
Do Fire Cues Enhance Germination of Soil Seed Stores across an Ecotone of Wet Eucalypt Forest to Cool Temperate Rainforest in the Central Highlands of South-Eastern Australia?

Abstract
:1. Introduction
- Does the soil seed store respond to fire-related germinations cues and do these cues differ among the three forest types? We hypothesize that: (i) fire-related germination cues will be consistent across species shared among the three forest types; (ii) requirement of high-heat will be concentrated in ant-dispersed species with hard seed coats; and (iii) high heat, with or without smoke, will lead to a decline in rainforest-associated species.
- Does the application of karrikinolide increase germinant diversity and abundance and is this dependent on the application of heat? We hypothesize that: (i) any increase in germination to karrikinolide will be independent of heat; and (ii) rainforest-associated species will demonstrate a stronger response to karrikinolide than species associated with more flammable forest types.
- Does the abundance and diversity of soil seed banks differ among forest types? We hypothesize that wet forest and mixed forest will support larger and more diverse soil seed banks than rainforest and in particular, a greater proportion of short-lived, light demanding species more typical of earlier successional stages.
- Does the soil seed bank reflect species diversity in the extant vegetation and does this differ among the three forest types? We hypothesize that rainforest-associated species will be concentrated in extant vegetation and poorly represented in the soil seed bank, while the soil seed bank will support a greater abundance of early successional species.
2. Materials and Methods
2.1. Study Area and Site Selection
2.2. Vegetation Survey
2.3. Soil Seed Bank Treatment
2.4. Functional Types
2.5. Statistical Analyses
2.5.1. Fire-Related Germination Cues and Forest Type
2.5.2. Associations between Vegetation Pool and Forest Type
3. Results
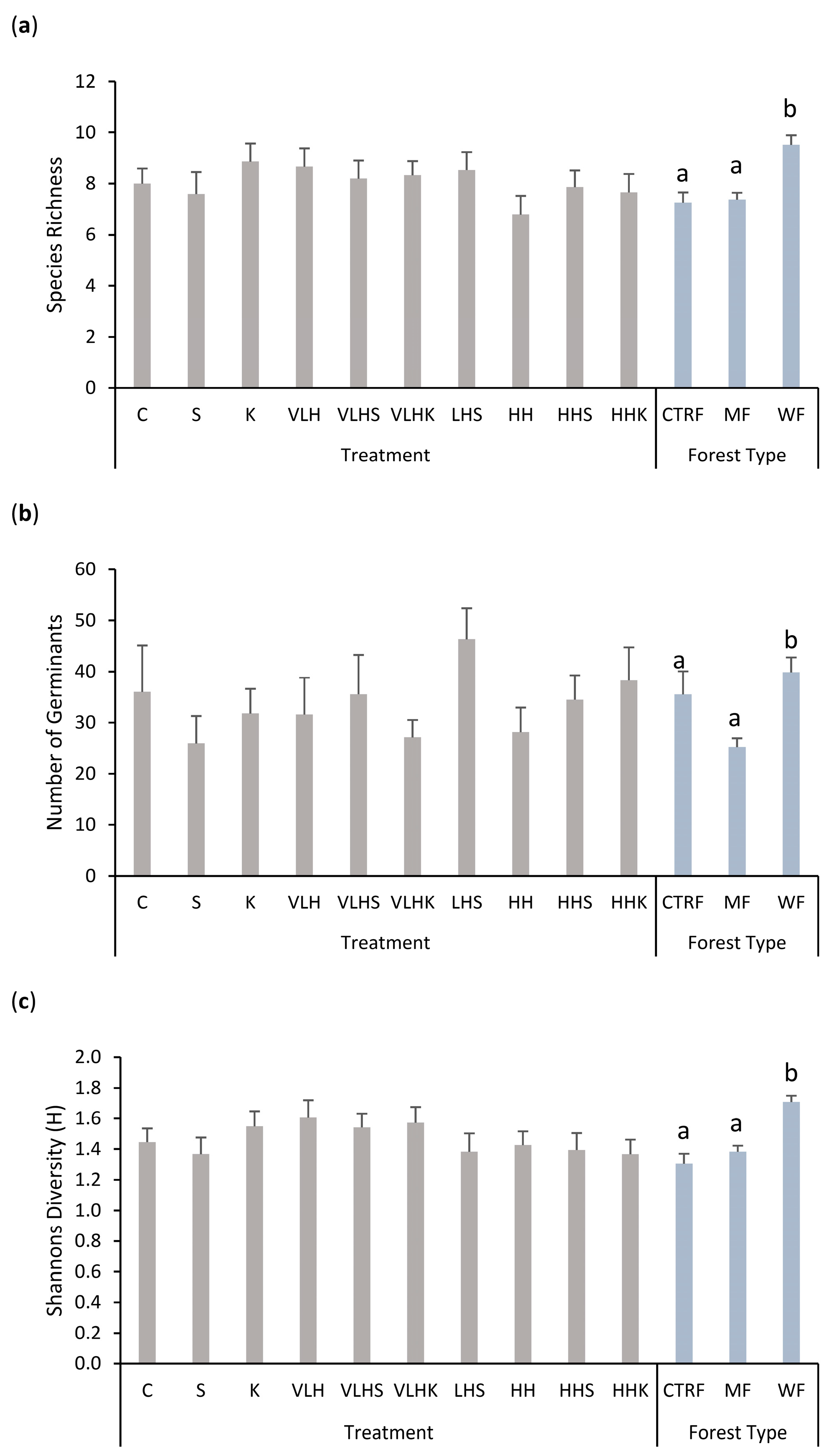
3.1. Fire-Related Germination Cues
3.2. Individual Species
3.3. All Species
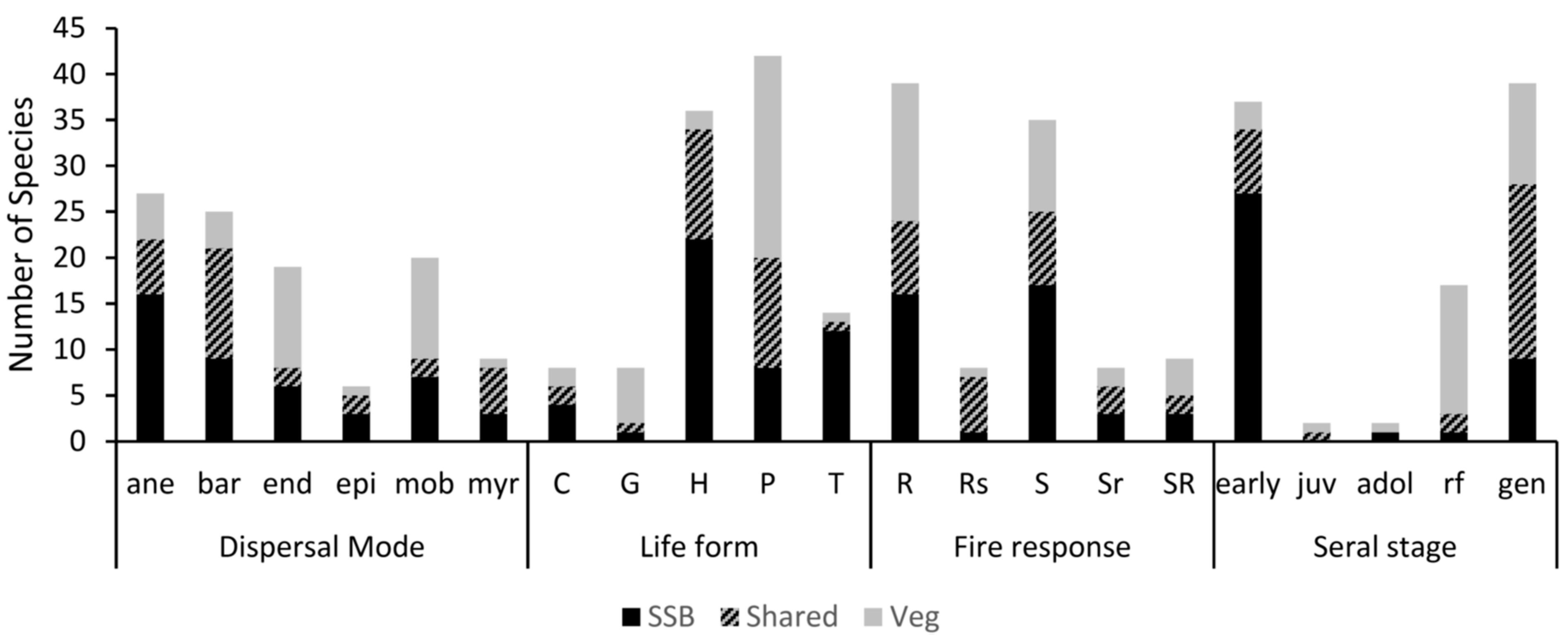
3.4. Functional Traits
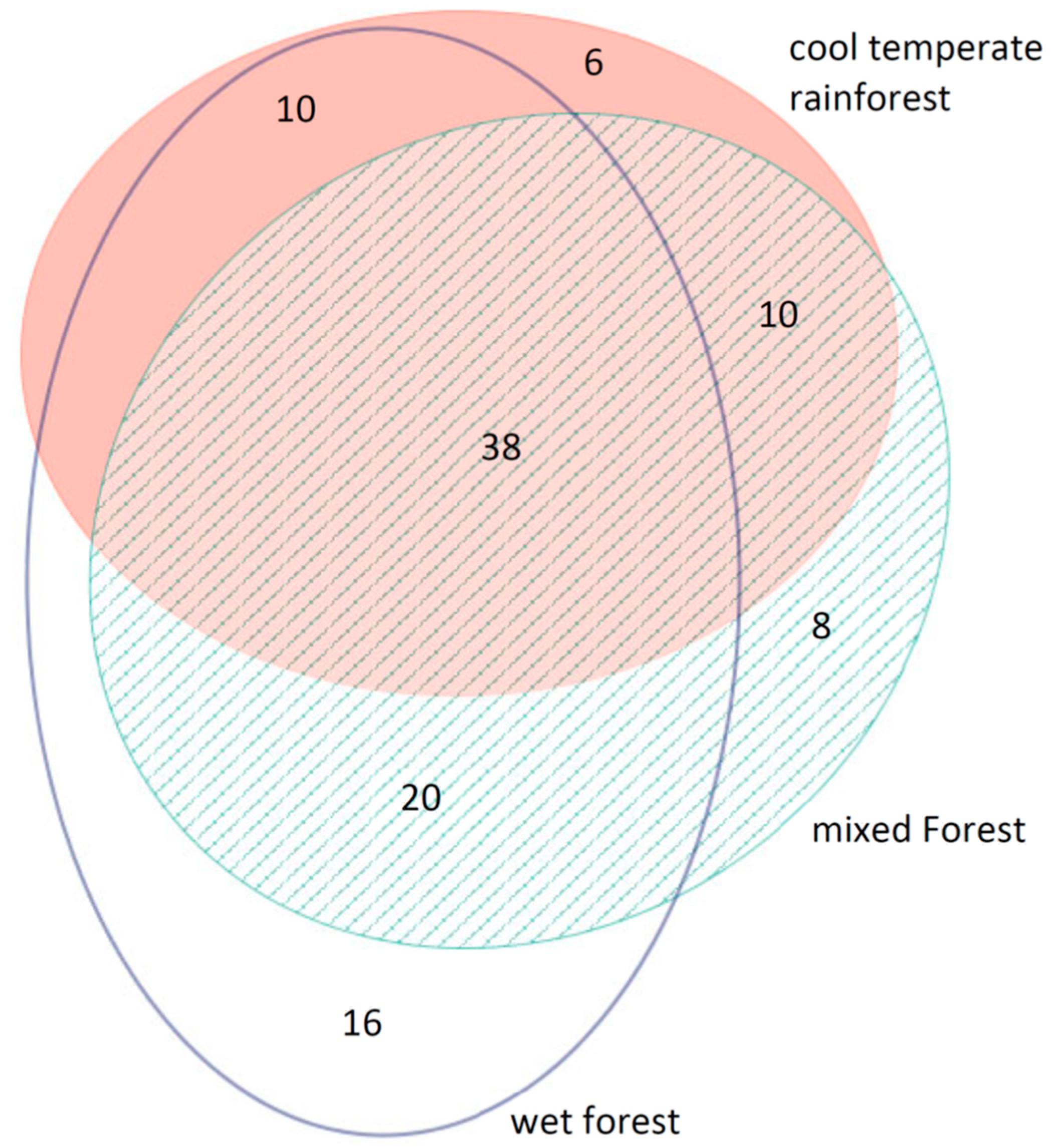
3.5. Vegetation Pool versus Forest Type
3.5.1. Soil Seed Bank Versus Extant Vegetation
3.5.2. Forest Type
4. Discussion
4.1. Does the Soil Seed Store Respond to Fire-Related Germination Cues and Do These Cues Differ among the Three Forest Types?
