830. [Google Scholar] [CrossRef]
- Rodriguez, R.J.; Henson, J.; Van Volkenburgh, E.; Hoy, M.; Wright, L.; Beckwith, F.; Kim, Y.O.; Redman, R.S. Stress Tolerance in Plants via Habitat-Adapted Symbiosis. ISME J. 2008, 2, 404–416. [Google Scholar] [CrossRef]
- Santos, M.; Cesanelli, I.; Diánez, F.; Sánchez-Montesinos, B.; Moreno-Gavíra, A. Advances in the Role of Dark Septate Endophytes in the Plant Resistance to Abiotic and Biotic Stresses. J. Fungi 2021, 7, 939. [Google Scholar] [CrossRef]
- Menge, J.; Davis, R. Mycorrhizal Fungi Increase Growth and Reduce Transplant Injury in Avocado. Calif. Agric. 1978, 32, 6–7. [Google Scholar]
- Subhan, S.; Sharmila, P.; Pardha Saradhi, P. Glomus Fasciculatum Alleviates Transplantation Shock of Micropropagated Sesbania Sesban. Plant Cell Rep. 1998, 17, 268–272. [Google Scholar] [CrossRef] [PubMed]
- Krishna, H.; Singh, S.K.; Minakshi; Patel, V.B.; Khawale, R.N.; Deshmukh, P.S.; Jindal, P.C. Arbuscular-Mycorrhizal Fungi Alleviate Transplantation Shock in Micropropagated Grapevine (Vitis vinifera L.). J. Hortic. Sci. Biotechnol. 2006, 81, 259–263. [Google Scholar] [CrossRef]
- Chen, H.; Jiang, J.G. Osmotic Adjustment and Plant Adaptation to Environmental Changes Related to Drought and Salinity. Environ. Rev. 2010, 18, 309–319. [Google Scholar] [CrossRef]
- Hameed, A.; Dilfuza, E.; Abd-Allah, E.F.; Hashem, A.; Kumar, A.; Ahmad, P. Salinity Stress and Arbuscular Mycorrhizal Symbiosis in Plants. In Beyond the Dance Floor; Springer: New York, NY, USA, 2014; pp. 139–159. ISBN 9781461494669. [Google Scholar]
- Pan, J.; Peng, F.; Tedeschi, A.; Xue, X.; Wang, T.; Liao, J.; Zhang, W.; Huang, C. Do Halophytes and Glycophytes Differ in Their Interactions with Arbuscular Mycorrhizal Fungi under Salt Stress? A Meta-Analysis. Bot. Stud. 2020, 61, 13. [Google Scholar] [CrossRef] [PubMed]
- Sembiring, H.; Raun, W.R.; Johnson, G.V.; Stone, M.L.; Solie, J.B.; Phillips, S.B. Detection of Nitrogen and Phosphorus Nutrient Status in Winter Wheat Using Spectral Radiance. J. Plant Nutr. 1998, 21, 1207–1233. [Google Scholar] [CrossRef]
- Jakobsen, I. Transport of Phosphorus and Carbon in Arbuscular Mycorrhizas. In Mycorrhiza; Springer: Berlin/Heidelberg, Germany, 2016; pp. 305–332. [Google Scholar] [CrossRef]
- Ryan, M.H.; Graham, J.H. Is There a Role for Arbuscular Mycorrhizal Fungi in Production Agriculture? Plant Soil 2002, 244, 263–271. [Google Scholar] [CrossRef]
- Mortimer, P.E.; Archer, E.; Valentine, A.J. Mycorrhizal C Costs and Nutritional Benefits in Developing Grapevines. Mycorrhiza 2005, 15, 159–165. [Google Scholar] [CrossRef]
