
Arbuscular Mycorrhizal Fungi Improve Growth and Phosphate Nutrition of Acacia seyal (Delile) under Saline Conditions
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Substrate
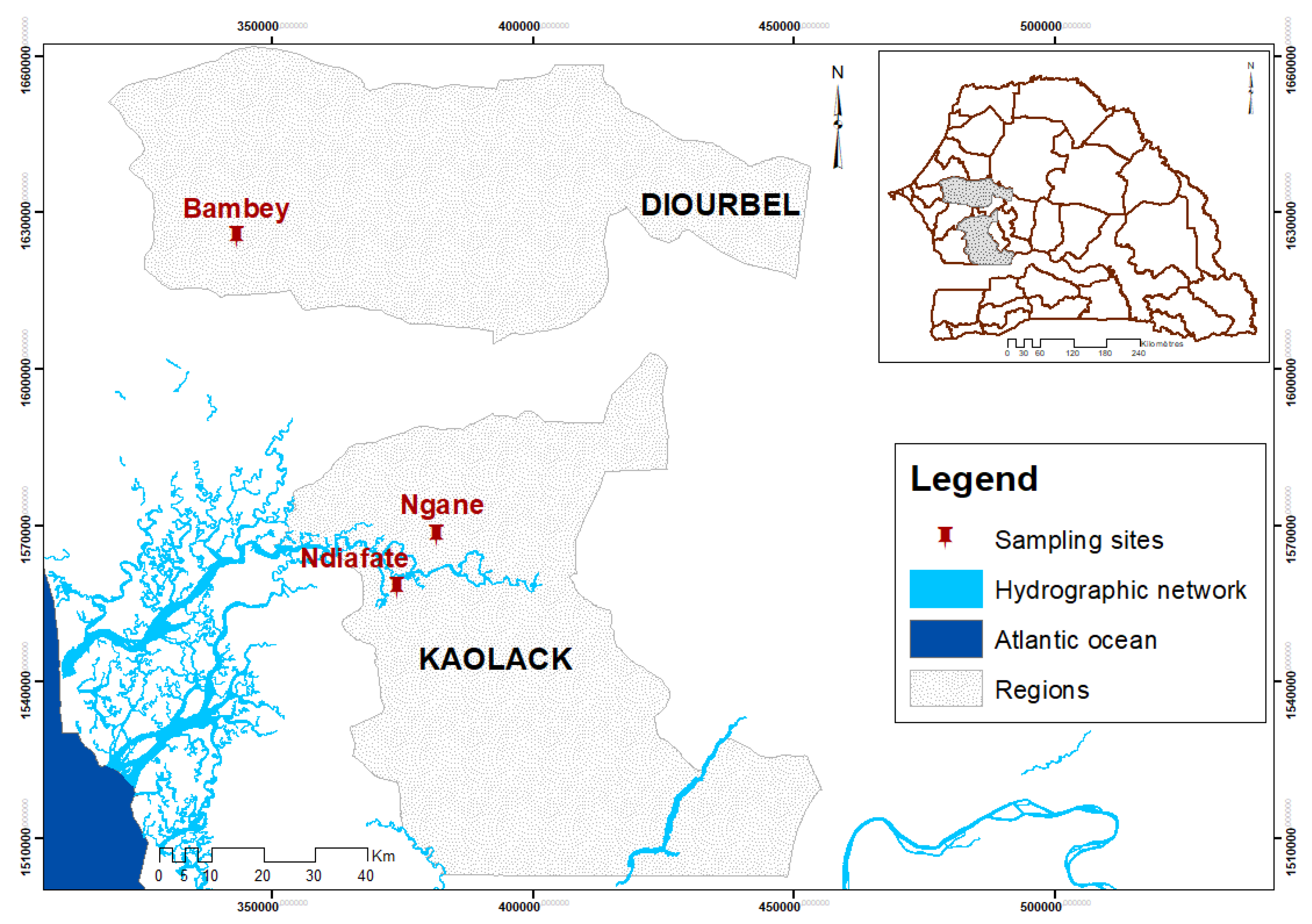
2.2. Sources of Arbuscular Mycorrhizal Fungi
2.3. Soil Physical and Chemical Properties
2.4. Mycorrhizal Inoculum Production
2.5. Experimental Design
2.6. Shoot and Root Height and Dry Weight
2.7. Plant Phosphorus Content
2.8. AMF Root Mycorrhization
2.9. Statistical Analysis
3. Results
3.1. Interaction between NaCl Level and AMF Inoculation on A. seyal Shoot Height and Root Length
3.2. Interaction between NaCl Level and AMF on A. seyal Shoot and Root Biomass
3.3. Interaction between NaCl Level and AMF on A. seyal P Uptake
3.4. Correlation Analysis
3.5. Mycorrhizal Colonisation
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bagyaraj, D.; Sharma, M.P.; Maiti, D. Phosphorus nutrition of crops through arbuscular mycorrhizal fungi. Curr. Sci. 2015, 108, 1288–1293. [Google Scholar]
- Mechri, B.; Manga, A.G.B.; Tekaya, M.; Attia, F.; Cheheb, H.; Ben Meriem, F.; Braham, M.; Boujnah, D.; Hammami, M. Changes in microbial communities and carbohydrate profiles induced by the mycorrhizal fungus (Glomus intraradices) in rhizosphere of olive trees (Olea europaea L.). Appl. Soil Ecol. 2014, 75, 124–133. [Google Scholar] [CrossRef]
- Schachtman, D.P.; Reid, R.J.; Ayling, S.M. Phosphorus uptake by plants: From soil to cell. Plant Physiol. 1998, 116, 447–453. [Google Scholar] [PubMed] [Green Version]
- Nussaume, L.; Kanno, S.; Javot, H.; Marin, E.; Pochon, N.; Ayadi, A.; Nakanishi, T.M.; Thibaud, M.C. Phosphate import in plants: Focus on the PHT1 transporters. Front. Plant Sci. 2011, 2, 83. [Google Scholar] [PubMed] [Green Version]
- Karandashov, V.; Bucher, M. Symbiotic phosphate transport in arbuscular mycorrhizas. Trends Plant Sci. 2005, 10, 22–29. [Google Scholar]
- Smith, S.E.; Jakobsen, I.; Grønlund, M.; Smith, F.A. Roles of arbuscular mycorrhizas in plant phosphorus nutrition: Interactions between pathways of phosphorus uptake in arbuscular mycorrhizal roots have important implications for understanding and manipulating plant phosphorus acquisition. Plant Physiol. 2011, 156, 1050–1057. [Google Scholar]
- Giovannetti, M.; Volpe, V.; Salvioli, A.; Bonfante, P. Fungal and plant tools for the uptake of nutrients in arbuscular mycorrhizas: A molecular view. In Mycorrhizal Mediation of Soil; Elsevier: Amsterdam, The Netherlands, 2017; pp. 107–128. [Google Scholar]