4.2. Does the Application of Karrikinolide Increase Germinant Diversity and Abundance and Is This Dependent on the Application of Heat?
4.3. Does the Abundance and Diversity of Soil Seed Banks Differ among Forest Types?
4.4. Does the Soil Seed Bank Reflect Species Diversity in the Extant Vegetation and Does This Differ among the Three Forest Types?
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Keeley, J.E.; Pausas, J.G. Evolutionary ecology of fire. Annu. Rev. Ecol. Evol. Syst. 2022, 53, 203–225. [Google Scholar] [CrossRef]
- Mariani, M.; Connor, S.E.; Theuerkauf, M.; Herbert, A.; Kunes, P.; Bowman, D.; Fletcher, M.S.; Head, L.; Kershaw, A.P.; Haberle, S.G.; et al. Disruption of cultural burning promotes shrub encroachment and unprecedented wildfires. Front. Ecol. Environ. 2022, 20, 292–300. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Williams, A.P. Impact of anthropogenic climate change on wildfire across western US forests. Proc. Natl. Acad. Sci. USA 2016, 113, 11770–11775. [Google Scholar] [CrossRef]
- Canadell, J.G.; Meyer, C.P.; Cook, G.D.; Dowdy, A.; Briggs, P.R.; Knauer, J.; Pepler, A.; Haverd, V. Multi-decadal increase of forest burned area in Australia is linked to climate change. Nat. Commun. 2021, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Clarke, H.; Evans, J.P. Exploring the future change space for fire weather in southeast Australia. Theor. Appl. Climatol. 2019, 136, 513–527. [Google Scholar] [CrossRef]
- Nolan, R.H.; Anderson, L.O.; Poulter, B.; Varner, J.M. Increasing threat of wildfires: The year 2020 in perspective: A Global Ecology and Biogeography special issue. Glob. Ecol. Biogeogr. 2022, 31, 1898–1905. [Google Scholar] [CrossRef]
- Stephens, S.L.; Burrows, N.; Buyantuyev, A.; Gray, R.W.; Keane, R.E.; Kubian, R.; Liu, S.R.; Seijo, F.; Shu, L.F.; Tolhurst, K.G.; et al. Temperate and boreal forest mega-fires: Characteristics and challenges. Front. Ecol. Environ. 2014, 12, 115–122. [Google Scholar] [CrossRef]
- Enright, N.J.; Fontaine, J.B.; Bowman, D.; Bradstock, R.A.; Williams, R.J. Interval squeeze: Altered fire regimes and demographic responses interact to threaten woody species persistence as climate changes. Front. Ecol. Environ. 2015, 13, 265–272. [Google Scholar] [CrossRef]
- Miller, A.D.; Thompson, J.R.; Tepley, A.J.; Anderson-Teixeira, K.J. Alternative stable equilibria and critical thresholds created by fire regimes and plant responses in a fire-prone community. Ecography 2019, 42, 55–66. [Google Scholar] [CrossRef]
- Fairman, T.A.; Nitschke, C.R.; Bennett, L.T. Too much, too soon? A review of the effects of increasing wildfire frequency on tree mortality and regeneration in temperate eucalypt forests. Int. J. Wildland Fire 2016, 25, 831–848. [Google Scholar] [CrossRef]
- McColl-Gausden, S.C.; Bennett, L.T.; Ababei, D.A.; Clarke, H.G.; Penman, T.D. Future fire regimes increase risks to obligate-seeder forests. Divers. Distrib. 2020, 28, 542–558. [Google Scholar] [CrossRef]
- Bowman, D.M.J.S.; Murphy, B.P.; Neyland, D.L.J.; Williamson, G.J.; Prior, L.D. Abrupt fire regime change may cause landscape-wide loss of mature obligate seeder forests. Glob. Chang. Biol. 2014, 20, 1008–1015. [Google Scholar] [CrossRef]
- Ashton, D.H.; Attiwill, P.M. Tall open forests. In Australian Vegetation; Groves, R.H., Ed.; Cambridge University Press: Cambridge, UK, 1994; pp. 157–196. [Google Scholar]
- Egler, F.E. Vegetation science concepts I. Initial floristic composition, a factor in old-field vegetation development with 2 figs. Vegetatio 1954, 4, 412–417. [Google Scholar] [CrossRef]
- Kasel, S.; Nitschke, C.R.; Baker, S.C.; Pryde, E.C. Concurrent assessment of functional types in extant vegetation and soil seed banks informs environmental constraints and mechanisms of plant community turnover in temperate forests of south-eastern Australia. For. Ecol. Manag. 2022, 519, 14. [Google Scholar] [CrossRef]
- Ashton, D.H. Fire in tall open-forests (wet sclerophyll). In Fire and the Australian biota; Gill, A.M., Groves, R.H., Noble, I.R., Eds.; Australian Academy of Science: Canberra, ACT, Australia, 1981; pp. 339–366. [Google Scholar]
- Pulsford, S.A.; Lindenmayer, D.B.; Driscoll, D.A. A succession of theories: Purging redundancy from disturbance theory. Biol. Rev. 2016, 91, 148–167. [Google Scholar] [CrossRef] [PubMed]
- Cameron, D. A Field Guide to Rainforest Identification in Victoria; Department of Sustainability and Environment: East Melbourne, VIC, Australia, 2011. [Google Scholar]
- Florence, R.G. Ecology and Silviculture of Eucalypt Forests; CSIRO Publishing: Clayton, VIC, Australia, 2004. [Google Scholar]
- Gilbert, J. Forest succession in the Florentine Valley, Tasmania. Pap. Proc. R. Soc. Tasman. 1959, 93, 129–152. [Google Scholar]
- Simkin, R.; Baker, P.J. Disturbance history and stand dynamics in tall open forest and riparian rainforest in the Central Highlands of Victoria. Austral Ecol. 2008, 33, 747–760. [Google Scholar] [CrossRef]
- Hickey, J.E. A floristic comparison of vascular species in Tasmanian oldgrowth mixed forest regeneration resulting from logging and wildfire. Aust. J. Bot. 1994, 42, 383–404. [Google Scholar] [CrossRef]
- Moroni, M.T.; Musk, R.; Wardlaw, T.J. Forest succession where trees become smaller and wood carbon stocks reduce. For. Ecol. Manag. 2017, 393, 74–80. [Google Scholar] [CrossRef]
- Riddington, M. Victoria’s rainforest and the potential impacts of a changing climate. Vic. Nat. 2014, 131, 209–218. [Google Scholar]