- Gavito, M.E.; Jakobsen, I.; Mikkelsen, T.N.; Mora, F. Direct Evidence for Modulation of Photosynthesis by an Arbuscular Mycorrhiza-induced Carbon Sink Strength. New Phytol. 2019, 223, 896–907. [Google Scholar] [CrossRef]
- Wang, L.; Chen, X.; Wang, S.; Du, Y.; Zhang, D.; Tang, Z. Effects of Arbuscular Mycorrhizal Symbiosis on the Growth and Reproduction of Cherry Tomato Can Be Persistent to the next Generation. Eur. J. Soil Biol. 2022, 112, 103429. [Google Scholar] [CrossRef]
- Zhu, B.; Gao, T.; Zhang, D.; Ding, K.; Li, C.; Ma, F. Functions of Arbuscular Mycorrhizal Fungi in Horticultural Crops. Sci. Hortic. 2022, 303, 111219. [Google Scholar] [CrossRef]
- Liu, S.; Guo, H.; Xu, J.; Song, Z.; Song, S.; Tang, J.; Chen, X. Arbuscular Mycorrhizal Fungi Differ in Affecting the Flowering of a Host Plant under Two Soil Phosphorus Conditions. J. Plant Ecol. 2018, 11, 623–631. [Google Scholar] [CrossRef]
- Conceição, S.I.R.; Róis, A.S.; Caperta, A.D. Nonreduction via Meiotic Restitution and Pollen Heterogeneity May Explain Residual Male Fertility in Triploid Marine Halophyte Limonium algarvense (Plumbaginaceae). Caryologia 2019, 72, 53–62. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | |
|---|---|
| pH | 6.6 ± 0.09 |
| Electrical conductivity (dS/m) | 39.8 ± 0.12 |
| Chloride (mg/L) | 15,455.9 ± 210.67 |
| Hidrogenocarbonate (mg/L) | 3.50 ± 0.122 |
| P (mg/L) | 0.44 ± 0.003 |
| Mg (mg/L) | 996.59 ± 3.689 |
| Na (mg/L) | 18,995.47 ± 625.884 |
| K (mg/L) | 265.76 ± 0.807 |
| Ca (mg/L) | 318.95 ± 5.149 |
| Fe (mg/L) | 0.13 ± 0.012 |
| Zn (mg/L) | <DL |
| Mn (mg/L) | <DL |
| Cu (mg/L) | <DL |
| Parameters | Fluvisol (FLU) | Amended Fluvisol (TEC) |
|---|---|---|
| pH | 7.9 ± 0.07 a | 7.6 ± 0.02 b |
| CE (dS/m) | 5.6 ± 0.30 a | 6.0 ± 0.26 a |
| Pextract (mg/kg) | 11.1 ± 0.28 b | 369.4 ± 7.67 a |
| Kextract (g/kg) | 0.09 ± 0.00 a | 1.1 ± 0.06 a |
| Total N (g/kg) | 1.7 ± 0.04 b | 2.8 ± 0.11 a |
| Organic C (g/kg) | 20.0 ± 0.95 b | 29.5 ± 1.18 a |
| N-NH4 (mg/kg) | 14.9 ± 2.72 a | 8.2 ± 1.67 a |
| N-NO3 (mg/kg) | 34.7 ± 1.74 b | 96.5 ± 9.70 a |
| Ca (g/kg) | 1.2 ± 0.03 b | 32.6 ± 12.26 a |
| Mg (g/kg) | 1.1 ± 0.01 b | 1.8 ± 0.02 a |
| Na (g/kg) | 5.3 ± 0.19 b | 15.0 ± 2.11 a |
| Fe (mg/kg) | 633.5 ± 10.39 a | 611.6 ± 50.96 a |
| Mn (mg/kg) | 248.5 ± 1.28 a | 226.8 ± 26.92 a |
| Zn (mg/kg) | 8.6 ± 0.13 b | 15.9 ± 1.07 a |
| Cu (mg/kg) | 7.0 ± 0.07 b | 10.2 ± 0.46 a |
| β-glucosidase 1 | 0.17 ± 0.014 b | 0.678 ± 0.086 a |
| Acid phosphatase 2 | 0.27 ± 0.039 b | 1.14 ± 0.090 a |
| Urease 3 | 1.51 ± 0.030 b | 3.75 ± 0.482 a |
| Cellulase 4 | 0.47 ± 0.160 a | 0.71 ±0.110 a |