- Barea, J.M. Vesicular-arbuscular mycorrhizae as modifiers of soil fertility. In Advances in Soil Science; Springer: Berlin/Heidelberg, Germany, 1991; pp. 1–40. [Google Scholar]
- Hernández, D.H.Y.; Larsen, J.; González-Rodríguez, A.; Tapia-Torres, Y.; de la Barrera, E.; Eguiarte, L.E.; García-Oliva, F. Cooperation between Sporobolus airoides and associated arbuscular mycorrhizal fungi for phosphorus acquisition under drought conditions in an oligotrophic desert ecosystem. Rhizosphere 2020, 15, 100225. [Google Scholar]
- Mohammed, M.H.; Röhle, H. Gum Talha from Acacia seyal Del. variety seyal in South Kordofan, Sudan. Res. J. For. 2011, 5, 17–26. [Google Scholar]
- Diouf, D.; Fall, D.; Chaintreuil, C.; Ba, A.T.; Dreyfus, B.; Neyra, M.; Ndoye, I.; Moulin, L. Phylogenetic analyses of symbiotic genes and characterization of functional traits of Mesorhizobium spp. strains associated with the promiscuous species Acacia seyal Del. J. Appl. Microbiol. 2010, 108, 818–830. [Google Scholar]
- Diatta, A.A.; Min, D.; Jagadish, S.K. Drought stress responses in non-transgenic and transgenic alfalfa—Current status and future research directions. In Advances in Agronomy; Elsevier: Amsterdam, The Netherlands, 2021; pp. 35–100. [Google Scholar]
- Manga, A.; Diop, A.; Diop, T.A. Functional diversity of mycorrhizal fungi has differential effects on salinity tolerance of Acacia seyal (Del.) seedlings. Open J. Soil Sci. 2017, 7, 315–332. [Google Scholar]
- Sene, G.; Thiao, M.; Manga, A.; Kane, A.; Samba-Mbaye, R.; Mbaye, M.S.; Khasa, D.; Sylla, S. Arbuscular mycorrhizal soil infectivity and spores distribution across plantations of tropical, subtropical and exotic tree species: A case study from the forest reserve of Bandia, Senegal. Afr. J. Ecol. 2012, 50, 218–232. [Google Scholar] [CrossRef]
- Broeckling, C.D.; Broz, A.K.; Bergelson, J.; Manter, D.K.; Vivanco, J.M. Root exudates regulate soil fungal community composition and diversity. Appl. Environ. Microbiol. 2008, 74, 738–744. [Google Scholar] [CrossRef] [Green Version]
- Panwar, J.; Tarafdar, J.C. Arbuscular mycorrhizal fungal dynamics under Mitragyna parvifolia (Roxb.) Korth. in Thar Desert. Appl. Soil Ecol. 2006, 34, 200–208. [Google Scholar] [CrossRef]
- Chebaane, A.; Symanczik, S.; Oehl, F.; Azri, R.; Gargouri, M.; Mäder, P.; Mliki, A.; Fki, L. Arbuscular mycorrhizal fungi associated with Phoenix dactylifera L. grown in Tunisian Sahara oases of different salinity levels. Symbiosis 2020, 81, 173–186. [Google Scholar]