- Cunningham, T.M.; Cremer, K.W. Control of the understorey in wet eucalypt forests. Aust. For. 1965, 29, 4–14. [Google Scholar] [CrossRef]
- Fedrigo, M.; Kasel, S.; Bennett, L.T.; Roxburgh, S.H.; Nitschke, C.R. Carbon stocks in temperate forests of south-eastern Australia reflect large tree distribution and edaphic conditions. For. Ecol. Manag. 2014, 334, 129–143. [Google Scholar] [CrossRef]
- Peel, B. Rainforest Restoration Manual for South-Eastern Australia; CSIRO Publishing: Collingwood, VIC, Australia, 2010. [Google Scholar]
- Baker, P.J.; Simkin, R.; Pappas, N.; McLeod, A.; McKenzie, M. Fire on the mountain: A multi-scale, multi-proxy assessment of the resilience of cool temperate rainforest to fire in Victoria’s Central Highlands. In Peopled Landscapes (Terra Australis 34). Archaeological and Biogeographic Approaches to Landscapes; Haberle, S.G., David, B., Eds.; ANU Press: Canberra, ACT, Australia, 2012; pp. 375–391. [Google Scholar]
- Trouvé, R.; Oborne, L.; Baker, P.J. The effect of species, size, and fire intensity on tree mortality within a catastrophic bushfire complex. Ecol. Appl. 2021, 31, 14. [Google Scholar] [CrossRef]
- Stanton, P.; Parsons, M.; Stanton, D.; Stott, M. Fire exclusion and the changing landscape of Queensland’s Wet Tropics Bioregion 2. The dynamics of transition forests and implications for management. Aust. For. 2014, 77, 58–68. [Google Scholar] [CrossRef]
- Thomas, P.B.; Watson, P.J.; Bradstock, R.A.; Penman, T.D.; Price, O.F. Modelling surface fine fuel dynamics across climate gradients in eucalypt forests of south-eastern Australia. Ecography 2014, 37, 827–837. [Google Scholar] [CrossRef]
- Turton, S.M.; Sexton, G.J. Environmental gradients across four rainforest-open forest boundaries in northeastern Queensland. Aust. J. Ecol. 1996, 21, 245–254. [Google Scholar] [CrossRef]
- Crockett, J.; Mackey, B.; Ash, J. The Role of Fire in Governing the Relative Distribution of Rainforest and Sclerophyll Forest: The Effects Ofrainforest Vegetation on Fire Spread; School of Botany and Zoology, Australian National University: Canberra, ACT, Australia, 2006. [Google Scholar]
- Chesterfield, E.A.; Taylor, S.J.; Molnar, C.D. Recovery after Wildfire: Warm Temperate Rainforest at Jones Creek, East Gippsland, Victoria. Aust. For. 1991, 54, 157–173. [Google Scholar] [CrossRef]
- Hill, R.S.; Read, J. Post-fire regeneration of rainforest and mixed forest in Western Tasmania. Aust. J. Bot. 1984, 32, 481–493. [Google Scholar] [CrossRef]
- Tolsma, A.; Hale, R.; Sutter, G.; Kohout, M. Post-fire dynamics of Cool Temperate Rainforest in the O’Shannassy Catchment; DELWP: Eskdale, Australia, 2019. [Google Scholar]
- Campbell, M.L.; Keith, D.A.; Clarke, P.J. Regulation of seedling recruitment and survival in diverse ecotonal temperate forest understories. Plant Ecol. 2016, 217, 801–816. [Google Scholar] [CrossRef]
- Pausas, J.G.; Bradstock, R.A.; Keith, D.A.; Keeley, J.E. Plant functional traits in relation to fire in crown-fire ecosystems. Ecology 2004, 85, 1085–1100. [Google Scholar] [CrossRef]
- Maikano, G.N.; Cohn, J.; Di Stefano, J. Are germination cues for soil-stored seed banks different in structurally different fire-prone communities? Austral Ecol. 2018, 43, 89–101. [Google Scholar] [CrossRef]
- Pausas, J.G.; Lamont, B.B. Fire-released seed dormancy—A global synthesis. Biol. Rev. 2022, 97, 1612–1639. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.; Boulter, S.L.; Kitching, R.L. Heat and smoke effects on the germination of seeds from soil seed banks across forest edges between subtropical rainforest and eucalypt forest at Lamington National Park, south-eastern Queensland, Australia. Aust. J. Bot. 2003, 51, 227–237. [Google Scholar] [CrossRef]
- Fletcher, M.S.; Cadd, H.R.; Mariani, M.; Hall, T.L.; Wood, S.W. The role of species composition in the emergence of alternate vegetation states in a temperate rainforest system. Landsc. Ecol. 2020, 35, 2275–2285. [Google Scholar] [CrossRef]
- Auld, T.D.; O’Connell, M.A. Predicting patterns of post-fire germination in 35 eastern Australian Fabaceae. Aust. J. Ecol. 1991, 16, 53–70. [Google Scholar] [CrossRef]
- Enright, N.J.; Kintrup, A. Effects of smoke, heat and charred wood on the germination of dormant soil-stored seeds from a Eucalyptus baxteri heathy-woodland in Victoria, SE Australia. Austral Ecol. 2001, 26, 132–141. [Google Scholar] [CrossRef]
- Tierney, D.A. The effect of fire-related germination cues on the germination of a declining forest understorey species. Aust. J. Bot. 2006, 54, 297–303. [Google Scholar] [CrossRef]
- Carthey, A.J.R.; Tims, A.; Geedicke, I.; Leishman, M.R. Broad-scale patterns in smoke-responsive germination from the south-eastern Australian flora. J. Veg. Sci. 2018, 29, 737–745. [Google Scholar] [CrossRef]
- Shi, Y.F.; Shi, S.H.; Jiang, Y.S.; Liu, J. A global synthesis of fire effects on soil seed banks. Glob. Ecol. Conserv. 2022, 36, 9. [Google Scholar] [CrossRef]
- Bradshaw, S.D.; Dixon, K.W.; Hopper, S.D.; Lambers, H.; Turner, S.R. Little evidence for fire-adapted plant traits in Mediterranean climate regions. Trends Plant Sci. 2011, 16, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Williams, P.R. Fire-stimulated rainforest seedling recruitment and vegetative regeneration in a densely grassed wet sclerophyll forest of north-eastern Australia. Aust. J. Bot. 2000, 48, 651–658. [Google Scholar] [CrossRef]
- Williams, P.R.; Parsons, M.; Jensen, R.; Tran, C. Mechanisms of rainforest persistence and recruitment in frequently burnt wet tropical eucalypt forests. Austral Ecol. 2012, 37, 268–275. [Google Scholar] [CrossRef]