| Dehydrogenase 5 | 26.03 ± 5.554 b | 255.75 ± 34.696 a |
| Number of mycorrhizal propagules per gram | 2.2 | 0.7 |
| Experimental Treatment | pH | EC (dS/m) | Org C (g/kg) | P (mg/kg) | K (g/kg) | Total N (g/kg) | N-NH4 (mg/kg) | N-NO3 (mg/kg) | Ca (g/kg) | Mg (g/kg) | Na (g/kg) | Fe (mg/kg) | Mn (mg/kg) | Zn (mg/kg) | Cu (mg/kg) | β-Glucosidase 1 | Acid_Phosphatase 2 | Urease 3 | Celulase 4 | Dehydrogenase 5 | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| TEC | Non-inoc | 7.85 ± 0.05 ab | 19.46 ± 0.946 a | 32.1 ± 0.76 a | 349.6 ± 10.52 a | 1.3 ± 0.05 a | 2.8 ± 0.03 a | 8.2 ± 0.38 a | 56.7 ± 20.68 a | 17.1 ± 0.41 b | 3.2 ± 0.11 a | 55.6 ± 1.7 a | 488.8 ± 11.92 a | 211.0 ± 2.67 ab | 17.9 ± 0.22 a | 11.3 ± 0.07 a | 0.27 ± 0.04 ab | 0.64 ± 0.05 ab | 2.01 ± 0.40 a | 0.47 ± 0.03 a | 47.99 ± 3.56 a |
| PGPB | 7.76 ± 0.02 b | 18.16 ± 1.022 ab | 30.1 ± 1.14 a | 349.2 ± 10.09 a | 1.3 ± 0.04 a | 2.7 ± 0.04 a | 5.4 ± 0.79 ab | 51.7 ± 15.12 ab | 17.6 ± 0.76 b | 3.0 ± 0.14 a | 29.3 ± 2.61 b | 479.1 ± 17.97 a | 218.0 ± 3.66 ab | 17.6 ± 0.41 a | 11.5 ± 0.12 a | 0.44 ± 0.10 a | 0.82 ± 0.10 a | 2.35 ± 0.22 a | 0.27 ± 0.03 a | 38.13 ± 6.51 a | |
| AMF | 7.78 ± 0.01 ab | 18.64 ± 0.332 ab | 31.0 ± 1.29 a | 337.5 ± 4.16 a | 1.3 ± 0.01 a | 2.7 ± 0.04 a | 4.4 ± 0.46 b | 39.4 ± 7.03 ab | 15.9 ± 1.14 b | 3.1 ± 0.04 a | 32.4 ± 1.09 b | 489 ± 11.02 a | 225.0 ± 5.50 a | 17.6 ± 0.23 a | 11.7 ± 0.17 a | 0.23 ± 0.04 b | 0.56 ± 0.05 b | 2.13 ± 0.32 a | 0.26 ± 0.07 a | 37.87 ± 4.50 a | |
| AMF + PGPB | 7.89 ± 0.04 a | 16.96 ± 0.404 b | 28.6 ± 1.12 a | 319.0 ± 1.82 b | 1.2 ± 0.02 a | 2.6 ± 0.02 a | 6.3 ± 1.35 ab | 22.7 ± 4.78 b | 43.4 ± 8.99 a | 2.9 ± 0.03 a | 32.3 ± 3.18 b | 347.5 ± 21.56 b | 185.9 ± 13.72 b | 14.9 ± 0.54 b | 10.6 ± 0.09 b | 0.29 ± 0.03 ab | 0.64 ± 0.02 ab | 1.68 ± 0.25 a | 0.24 ± 0.09 a | 32.88 ± 4.50 a | |
| FLU | Non-inoc * | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - |
| PGPB * | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | - | |
| AMF | 7.86 ± 0.02 A | 18.95 ± 0.495 A | 24.4 ± 0.24 A | 11.8 ± 0.13 A | 1.1 ± 0.0 A | 2.0 ± 0.0 A | 2.0 ± 0.17 A | 32.8 ± 2.22 A | 4.4 ± 0.10 A | 3.2 ± 0.04 A | 30.4 ± 0.23 A | 216.0 ± 2.47 A | 272.1 ± 8.75 A | 11.9 ± 0.18 A | 10.5 ± 0.06 A | 0.11 ± 0.0 A | 0.17 ± 0.04 A | 0.90 ± 0.04 A | 0.51 ± 0.26 A | 6.90 ± 0.65 A | |
| AMF + PGPB | 7.91 ± 0.02 A | 15.40 ± 0.318 B | 19.3 ± 0.71 B | 10.3 ± 0.51 A | 1.1 ± 0.02 A | 1.7 ± 0.01 B | 2.8 ± 0.23 A | 24.3 ± 4.67 A | 3.8 ± 0.19 A | 2.5 ± 0.05 B | 38.8 ± 5.63 A | 197.8 ± 8.72 A | 301.1 ± 8.57 A | 9.9 ± 0.07 B | 10.2 ± 0.09 A | 0.16 ± 0.02 A | 0.22 ± 0.01 A | 0.93 ± 0.09 A | 0.29 ± 0.02 A | 6.24 ± 0.96 A | |
