
Quaternary Mammals from Central-Western Argentina in the Stratigraphic Context of Southern South America

Abstract
:1. Introduction
2. Material and Methods
3. Geological and Sedimentary Context
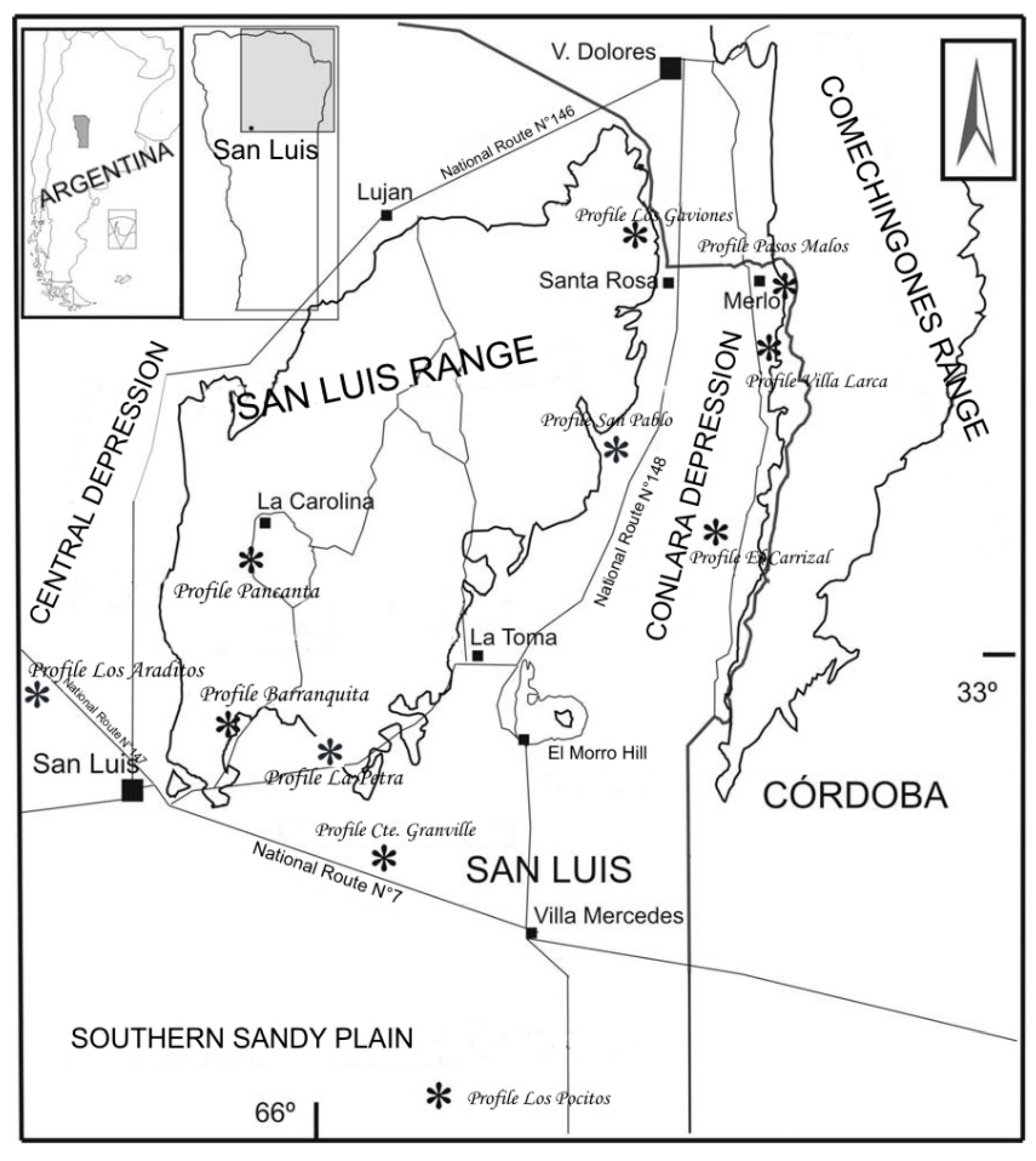
3.1. San Luis Range and Río Quinto Basin
3.2. Oriental or Conlara Depression
3.2.1. Oriental Sector (Pasos Malos Creek and Villa Larca Creek)