- Lloyd, M.V.; Dixon, K.W.; Sivasithamparam, K. Comparative effects of different smoke treatments on germination of Australian native plants. Austral Ecol. 2000, 25, 610–615. [Google Scholar] [CrossRef]
- Dixon, K.W.; Merritt, D.J.; Flematti, G.R.; Ghisalberti, E.L. Karrikinolide—A phytoreactive compound derived from smoke with applications in horticulture, ecological restoration and agriculture. Acta Hortic. 2009, 813, 155–170. [Google Scholar] [CrossRef]
- Flematti, G.R.; Ghisalberti, E.L.; Dixon, K.W.; Trengove, R.D. A compound from smoke that promotes seed germination. Science 2004, 305, 977. [Google Scholar] [CrossRef]
- Stevens, J.C.; Merritt, D.J.; Flematti, G.R.; Ghisalberti, E.L.; Dixon, K.W. Seed germination of agricultural weeds is promoted by the butenolide 3-methyl-2H-furo 2,3-c pyran-2-one under laboratory and field conditions. Plant Soil 2007, 298, 113–124. [Google Scholar] [CrossRef]
- Rokich, D.P.; Dixon, K.W.; Sivasithamparam, K.; Meney, K.A. Smoke, mulch, and seed broadcasting effects on woodland restoration in Western Australia. Restor. Ecol. 2002, 10, 185–194. [Google Scholar] [CrossRef]
- Nelson, D.C.; Riseborough, J.A.; Flematti, G.R.; Stevens, J.; Ghisalberti, E.L.; Dixon, K.W.; Smith, S.M. Karrikins discovered in smoke trigger Arabidopsis seed germination by a mechanism requiring gibberellic acid synthesis and light. Plant Physiol. 2009, 149, 863–873. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Pei, Y.H.; Chen, S.F.; Li, S.G.; Hu, X.L.; Chen, G.; Lin, B.; Wang, H.F. Three new butenolides from the fungus Aspergillus sp CBS-P-2. Molecules 2016, 21, 10. [Google Scholar] [CrossRef]
- Li, D.H.; Zhu, T.J.; Liu, H.B.; Fang, Y.C.; Gu, Q.Q.; Zhu, W.M. Four butenolides are novel cytotoxic compounds isolated from the marine-derived bacterium, Streptoverticillium luteoverticillatum 11014. Arch. Pharm. Res. 2006, 29, 624–626. [Google Scholar] [CrossRef]
- Wang, Y.M.; Peng, S.Q.; Zhou, Q.; Wang, M.W.; Yan, C.H.; Yang, H.Y.; Wang, G.Q. Depletion of intracellular glutathione mediates butenolide-induced cytotoxicity in HepG2 cells. Toxicol. Lett. 2006, 164, 231–238. [Google Scholar] [CrossRef]
- Ghebrehiwot, H.M.; Kulkarni, M.G.; Szalai, G.; Soos, V.; Balazs, E.; Van Staden, J. Karrikinolide residues in grassland soils following fire: Implications on germination activity. S. Afr. J. Bot. 2013, 88, 419–424. [Google Scholar] [CrossRef]
- Tng, D.Y.P.; Jordan, G.J.; Bowman, D.M.J.S. Plant traits demonstrate that temperate and tropical giant eucalypt forests are ecologically convergent with rainforest not savanna. PLoS ONE 2013, 8, 13. [Google Scholar] [CrossRef] [PubMed]
- Jackson, W.D. Fire, air, water and earth--an elemental ecology of Tasmania. Proc. Ecol. Soc. Aust. 1968, 3, 9–16. [Google Scholar]
- Wood, S.W.; Bowman, D.M.J.S. Alternative stable states and the role of fire-vegetation-soil feedbacks in the temperate wilderness of southwest Tasmania. Landsc. Ecol. 2012, 27, 13–28. [Google Scholar] [CrossRef]
- Campbell, M.L.; Clarke, P.J.; Keith, D.A. Seed traits and seed bank longevity of wet sclerophyll forest shrubs. Aust. J. Bot. 2012, 60, 96–103. [Google Scholar] [CrossRef]
- Plue, J.; Colas, F.; Auffret, A.G.; Cousins, S.A.O. Methodological bias in the seed bank flora holds significant implications for understanding seed bank community functions. Plant Biol. 2017, 19, 201–210. [Google Scholar] [CrossRef]
- Leckie, S.; Vellend, M.; Bell, G.; Waterway, M.J.; Lechowicz, M.J. The seed bank in an old-growth, temperate deciduous forest. Can. J. Bot.-Rev. Can. Bot. 2000, 78, 181–192. [Google Scholar] [CrossRef]
- Bossuyt, B.; Heyn, M.; Hermy, M. Concentrating samples estimates a larger seed bank density of a forest soil. Funct. Ecol. 2000, 14, 766–767. [Google Scholar] [CrossRef]
- Cornelissen, J.H.C.; Lavorel, S.; Garnier, E.; Diaz, S.; Buchmann, N.; Gurvich, D.E.; Reich, P.B.; ter Steege, H.; Morgan, H.D.; van der Heijden, M.G.A.; et al. A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust. J. Bot. 2003, 51, 335–380. [Google Scholar] [CrossRef]
- Raunkiaer, C. The Life Forms of Plants and Statistical Plant Geography; Oxford University Press: Oxford, UK, 1934. [Google Scholar]
- Kattge, J.; Diaz, S.; Lavorel, S.; Prentice, C.; Leadley, P.; Bonisch, G.; Garnier, E.; Westoby, M.; Reich, P.B.; Wright, I.J.; et al. TRY—A global database of plant traits. Glob. Chang. Biol. 2011, 17, 2905–2935. [Google Scholar] [CrossRef]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Anderson, M.J.; Gorley, R.N.; Clarke, K.R. PERMANOVA + for PRIMER: Guide to Software and Statistical Methods; PRIMER-E: Plymouth, England, 2008. [Google Scholar]
- Anderson, M.J.; Connell, S.D.; Gillanders, B.M.; Diebel, C.E.; Blom, W.M.; Saunders, J.E.; Landers, T.J. Relationships between taxonomic resolution and spatial scales of multivariate variation. J. Anim. Ecol. 2005, 74, 636–646. [Google Scholar] [CrossRef]
- Burrows, N.; Ward, B.; Wills, A.; Williams, M.; Cranfield, R. Fine-scale temporal turnover of jarrah forest understory vegetation assemblages is independent of fire regime. Fire Ecol. 2019, 15, 18. [Google Scholar] [CrossRef]
- Penman, T.; Binns, D.; Allen, R.; Shiels, R.; Plummer, S. Germinations responses of a dry sclerophyll forest soil-stored seedbank to fire related cues. Cunninghamia 2008, 10, 547–555. [Google Scholar]
- IBM SPSS for Windows, Version 28.0. IBM Corporation: Armonk, NY, USA, 2016.