- Augé, R.M.; Toler, H.D.; Saxton, A.M. Arbuscular mycorrhizal symbiosis alters stomatal conductance of host plants more under drought than under amply watered conditions: A meta-analysis. Mycorrhiza 2015, 25, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Bell, L.W.; Harrison, M.T.; Kirkegaard, J.A. Dual-purpose cropping—Capitalising on potential grain crop grazing to enhance mixed-farming profitability. Crop Pasture Sci. 2015, 66, I–IV. [Google Scholar] [CrossRef] [Green Version]
- Harrison, M.T.; Evans, J.R.; Moore, A.D. Using a mathematical framework to examine physiological changes in winter wheat after livestock grazing: 1. Model derivation and coefficient calibration. Field Crop. Res. 2012, 136, 116–126. [Google Scholar] [CrossRef]
- Harrison, M.T.; Evans, J.R.; Moore, A.D. Using a mathematical framework to examine physiological changes in winter wheat after livestock grazing: 2. Model validation and effects of grazing management. Field Crop. Res. 2012, 136, 127–137. [Google Scholar]
- Tigka, T.; Ipsilantis, I. Effects of sand dune, desert and field arbuscular mycorrhizae on lettuce (Lactuca sativa, L.) growth in a natural saline soil. Sci. Hortic. 2020, 264, 109191. [Google Scholar] [CrossRef]
- Boutaj, H.; Meddich, A.; Chakhchar, A.; Wahbi, S.; El Alaoui-Talibi, Z.; Douira, A.; Filali-Maltouf, A.; El Modafar, C. Arbuscular mycorrhizal fungi improve mineral nutrition and tolerance of olive tree to Verticillium wilt. Arch. Phytopathol. Plant Prot. 2020, 53, 673–689. [Google Scholar] [CrossRef]
- Alam, H.; Zamin, M.; Adnan, M.; Ahmad, N.; Nawaz, T.; Saud, S.; Basir, A.; Liu, K.; Harrison, M.T.; Hassan, S.; et al. Evaluating the resistance mechanism of Atriplex leucoclada to salt and water stress; a potential crop for biosaline agriculture. Front. Plant Sci. 2022, 13, 948736. [Google Scholar] [CrossRef]
- Dabin, B. Application des dosages automatiques à l’analyse des sols. Cah. Orstom. Ser. Pedofil 1965, 3, 335–366. [Google Scholar]
- Rayment, G.E.; Higginson, F.R. Australian Laboratory Handbook of Soil and Water Chemical Methods; Inkata Press: Melbourne, Australia, 1992. [Google Scholar]
- Dalpé, Y.; Diop, T.A.; Plenchette, C.; Gueye, M. Glomales species associated with surface and deep rhizosphere of Faidherbia albida in Senegal. Mycorrhiza 2000, 10, 125–129. [Google Scholar] [CrossRef]
- Manga, A.; Dalpé, Y.; Soumaré, A.; Diop, T.A. Diversity of Arbuscular mycorrhizal fungi associated to Acacia seyal (Delile) in semi-arid zone of Senegal. World J. Microbiol. 2017, 3, 120–127. [Google Scholar]
- Gerdemann, J.W.; Nicolson, T.H. Spores of mycorrhizal Endogone species extracted from soil by wet sieving and decanting. Trans. Br. Mycol. Soc. 1963, 46, 235–244. [Google Scholar] [CrossRef]