3.2.2. Western Sector (Conlara River Valley)
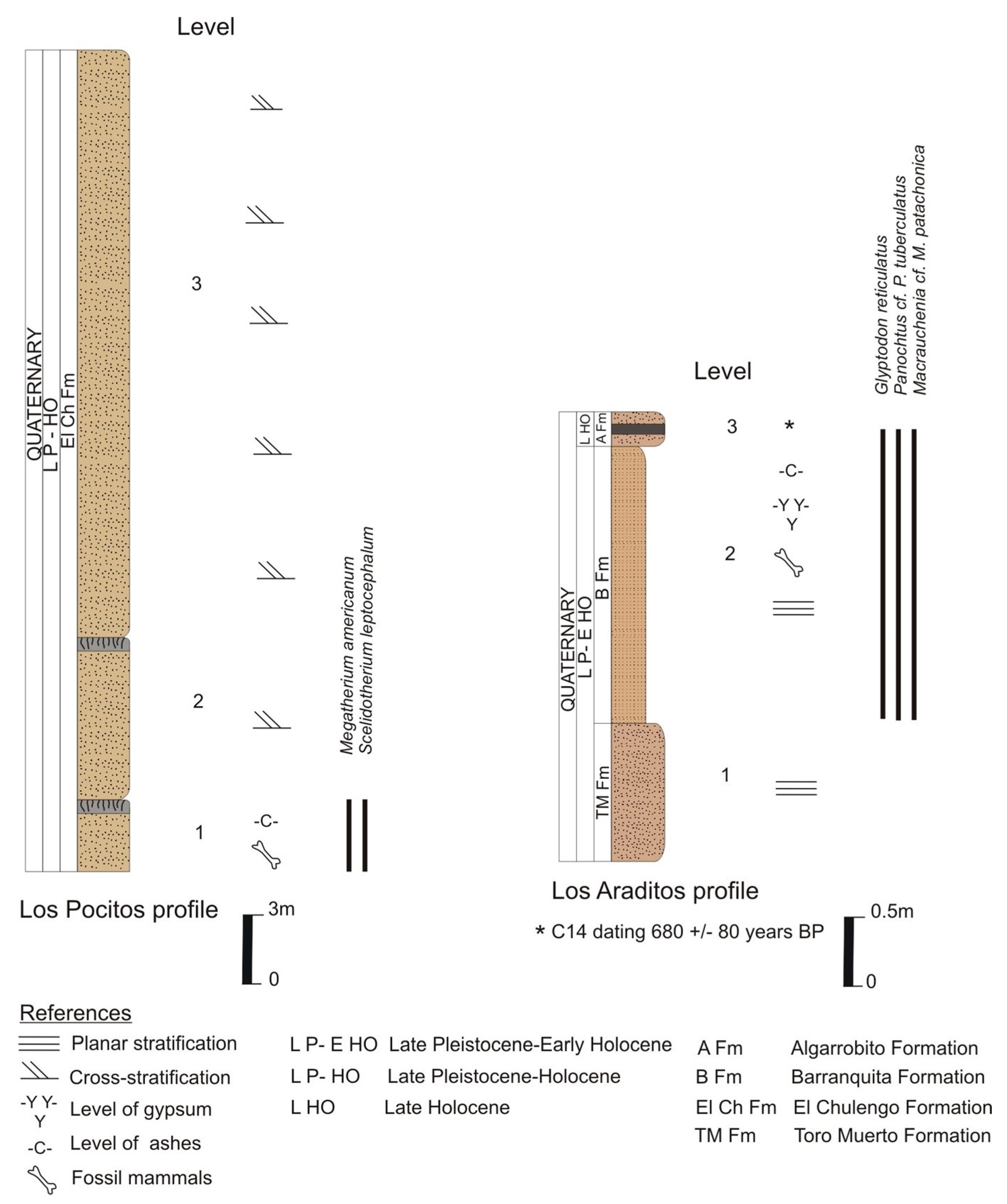
3.3. Southern Sandy Plain (Los Pocitos Lake)
3.4. Central Depression or Beazley Basin (Los Araditos Creek)
4. Systematic Paleontology
4.1. Xenarthra Cope, 1889
4.1.1. Cingulata Illiger, 1811
Glyptodontidae Gray, 1869
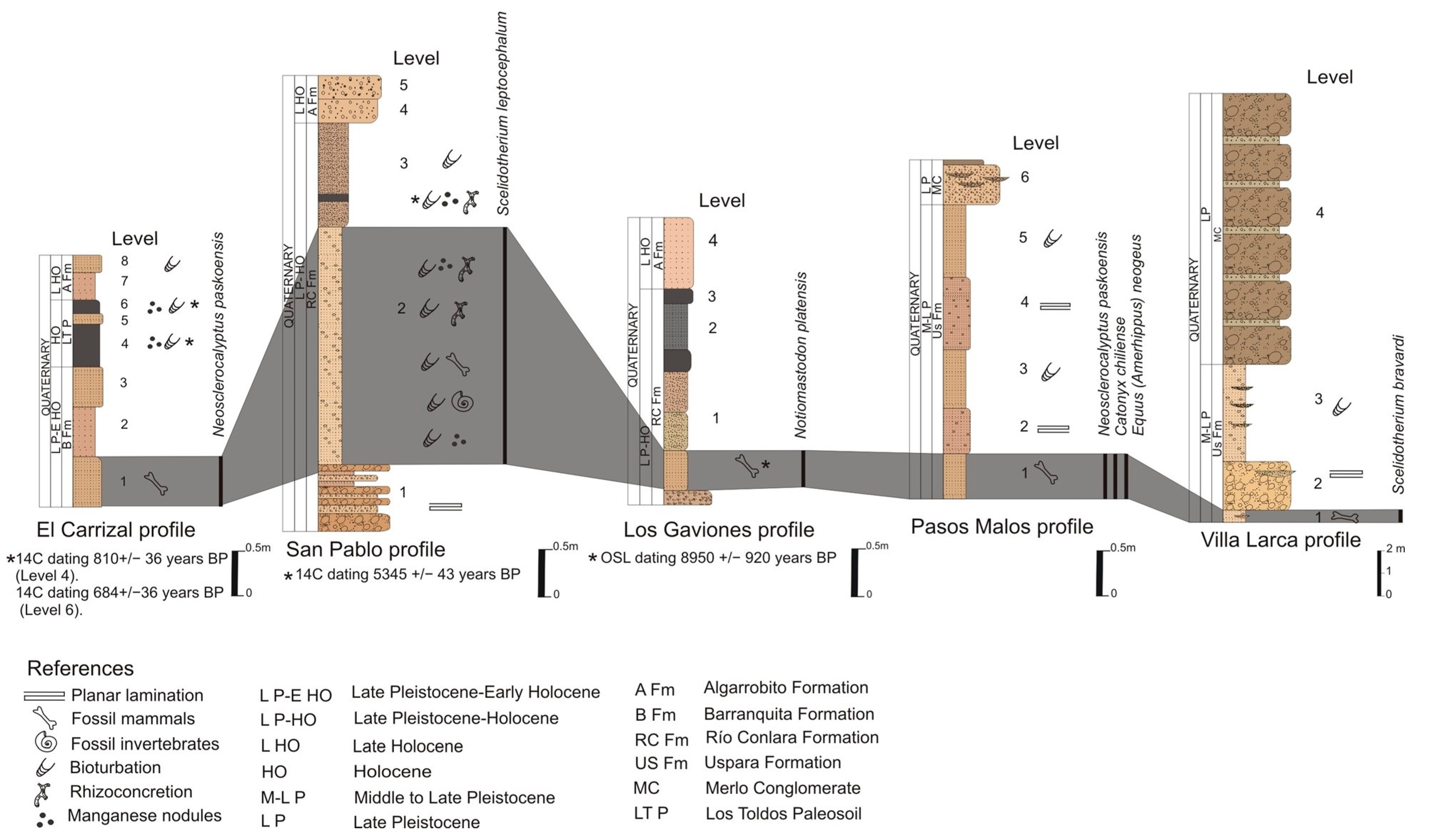
- Referred material. MHIN-UNSL-GEO V 198, complete dorsal carapace; Uspara Fm., Pasos Malos Creek, Merlo. MHIN-UNSL-GEO V 487–500, carapace, cephalic shield, caudal ring fragments, skull, mandible, vertebrae, ribs, scapula, pelvis, humeri, left ulna and radius, femora, tibio-fibulae, carpals, and tarsals of a single individual; Barranquita Fm., El Carrizal Creek.