- Meers, T.L.; Enright, N.J.; Bell, T.L.; Kasel, S. Deforestation strongly affects soil seed banks in eucalypt forests: Generalisations in functional traits and implications for restoration. For. Ecol. Manag. 2012, 266, 94–107. [Google Scholar] [CrossRef]
- Meers, T.L.; Bell, T.L.; Enright, N.J.; Kasel, S. Role of plant functional traits in determining vegetation composition of abandoned grazing land in north-eastern Victoria, Australia. J. Veg. Sci. 2008, 19, 515–524. [Google Scholar] [CrossRef]
- Keeley, J.E.; Pausas, J.G.; Rundel, P.W.; Bond, W.J.; Bradstock, R.A. Fire as an evolutionary pressure shaping plant traits. Trends Plant Sci. 2011, 16, 406–411. [Google Scholar] [CrossRef]
- Lindenmayer, D.B.; Taylor, C. New spatial analyses of Australian wildfires highlight the need for new fire, resource, and conservation policies. Proc. Natl. Acad. Sci. USA 2020, 117, 12481–12485. [Google Scholar] [CrossRef]
- Maisey, A.C.; Haslem, A.; Leonard, S.W.J.; Bennett, A.F. Foraging by an avian ecosystem engineer extensively modifies the litter and soil layer in forest ecosystems. Ecol. Appl. 2021, 31, 13. [Google Scholar] [CrossRef]
- Ashton, D.H.; Chinner, J.H. Problems of regeneration of the mature Eucalyptus regnans F. Muell, (The Big Ash) forest, in the absence of fire at Wallaby Creek, Victoria, Australia. Aust. For. 1999, 62, 265–280. [Google Scholar] [CrossRef]
- Ashton, D.H.; Bassett, O.D. The effects of foraging by the superb lyrebird (Menura novae-hollandiae) in Eucalyptus regnans forests at Beenak, Victoria. Aust. J. Ecol. 1997, 22, 383–394. [Google Scholar] [CrossRef]
- Ashton, D.H. The Big Ash Forest, Wallaby Creek, Victoria—Changes during one lifetime. Aust. J. Bot. 2000, 48, 1–26. [Google Scholar] [CrossRef]
- Brock, J.M.R.; Burns, B.R. Patterns of woody plant epiphytism on tree ferns in New Zealand. N. Z. J. Ecol. 2021, 45, 10. [Google Scholar] [CrossRef]
- Howard, T.M. Studies in the Ecology of Nothofagus Cunninghamii Oerst. I Natural Regeneration on the Mt. Donna Buang Massif, Victoria. Aust. J. Bot. 1973, 21, 67–78. [Google Scholar] [CrossRef]
- Ough, K.; Murphy, A. The effect of clearfell logging on tree-ferns in Victorian Wet Forest. Aust. For. 1996, 59, 178–188. [Google Scholar] [CrossRef]
- Winoto-Lewin, Y.; Kirkpatrick, J.B. Species of accidental woody epiphytes vary between host trees in Tasmanian wet forests. Aust. J. Bot. 2020, 68, 532–541. [Google Scholar] [CrossRef]
- Palmer, H.D.; Denham, A.J.; Ooi, M.K.J. Fire severity drives variation in post-fire recruitment and residual seed bank size of Acacia species. Plant Ecol. 2018, 219, 527–537. [Google Scholar] [CrossRef]
- Ashton, D.H.; Turner, J.S. Studiens on the light compensation point of Eucalyptus regnans F. Muell. Aust. J. Bot. 1979, 27, 589–607. [Google Scholar] [CrossRef]
- Vickers, H.; Kasel, S.; Duff, T.; Nitschke, C. Recruitment and growth dynamics of a temperate forest understorey species following wildfire in southeast Australia. Dendrochronologia 2021, 67, 9. [Google Scholar] [CrossRef]
- Floyd, A.G. Effect of burning on regeneration from seeds in wet sclerophyll forest. Aust. For. 1976, 39, 210–220. [Google Scholar] [CrossRef]
- White, D.J.; Vesk, P.A. Fire and legacy effects of logging on understorey assemblages in wet-sclerophyll forests. Aust. J. Bot. 2019, 67, 341–357. [Google Scholar] [CrossRef]
- Baker, S.C.; Kasel, S.; van Galen, L.G.; Jordan, G.J.; Nitschke, C.R.; Pryde, E.C. Identifying regrowth forests with advanced mature forest values. For. Ecol. Manag. 2019, 433, 73–84. [Google Scholar] [CrossRef]
- Jennings, S.M.; Hickey, J.E. Regeneration after seed-tree retention in tall Nothofagus rainforest in Tasmania. Tasforests 2003, 14, 15–22. [Google Scholar]
- Pausas, J.G.; Keeley, J.E. Evolutionary ecology of resprouting and seeding in fire-prone ecosystems. New Phytol. 2014, 204, 55–65. [Google Scholar] [CrossRef]
- Understorey_Network. Database of Native Species and How to Grow Them. Available online: http://www.understorey-network.org.au/ (accessed on 8 February 2023).