- Omar, M.B.; Bolland, L.; Heather, W.A. A permanent mounting medium for fungi. Bull. Br. Mycol. Soc. 1979, 13, 31–32. [Google Scholar] [CrossRef]
- Diouf, D.; Samba-Mbaye, R.; Lesueur, D.; Ba, A.T.; Dreyfus, B.; de Lajudie, P.; Neyra, M. Genetic diversity of Acacia seyal Del. rhizobial populations indigenous to Senegalese soils in relation to salinity and pH of the sampling sites. Microb. Ecol. 2007, 54, 553–566. [Google Scholar] [CrossRef]
- Manga, A.; Ndiaye, F.; Sow, H.A.; Diop, T.A. Influence of sodium chloride on the growth, the mineral nutrition and the mycorrhization of Acacia seyal. J. Sci. Technol. 2008, 7, 11–20. [Google Scholar]
- Hewitt, E.J. Sand and water culture methods used in the study of plant nutrition. In London and Reading: Commonwealth Agricultural Bureau, 2nd ed.; The Eastern Press: Norwalk, CT, USA, 1966; 547p. [Google Scholar]
- Van Veldhoven, P.P.; Mannaerts, G.P. Inorganic and organic phosphate measurements in the nanomolar range. Anal. Biochem. 1987, 161, 45–48. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.L.; Gianinazzi-Pearson, V. Mesure du taux de mycorhization VA d’un système radiculaire. Recherche de méthode d’estimation ayant une signification fonctionnelle. In Physiological and Genetical Aspects of Mycorrhizae, Proceedings of the 1st European Symposium on Mycorrhizae, Dijon, France, 1–5 July 1985; INRA: Paris, France, 1986. [Google Scholar]
- Grattan, S.R.; Grieve, C.M. Salinitymineral nutrient relations in horticultural crops. Sci. Hortic. 1999, 78, 127–157. [Google Scholar]
- Herman, P. Biodiversity and evolution in mycorrhizae of the desert. In Microbial Endophytes; Marcel Dekker: New York, NY, USA, 2000; pp. 141–160. [Google Scholar]
- Navarro, J.M.; Pérez-Tornero, O.; Morte, A. Alleviation of salt stress in citrus seedlings inoculated with arbuscular mycorrhizal fungi depends on the rootstock salt tolerance. J. Plant Physiol. 2014, 171, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Miransari, M. Arbuscular mycorrhizal fungi and uptake of nutrients. In Symbiotic Endophytes; Springer: Berlin/Heidelberg, Germany, 2013; pp. 253–270. [Google Scholar]
- Nakmee, P.S.; Techapinyawat, S.; Ngamprasit, S. Comparative potentials of native arbuscular mycorrhizal fungi to improve nutrient uptake and biomass of Sorghum bicolor Linn. Agric. Nat. Resour. 2016, 50, 173–178. [Google Scholar] [CrossRef]
- Karagiannidis, N.; Nikolaou, N. Arbuscular mycorrhizal root infection as an important factor of grapevine nutrition status. Multivariate analysis application for evaluation and characterization of the soil and leaf parameters [Vitis vinifera L.-Greece] [1999]. Agrochimica 2001, 43, 151–165. [Google Scholar]