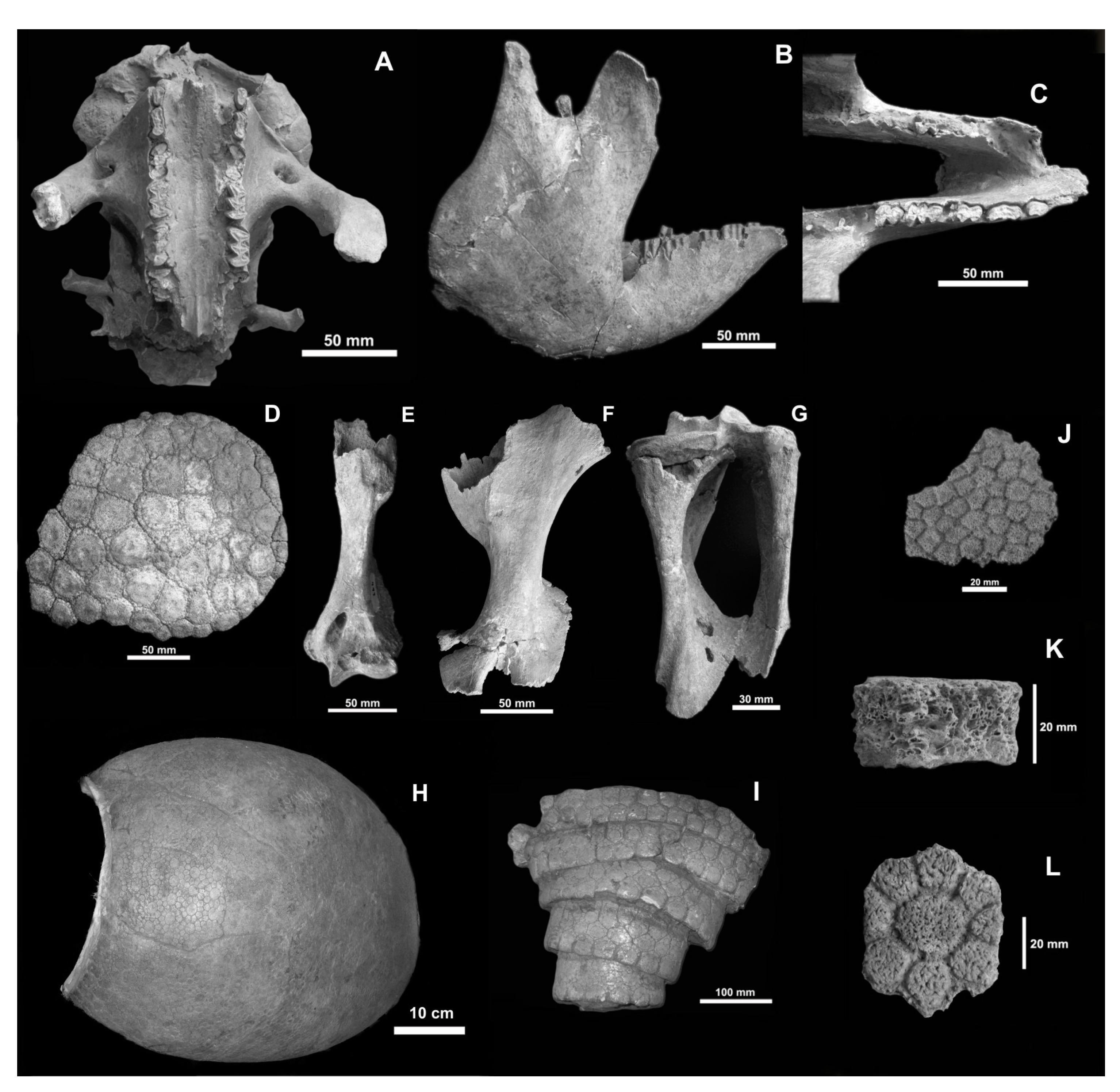
- Comments. The referred material had been mentioned by Bargo et al. [17], but the detailed data remained unpublished [18]. Morphological features correspond to Neosclerocalyptus paskoensis [22]. The dorsal carapace is oval and low, with a rather straight dorsal profile. The dorsal osteoderms are thin (3 mm thick), hexagonal to quadrangular, with well-defined sutures and piliferous foramina; the central figure is oval, somewhat depressed, and surrounded by small peripheral figures. Ventral osteoderms are smaller, rectangular, anteroposteriorly long, with a large central figure, and small or lacking peripheral figures. Caudal rings are formed by a double row of osteoderms, with strongly developed piliferous foramina. The skull has the parieto-temporal region directed ventrally; there is a concavity from the nuchal crest to the posterior third of frontals; the descending maxillary processes are not robust; frontonasal sinuses are strongly pneumatized and expanded laterally, with their borders separated from the rest of the skull; nasal cavities are separated from each other by a robust septum. The mandible is particularly slender in the ascending ramus, and the condyle and the coronoid