- Carroll, E.; Ashton, D.H. Seed storage in soils of several Victorian plant communities. Vic. Nat. 1965, 82, 102–110. [Google Scholar]
- Neyland, M.G.; Brown, M.J. Disturbance of cool temperate rainforest patches in eastern Tasmania. Aust. For. 1994, 57, 1–10. [Google Scholar] [CrossRef]
- Tabor, J.; McElhinny, C.; Hickey, J.; Wood, J. Colonisation of clearfelled coupes by rainforest tree species from mature mixed forest edges, Tasmania, Australia. For. Ecol. Manag. 2007, 240, 13–23. [Google Scholar] [CrossRef]
- Hickey, J.E.; Blakesley, A.J.; Turner, B. Seedfall and germination of Nothofagus cunninghamii (Hook.) Oerst., Eucryphia lucida (Labill.) Baill and Atherosperma moschatum Labill. Implications for regeneration practice. Aust. For. Res. 1983, 13, 21–28. [Google Scholar]
- Hickey, J.E.; Wilkinson, G.R. Long-term regeneration trends from a silvicultural systems trial in lowland cool temperate rainforest in Tasmania. Tasforest 1999, 11, 1–22. [Google Scholar]
- Le Breton, T.D.; Lyons, M.B.; Nolan, R.H.; Penman, T.; Williamson, G.J.; Ooi, M.K.J. Megafire-induced interval squeeze threatens vegetation at landscape scales. Front. Ecol. Environ. 2022, 20, 327–334. [Google Scholar] [CrossRef]
- Mackey, B.; Lindenmayer, D.; Norman, P.; Taylor, C.; Gould, S. Are fire refugia less predictable due to climate change? Environ. Res. Lett. 2021, 16, 13. [Google Scholar] [CrossRef]
- Beers, T.W.; Dress, P.E.; Wensel, L.C. Aspect transformation in site productivity research. J. For. 1966, 64, 691. [Google Scholar]
- DSNR. Soil Survey Standard Test Method—Particle Size Analysis; Department of Sustainable Natural Resources: NSW, Australia, 2002. [Google Scholar]
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- McCune, B.; Mefford, M.J. HyperNiche. Nonparametric Multiplicative Habitat Modeling, Version 2; MjM Software: Gleneden Beach, OR, USA, 2009. [Google Scholar]
- Post, W.M.; Kwon, K.C. Soil carbon sequestration and land-use change: Processes and potential. Glob. Chang. Biol. 2000, 6, 317–327. [Google Scholar] [CrossRef]
- Rayment, G.E.; Higginson, F.R. Australian Laboratory Handbook of Soil and Water Chemical Methods; Inkata Press: Port Melbourne, VIC, Australia, 1992. [Google Scholar]
- Rayment, G.E.; Lyons, D.J. Soil Chemical Methods: Australasia; CSIRO Publishing: Collingwood, VIC, Australia, 2010. [Google Scholar]
- DELWP. Vital Attributes Database; Unpublished Database; Arthur Rylah Institute, Department of Environment, Land, Water and Planning: Heidelberg, VIC, Australia, 2017. [Google Scholar]
- McIntyre, S.; Lavorel, S.; Tremont, R.M. Plant life-history attributes: Their relationships to disturbance response in herbaceous vegetation. J. Ecol. 1995, 83, 31–44. [Google Scholar] [CrossRef]
- Meers, T.L.; Bell, T.L.; Enright, N.J.; Kasel, S. Do generalisations of global trade-offs in plant design apply to an Australian sclerophyllous flora? Aust. J. Bot. 2010, 58, 257–270. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | Description | Maximum Temperature (°C) |
|---|---|---|
| C | Control | |
| S | Smoke (vermiculite product) | |
| K | karrikinolide | |
| LHS | Low heat plus smoke | 60–65 |
| HH | High heat | 85–90 |
| HHS | High heat plus smoke | 85–90 |
| HHK | High heat plus karrikinolide | 85–90 |
| VLH | Very low heat | 40–45 |
| VLHS | Very low heat plus smoke | 40–45 |
| VLHK | Very low heat plus karrikinolide | 40–45 |
| Trait and Attributes | Abbreviation | Description |
|---|---|---|
| Life form A | ||
| Chamaephyte | C | Persistent buds ≥1 cm and <20–30 cm above ground surface |
| Geophyte | G | Persistent buds buried to a depth of 2–3 cm |
| Hemicryptophytes | H | Persistent buds are in the immediate vicinity of the soil surface only, maximum height 1 cm |
| Phanerophyte | P | Persistent buds >20–30 cm on stems above the ground, includes twiners, vines, and epiphytes |
| Therophyte | T | Annual (monocarpic) plants, includes some facultatively perennial plants (polycarpic) that were judged to be predominantly annual |
| Dispersal mode B | ||
| Anemochory | ane | Wind-dispersed; pappus, coma, samara, or similar attachment |
| Barochory | bar | Gravity-dispersed; no apparent seed dispersal mechanism |
| Endozoochory | end | Ingestion by vertebrates (mainly mammals and birds); fleshy dispersal units (berries, drupes, or aggregate fruits) |
| Epizoochory | epi | Dispersal by adhesion to the outside of animals—usually on the hair of mammals, via appendages including barbs, hooks, spines, burrs, or awns |
| Mobile | mob | Long-distance wind dispersal of small seed; includes barochores with seed dimensions <0.5 mm, and mass generally <0.1 mg |
| Myrmecochory | myr | Ant-dispersed, elaiosome attached to seed to attract ants |
| Fire Response C | ||
| Obligate resprouters | R | Plants that rely on resprouting to regenerate after fire |
| Obligate seeders | S | Plants that do not resprout and rely on seeding to regenerate after fire |
| Facultative seeders | SR | Plants that can resprout and germinate seeds after fire |
| Weak seeders | Sr | Seeders that also have some capacity to resprout |
| Weak resprouters | Rs | Resprouters that have some capacity to regenerate from seed |