- Feng, G.; Zhang, F.; Li, X.; Tian, C.; Tang, C.; Rengel, Z. Improved tolerance of maize plants to salt stress by arbuscular mycorrhiza is related to higher accumulation of soluble sugars in roots. Mycorrhiza 2002, 12, 185–190. [Google Scholar] [PubMed]
- Ruiz-Lozano, J.M.; Azcon, R.; Gomez, M. Alleviation of salt stress by arbuscular-mycorrhizal Glomus species in Lactuca sativa plants. Physiol. Plant. 1996, 98, 767–772. [Google Scholar] [CrossRef]
- Jakobsen, I.; Abbott, L.K.; Robson, A.D. External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifolium subterraneum L. 1. Spread of hyphae and phosphorus inflow into roots. New Phytol. 1992, 120, 371–380. [Google Scholar]
- Jakobsen, I.; Abbott, L.K.; Robson, A.D. External hyphae of vesicular-arbuscular mycorrhizal fungi associated with Trifolium subterraneum L. 2. Hyphal transport of 32P over defined distances. New Phytol. 1992, 120, 509–516. [Google Scholar]
- Quintero-Ramos, M.; Espinoza-Victoria, D.; Ferrera-Cerrato, R.; Bethlenfalvay, G.J. Fitting plants to soil through mycorrhizal fungi: Mycorrhiza effects on plant growth and soil organic matter. Biol. Fertil. Soils 1993, 15, 103–106. [Google Scholar] [CrossRef]
- Heydari, S.; Pirzad, A. Mycorrhizal fungi and Thiobacillus co-inoculation improve the physiological indices of Lallemantia iberica under salinity stress. Curr. Microbiol. 2020, 77, 2523–2534. [Google Scholar] [CrossRef]
- Lehmann, A.; Veresoglou, S.; Leifheit, E.F.; Rillig, M. Arbuscular mycorrhizal influence on zinc nutrition in crop plants—A meta-analysis. Soil Biol. Biochem. 2014, 69, 123–131. [Google Scholar] [CrossRef]
- Bolan, N.S. A critical review on the role of mycorrhizal fungi in the uptake of phosphorus by plants. Plant Soil 1991, 134, 189–207. [Google Scholar] [CrossRef]
- Tarafdar, J.C.; Marschner, H. Phosphatase activity in the rhizosphere and hyphosphere of VA mycorrhizal wheat supplied with inorganic and organic phosphorus. Soil Biol. Biochem. 1994, 26, 387–395. [Google Scholar] [CrossRef]
- McArthur, D.A.J.; Knowles, N.R. Influence of vesicular-arbuscular mycorrhizal fungi on the response of potato to phosphorus deficiency. Plant Physiol. 1993, 101, 147–160. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khalil, S.; Loynachan, T.E.; Tabatabai, M.A. Mycorrhizal dependency and nutrient uptake by improved and unimproved corn and soybean cultivars. Agron. J. 1994, 86, 949–958. [Google Scholar] [CrossRef]