process are approximately at the same level. Dimensions of different specimens are shown in the Supplementary Material, Tables S1 and S2. The femur MHIN-UNSL-GEO V 497 has a maximal length of 260 mm and a minimal width of the diaphysis of 54 mm. The length of the tibia MHIN-UNSL-GEO V 498 is 180 mm.
- Panochthini Castellanos, 1927
- Referred material. MHIN-UNSL-GEO V 638, 10 osteoderms of the dorsal carapace; Barranquita Fm., Los Araditos Creek.
- Comments. The osteoderms are very thick, with ornamentation composed of small polygonal, flattened figures, without differentiating a central figure or an ordering pattern. They resemble those of Panochthus tuberculatus, but the scarce material leads us to maintain a tentative identification.
- Referred material. MHIN-UNSL-GEO V 583, three osteoderms of the dorsal carapace; Uspara Fm., Papagayos Creek, Conlara Depression, Oriental sector. MHIN-UNSL-GEO V 466, 12 osteoderms of the dorsal carapace; Barranquita Fm., Los Araditos Creek.
- Comments. The osteoderms are very thick (20 mm) and show a central figure limited by a subcircular groove and radial grooves that delimit eight subcircular-polygonal peripheral figures. The surface of the osteoderms is rugose, with vascular and piliferous foramina. The fossil-bearing levels at Los Araditos Creek can be correlated with those at Chosmes Creek, in which an unpublished radiocarbon dating of a paleosol provides an age of around 7000 years (JOC personal data).
4.1.2. Pilosa Flower, 1883
Mylodontiidae Gill, 1872
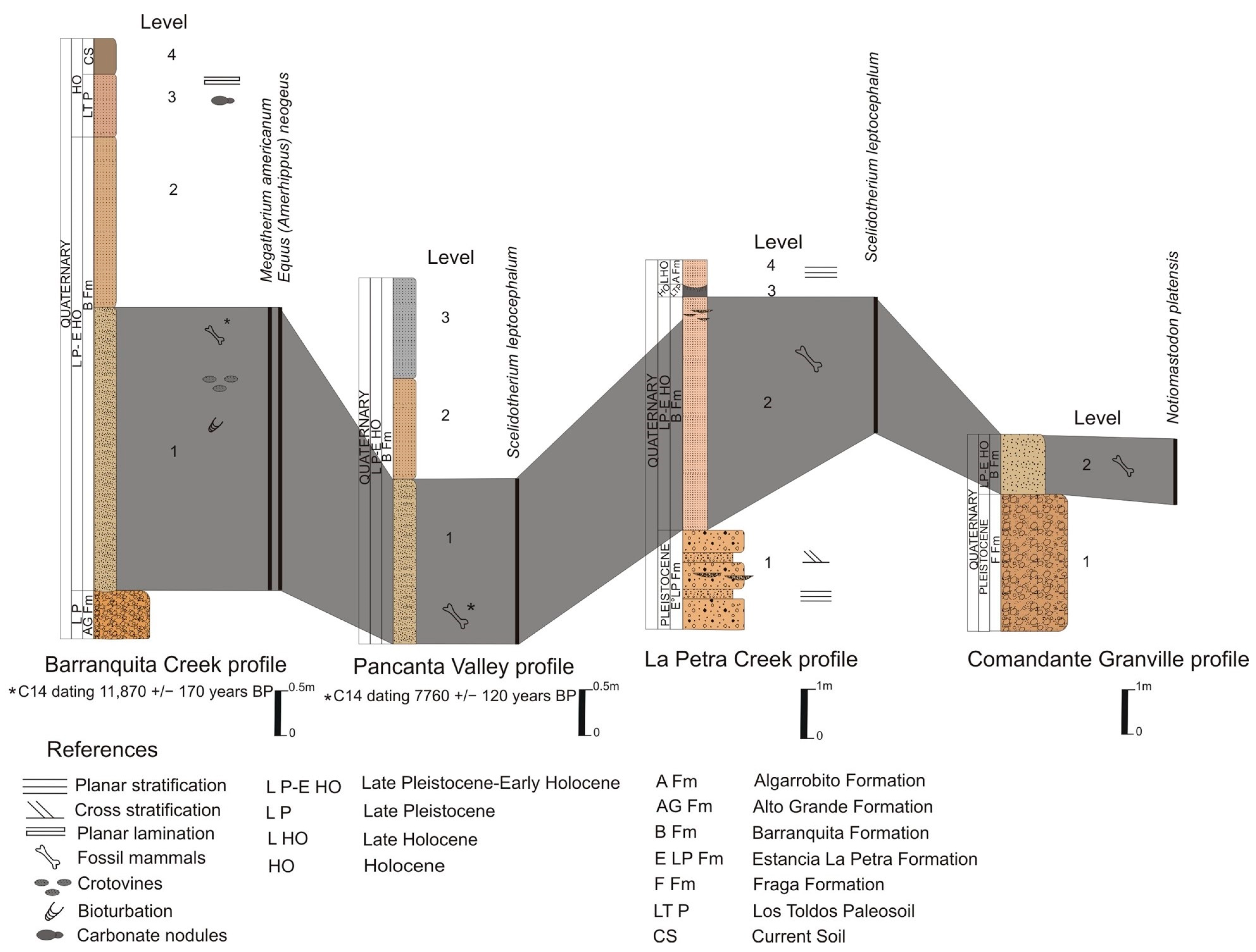
- Referred material. MHIN-UNSL-GEO V 200–211, tooth, caudal vertebrae, synsacrum, humeri, femora, tibiae, and left calcaneum; Barranquita Fm., Pancanta profile. MHIN-UNSL-GEO V 513, third phalanx; Barranquita Fm., La Petra Creek. MHIN-UNSL-GEO V 536, five lower and upper molariforms; Río Conlara Fm., Puente San Pablo, Conlara Valley. MHIN-UNSL-GEO V 690, left maxillary fragment with complete dentition, El Chulengo Fm., Los Pocitos Lake, Southern Sandy Plain.
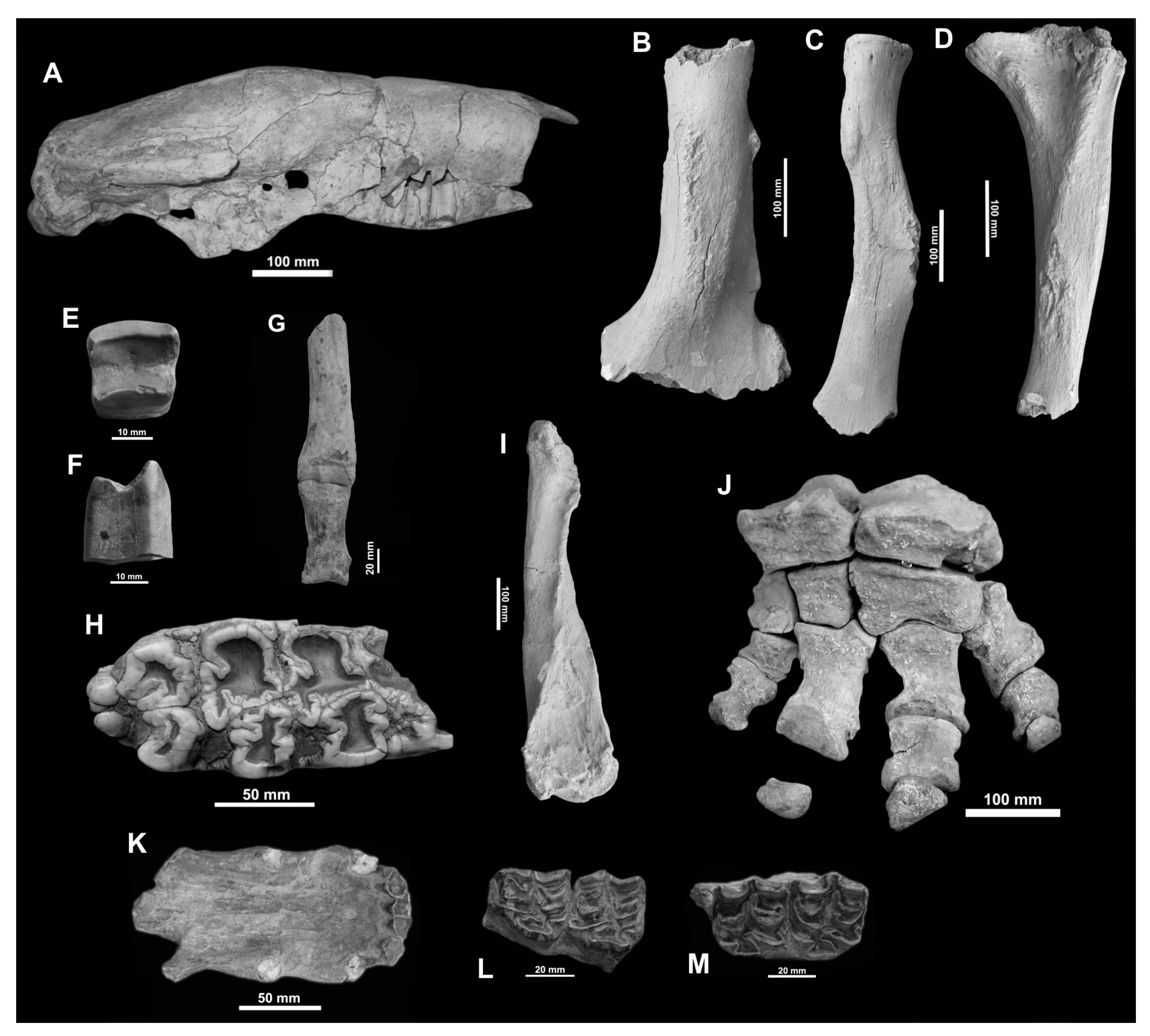
- Comments. Chiesa et al. [2] indicated an age of 7760 ± 120 years BP for levels in which MHIN-UNSL-GEO V 200–211 were found. Lucero [18], however, mentioned that MHIN-UNSL-GEO V 536 comes from a level placed below a paleosol dated to 5345 ± 43 years BP. Bargo et al. [17] mentioned the specimens MHIN-UNSL-GEO V 200-211, MHIN-UNSL-GEO V 372 (see below for this specimen) from Villa Larca Creek (Uspara Fm.), and MHIN-UNSL-GEO V 513 but without any description. Molars of MHIN-UNSL-GEO V 536 are identified as S. leptocephalum based on the following characters: caniniform cf1 subtriangular, with a small lingual lobe, and molariform mf3 slightly curved lingually in the posterior lobe. The complete series MHIN-UNSL-GEO V 690 occupies a length of 60 mm; Cf1 and Mf1–4 are subelliptic or subtriangular, obliquely implanted, and slightly anteroposteriorly elongated. The humerus is proximally massive, with a short diaphysis, and it enlarges distally; the entepicondylar foramen is preserved. The femur fragments lack the proximal epiphysis; the bone is anteroposteriorly flattened. The tibia is short and independent of the fibula. The calcaneus is large and extended posteriorly, with a typical distolateral diagonal crest. The astragalus has the typically concave cuboid facet of Scelidotheriinae and has a medial odontoid condyle. Dimensions of these bones can be found in the Supplementary Material, Tables S3 and S4. The ungual phalanx shows the proximal articulation extended posteriorly, with a crest between the two parallel facets; the palmar face has two large, deep foramina close to the articular border. The total length of the phalanx is 160 mm.