| Seral Stage D | ||
| Renewal to founding (early) | early | <3 years post fire |
| Juvenile | juv | 3–9 years post fire |
| Adolescent | adol | 9–35 years post fire |
| Mature | mat | 35–250 years post fire |
| Waning to Senescence (rainforest) | rf | ≥250 years post fire (rainforest associated species) |
| Generalist | gen | Species occurring across all seral stages <250 years |
| Trait and Attribute | Soil Seed Bank | Extant Vegetation | Dissimilarity (%) | ||
|---|---|---|---|---|---|
| mean | SE | mean | SE | ||
| Life form | Pseudo-F = 35.3, p = 0.0001 | ||||
| Hemicryptophyte | 9.80 | (0.82) | 3.80 | (0.62) | 36.9 |
| Therophyte | 2.20 | (0.40) | 0.20 | (0.11) | 33.6 |
| Phanerophyte | 8.07 | (0.57) | 11.07 | (0.69) | 10.4 |
| Geophyte | 1.07 | (0.07) | 3.00 | (0.24) | 10.0 |
| Chamaephyte | 1.53 | (0.17) | 0.93 | (0.21) | 9.1 |
| Dispersal mode | Pseudo-F = 9.7, p = 0.0001 | ||||
| Myrmecochore | 4.67 | (0.35) | 2.07 | (0.34) | 24.9 |
| Epizoochore | 1.27 | (0.21) | 0.47 | (0.17) | 21.1 |
| Barochore | 6.53 | (0.47) | 4.47 | (0.72) | 19.4 |
| Endozoochore | 3.07 | (0.46) | 3.07 | (0.30) | 13.0 |
| Anemochore | 4.07 | (0.68) | 3.40 | (0.19) | 10.8 |
| Mobile | 3.60 | (0.29) | 5.87 | (0.35) | 10.7 |
| Fire response | Pseudo-F = 5.1, p = 0.0002 | ||||
| Facultative seeder (SR) | 1.27 | (0.27) | 1.47 | (0.13) | 38.4 |
| Obligate resprouter (R) | 6.40 | (0.70) | 8.00 | (0.65) | 29.4 |
| Obligate seeder (S) | 6.73 | (0.65) | 4.60 | (0.36) | 20.8 |
| Weak seeder (Sr) | 2.27 | (0.21) | 1.73 | (0.27) | 6.9 |
| Weak resprouter (Rs) | 4.40 | (0.24) | 3.07 | (0.25) | 4.4 |
| Seral stage | Pseudo-F = 42.1, p = 0.0001 | ||||
| Early | 7.53 | (1.00) | 1.80 | (0.58) | 53.5 |
| Rainforest | 1.00 | (0.14) | 6.00 | (0.61) | 33.3 |
| Generalist | 12.47 | (0.82) | 10.27 | (1.23) | 13.2 |
| Origin | |||||
| Pseudo-F = 1.3, p = 0.256 | |||||
| Native | 20.00 | (1.06) | 18.53 | (1.09) | |
| Pseudo-F = 25.1, p = 0.0002 | |||||
| Introduced | 1.80 | (0.30) | 0.47 | (0.17) | |
| Pseudo-F = 12.5, p = 0.0021 | |||||
| All species | 24.40 | (1.29) | 19.33 | (1.26) | |
| Trait and Attribute | CTRF | Mixed Forest (MF) | Wet Forest (WF) | Dissimilarity (%) | ||||
|---|---|---|---|---|---|---|---|---|
| mean | SE | mean | SE | mean | SE | |||
| Life form | Pseudo-F = 3.2, p = 0.0043 | |||||||
| a | b | b | CTRF v WF | CTRF v MF | ||||
| Hemicryptophyte | 5.9 | (1.5) | 7.1 | (1.2) | 7.4 | (1.3) | 33.6 | 38.8 |
| Therophyte | 0.7 | (0.3) | 1.1 | (0.4) | 1.8 | (0.7) | 31.1 | 23.4 |
| Chamaephyte | 0.7 | (0.2) | 1.5 | (0.2) | 1.5 | (0.2) | 15.4 | 17.6 |
| Phanerophyte | 8.4 | (0.9) | 9.2 | (0.9) | 11.1 | (0.7) | 11.7 | 12.9 |
| Geophyte | 1.6 | (0.3) | 2.1 | (0.4) | 2.4 | (0.5) | 8.1 | 7.3 |
| Dispersal mode | Pseudo-F = 2.4, p = 0.0132 | |||||||
| a | ab | b | CTRF v WF | |||||
| Myrmecochore | 3.2 | (0.7) | 3.2 | (0.6) | 3.7 | (0.5) | 22.5 | |
| Epizoochore | 0.4 | (0.2) | 0.9 | (0.2) | 1.3 | (0.3) | 20.5 | |
| Endozoochore | 2.9 | (0.5) | 2.9 | (0.5) | 3.4 | (0.5) | 20.0 | |
| Barochore | 3.9 | (0.7) | 6.1 | (0.9) | 6.5 | (0.6) | 15.2 | |
| Anemochore | 2.8 | (0.4) | 3.6 | (0.2) | 4.8 | (0.9) | 13.1 | |
| Mobile | 4.4 | (0.6) | 4.9 | (0.6) | 4.9 | (0.4) | 8.5 | |
| Fire response | Pseudo-F = 2.1, p = 0.0353 | |||||||
| a | ab | b | CTRF v WF | |||||
| Obligate seeder (S) | 4.3 | (0.4) | 5.8 | (0.5) | 6.9 | (0.9) | 26.8 | |
| Obligate resprouter (R) | 6.0 | (0.9) | 7.4 | (0.9) | 8.2 | (0.7) | 26.4 | |
| Facultative seeder (SR) | 1.4 | (0.2) | 1.2 | (0.2) | 1.5 | (0.3) | 22.2 | |
| Weak seeder (Sr) | 1.5 | (0.3) | 2.1 | (0.3) | 2.4 | (0.2) | 16.8 | |
| Weak resprouter (Rs) | 3.4 | (0.3) | 3.6 | (0.4) | 4.2 | (0.4) | 7.8 | |
| Seral stage | Pseudo-F = 7.2, p = 0.0003 | |||||||
| a | b | b | CTRF v WF | CTRF v MF | ||||
| Early | 3.1 | (1.3) | 4.2 | (1.0) | 6.7 | (1.5) | 53.1 | 48.0 |
| Rainforest | 4.9 | (1.2) | 3.3 | (0.9) | 2.3 | (0.6) | 24.3 | 29.8 |
| Generalist | 8.3 | (1.4) | 12.2 | (1.0) | 13.6 | (0.9) | 22.7 | 22.1 |
| Origin | Pseudo-F = 7.0, p = 0.0036 | |||||||
| Native | 16.3a | (1.5) | 19.5b | (0.7) | 22.0b | (1.1) | ||
| Pseudo-F = 1.9, p = 0.1649 | ||||||||
| Introduced | 0.8 | (0.3) | 1.1 | (0.3) | 1.5 | (0.5) | ||
| Pseudo-F = 8.7, p = 0.0015 | ||||||||
| All species | 18.2a | (1.9) | 22.1b | (0.9) | 25.3b | (1.6) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Younis, S.; Kasel, S. Do Fire Cues Enhance Germination of Soil Seed Stores across an Ecotone of Wet Eucalypt Forest to Cool Temperate Rainforest in the Central Highlands of South-Eastern Australia? Fire 2023, 6, 138. https://doi.org/10.3390/fire6040138
Younis S, Kasel S. Do Fire Cues Enhance Germination of Soil Seed Stores across an Ecotone of Wet Eucalypt Forest to Cool Temperate Rainforest in the Central Highlands of South-Eastern Australia? Fire. 2023; 6(4):138. https://doi.org/10.3390/fire6040138
Chicago/Turabian StyleYounis, Samuel, and Sabine Kasel. 2023. "Do Fire Cues Enhance Germination of Soil Seed Stores across an Ecotone of Wet Eucalypt Forest to Cool Temperate Rainforest in the Central Highlands of South-Eastern Australia?" Fire 6, no. 4: 138. https://doi.org/10.3390/fire6040138