- Dodd, J.C.; Burton, C.C.; Burns, R.G.; Jeffries, P. Phosphatase activity associated with the roots and the rhizosphere of plants infected with vesicular-arbuscular mycorrhizal fungi. New Phytol. 1987, 107, 163–172. [Google Scholar] [CrossRef]
- Tisserant, E.; Kohler, A.; Dozolme-Seddas, P.; Balestrini, R.; Benabdellah, K.; Colard, A.; Croll, D.; Da Silva, C.; Gomez, S.K.; Koul, R.; et al. The transcriptome of the arbuscular mycorrhizal fungus Glomus intraradices (DAOM 197198) reveals functional tradeoffs in an obligate symbiont. New Phytol. 2012, 193, 755–769. [Google Scholar] [CrossRef]
- Salvioli, A.; Ghignone, S.; Novero, M.; Navazio, L.; Venice, F.; Bagnaresi, P.; Bonfante, P. Symbiosis with an endobacterium increases the fitness of a mycorrhizal fungus, raising its bioenergetic potential. ISME J. 2016, 10, 130–144. [Google Scholar] [CrossRef]
- Volpe, V.; Giovannetti, M.; Sun, X.-G.; Fiorilli, V.; Bonfante, P. The phosphate transporters LjPT4 and MtPT4 mediate early root responses to phosphate status in non mycorrhizal roots. Plant Cell Environ. 2016, 39, 660–671. [Google Scholar] [CrossRef] [Green Version]
- Estrada, B.; Beltrán-Hermoso, M.; Palenzuela, J.; Iwase, K.; Ruiz-Lozano, J.M.; Barea, J.M.; Oehl, F. Diversity of arbuscular mycorrhizal fungi in the rhizosphere of Asteriscus maritimus (L.) Less., a representative plant species in arid and saline Mediterranean ecosystems. J. Arid. Environ. 2013, 97, 170–175. [Google Scholar] [CrossRef]
- Shi, Z.Y.; Zhang, L.Y.; Li, X.L.; Feng, G.; Tian, C.Y.; Christie, P. Diversity of arbuscular mycorrhizal fungi associated with desert ephemerals in plant communities of Junggar Basin, northwest China. Appl. Soil Ecol. 2007, 35, 10–20. [Google Scholar] [CrossRef] [Green Version]
- Moeneclaey, I.; Schelfhout, S.; Vanhellemont, M.; DeCock, E.; Van Coillie, F.; Verheyen, K.; Baeten, L. Species ecological strategy and soil phosphorus supply interactively affect plant biomass and phosphorus concentration. Basic Appl. Ecol. 2022, 62, 1–11. [Google Scholar] [CrossRef]
- Chandra, P.; Singh, A.; Prajapat, K.; Rai, A.K.; Yadav, R.K. Native arbuscular mycorrhizal fungi improve growth, biomass yield, and phosphorus nutrition of sorghum in saline and sodic soils of the semi–arid region. Environ. Exp. Bot. 2022, 201, 104982. [Google Scholar] [CrossRef]
- Tian, C.Y.; Feng, G.; Li, X.L.; Zhang, F.S. Different effects of arbuscular mycorrhizal fungal isolates from saline or non-saline soil on salinity tolerance of plants. Appl. Soil Ecol. 2004, 26, 143–148. [Google Scholar] [CrossRef]
- Elgharably, A.; Nafady, N.A. Inoculation with Arbuscular mycorrhizae, Penicillium funiculosum and Fusarium oxysporum enhanced wheat growth and nutrient uptake in the saline soil. Rhizosphere 2021, 18, 100345. [Google Scholar] [CrossRef]