| Factor/Effect | pH | EC | Org C | P Ext | K | Total N | N-NH4 | N-NO3 | Ca | Mg | Na | Fe | Mn | Zn | Cu | β-Glucosidase | Acid Phosphatase | Urease | Cellulase | Dehydrogenase |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AMF | 0.38 | 0.22 | 0.36 | 0.02 | 0.22 | 0.02 | 0.24 | 0.10 | 0.05 | 0.42 | <0.01 | 0.01 | 0.43 | 0.01 | 0.08 | 0.16 | 0.07 | 0.46 | 0.18 | 0.21 |
| PGPB | 0.79 | 0.32 | 0.14 | 0.25 | 0.33 | 0.02 | 0.71 | 0.42 | 0.01 | 0.06 | <0.01 | <0.01 | 0.18 | 0.01 | 0.02 | 0.09 | 0.08 | 0.88 | 0.23 | 0.22 |
| Interaction | 0.02 | 0.25 | 0.89 | 0.27 | 0.25 | 0.40 | 0.08 | 0.66 | 0.02 | 0.94 | <0.01 | 0.01 | 0.06 | 0.03 | 0.00 | 0.41 | 0.46 | 0.29 | 0.34 | 0.68 |
| (A) | Factor/Effect | N° Leaves | Shoot Fresh Biomass | Root Fresh Biomass | |||
|---|---|---|---|---|---|---|---|
| AMF | 0.96 | 0.09 | 0.13 | ||||
| PGPB | 0.06 | 0.64 | 0.39 | ||||
| Interaction | 0.34 | 0.85 | 0.28 | ||||
| (B) | N° Leaves | Shoot Fresh Biomass | Root Fresh Biomass | ||||
| Treatment | TEC | FLU | TEC | FLU | TEC | FLU | |
| Non-inoculated | 34 ± 1.4 | - | 11.6 ± 0.23 | - | 2.8 ± 0.99 | - | |
| PGPB | 29 ± 6.4 | - | 11.2 ± 0.87 | - | 4.2 ± 0.69 | - | |
| AMF | 38 ± 1.6 | 9 ± 0 * | 13.8 ± 0.69 | 4.4 ± 0.40 * | 4.7 ± 0.41 | 1.5 ± 0.30 * | |
| AMF + PGPB | 24 ± 4.2 | 8 ± 0.3 * | 13.1 ± 1.41 | 3.6 ± 0.65 * | 4.5 ± 0.05 | 1.4 ± 0.78 * | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nogales, A.; Navarro-Torre, S.; Abreu, M.M.; Santos, E.S.; Cortinhas, A.; Fors, R.; Bailly, M.; Róis, A.S.; Caperta, A.D. Unravelling the Combined Use of Soil and Microbial Technologies to Optimize Cultivation of Halophyte Limonium algarvense (Plumbaginaceae) Using Saline Soils and Water. Soil Syst. 2023, 7, 74. https://doi.org/10.3390/soilsystems7030074
Nogales A, Navarro-Torre S, Abreu MM, Santos ES, Cortinhas A, Fors R, Bailly M, Róis AS, Caperta AD. Unravelling the Combined Use of Soil and Microbial Technologies to Optimize Cultivation of Halophyte Limonium algarvense (Plumbaginaceae) Using Saline Soils and Water. Soil Systems. 2023; 7(3):74. https://doi.org/10.3390/soilsystems7030074
Chicago/Turabian StyleNogales, Amaia, Salvadora Navarro-Torre, Maria Manuela Abreu, Erika S. Santos, Ana Cortinhas, Rosalba Fors, Marion Bailly, Ana Sofia Róis, and Ana Delaunay Caperta. 2023. "Unravelling the Combined Use of Soil and Microbial Technologies to Optimize Cultivation of Halophyte Limonium algarvense (Plumbaginaceae) Using Saline Soils and Water" Soil Systems 7, no. 3: 74. https://doi.org/10.3390/soilsystems7030074