- Al-Karaki, G.N.; Hammad, R.; Rusan, M. Response of two tomato cultivars differing in salt tolerance to inoculation with mycorrhizal fungi under salt stress. Mycorrhiza 2001, 11, 43–47. [Google Scholar] [CrossRef]
- Karagiannidis, N.; Nikolaou, N.; Mattheou, A. Influence of three VA-mycorrhiza species on the growth and nutrient uptake of three grapevine rootstocks and one table grape cultivar. Vitis 1995, 34, 85–89. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ngane | Ndiafate | Bambey | |
|---|---|---|---|
| EC mmho/cm | 28.3 | 100.8 | 0.49 |
| pH water | 5.6 | 3.6 | 5.5 |
| pH KCl | 4.9 | 4.2 | 5.2 |
| CEC | 2.42 | 4.26 | 1.78 |
| C/N | 11 | 20 | 10 |
| Assimilable Olsen P2O2 (ppm) | 13.7 | 16.0 | 11.5 |
| Clay | 4.95 | 5.67 | 3.86 |
| Coarse silt 20–50 | 7.88 | 8.57 | 5.52 |
| Coarse sand > 200 | 32.1 | 21.8 | 24.9 |
| Texture | sandy | sandy | sandy |
| AMF Isolates | Intensity (%) | Frequency (%) | ||||
|---|---|---|---|---|---|---|
| NaCl (mM) | ||||||
| 0 | 340 | 680 | 0 | 340 | 680 | |
| Ndiafate | 4.6 ± 0.02 bc | 1.3 ± 0.13 a | 0.6 ± 0.1 a | 42 ± 8.19 c | 32 ± 3.61 abc | 16 ± 3.61 a |
| Ngane | 2.9 ± 0.28 ab | 6.0 ± 0.90 c | 0.7 ± 0.0 a | 32 ± 9.64 abc | 52 ± 5.57 c | 18 ± 3.00 ab |
| Bambey | 2.3 ± 0.69 ab | 1.8 ± 0.47 ab | 1.5 ± 0.2 a | 50 ± 4.58 c | 40 ± 69.54 bc | 34 ± 5.00 abc |
| R. aggregatum | 1.5 ± 0.10 a | 2.1 ± 0.27 ab | 0.7 ± 0.0 a | 32 ± 6.25 abc | 34 ± 5.57 abc | 16 ± 1.00 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Manga, A.G.B.; Ndiaye, M.; Ndiaye, M.A.F.; Sané, S.; Diop, T.A.; Diatta, A.A.; Bassene, C.; Min, D.; Battaglia, M.; Harrison, M.T. Arbuscular Mycorrhizal Fungi Improve Growth and Phosphate Nutrition of Acacia seyal (Delile) under Saline Conditions. Soil Syst. 2022, 6, 79. https://doi.org/10.3390/soilsystems6040079
Manga AGB, Ndiaye M, Ndiaye MAF, Sané S, Diop TA, Diatta AA, Bassene C, Min D, Battaglia M, Harrison MT. Arbuscular Mycorrhizal Fungi Improve Growth and Phosphate Nutrition of Acacia seyal (Delile) under Saline Conditions. Soil Systems. 2022; 6(4):79. https://doi.org/10.3390/soilsystems6040079
Chicago/Turabian StyleManga, Anicet Georges Bruno, Malick Ndiaye, Mame Arama Fall Ndiaye, Seyni Sané, Tahir Abdoulaye Diop, André Amakobo Diatta, César Bassene, Doohong Min, Martin Battaglia, and Matthew Tom Harrison. 2022. "Arbuscular Mycorrhizal Fungi Improve Growth and Phosphate Nutrition of Acacia seyal (Delile) under Saline Conditions" Soil Systems 6, no. 4: 79. https://doi.org/10.3390/soilsystems6040079