- (Figure 7J,K)
- Referred material. MHIN-UNSL-GEO V 372–374, cranial fragment with left jugal series and remains of right teeth, left mandibular fragment, and vertebral and rib remains; Uspara Fm., Villa Larca Creek.
- Comments. The horizontal ramus of the mandible shows a narrow, smoothly curved anterior area, twice as long as that occupied by jugal teeth. Upper teeth are prismatic, subtriangular-subelliptic in outline, anteroposteriorly elongated, and very oblique with respect to Cf1; Mf4 is smaller than the other molariforms. These remains were preliminarily identified as Scelidotherium cf. S. leptocephalum but later recognized as S. bravardi based on the mandibular morphology and a smaller size than S. leptocephalum [16,27]. The main dimensions of the mandible are as follows: total length = 380 mm; pre-dental length = 180 mm; height of the horizontal ramus at the level of mf1 = 85 mm.
- Referred material. MHIN-UNSL-GEO V 199, partial skull with only the right Cf1.
- Uspara Fm., Pasos Malos Creek (Merlo).
- Comments. This is one of the few Quaternary remains from San Luis that have been figured and described in detail [4]—as Scelidodon chiliense—and also included in a more recent paper by Miño-Boilini and Quiñones [28], in which materials from Tucumán Province and outside Argentina (Peru, Bolivia, Chile, and Ecuador) are also ascribed to Catonyx chiliensis. MHIN-UNSL-GEO V 199 was wrongly mentioned by Bargo et al. [17] as coming from Villa Larca Creek.
Megatheriidae Gray, 1821
- Referred material. MHIN-UNSL-GEO V 212–220, and V 511, right mf1 and mf4, and not associated postcranial remains. Barranquita Fm., Barranquita Creek. MHIN-UNSL-GEO V 512, one tooth and maxillary fragments, El Chulengo Fm., Los Pocitos Lake.
- Comments. Concerning the fossils from the Barranquita Profile, there is a radiocarbon date of 11,870 ± 170 years BP [2]. Besides the referred material, Bargo et al. [17] also mentioned a femur from the Fraga Fm. at Cantera Díaz Nogarol in the Río Quinto Basin, but this bone is not stored in an official institution.
4.2. Litopterna Ameghino, 1889
4.2.1. Macraucheniidae Gervais, 1855
Macraucheniinae Gervais, 1855
- Referred material. MHIN-UNSL-GEO V 467, lateral metacarpal (Mc II?) fragment and first phalanx in anatomical connection. Barranquita Fm., Los Araditos Creek.
- Comments. The distal curvature of the preserved metapodial allows its identification as a lateral digit (II or IV); it could correspond to a metacarpal rather than to a metatarsal, as the latter is more regularly concave [31,32] than in MHIN-UNSL-GEO V 467. The phalanx is high and relatively narrow, slightly asymmetrical, and its dimensions (Supplementary Material, Table S6) fall into the size range of M. patachonica, rather close to the mean values provided by Guérin and Faure [33]. This species is another common element of the Lujanian megafauna in Argentina and other South American areas.
4.3. Proboscidea Illiger, 1811
4.3.1. Gomphotheriidae Cabrera, 1929
Anancinae Hay, 1922
- Referred material. MHIN-UNSL-GEO V 376, molar; MHIN-UNSL-GEO V 510, 515, 221–224, associated postcranial elements (vertebrae, scapula, forelimb bones, and tibia), Río Conlara Fm., at Punta del Agua, Los Gaviones Profile (Conlara Depression, Western sector). MHIN-UNSL-GEO V 377–378, two worn molars, Fraga Fm., Comandante Granville Profile.
- Comments. The material from Punta del Agua comes from sediments dated by OSL to 8950 ± 920 years BP [2]. The recovered molars are incomplete and most are very worn; only MHIN-UNSL-GEO V 376 clearly shows the “clover” morphology, with few identified accessory conules; the enamel is typically rugose and striated along the crown base; transverse valleys are narrow; there is a uniform, thin cover of cement, except in the medial sulcus, where the main cusps of pretrite and postrite are in close contact. Postcranial bones are large and massive (Supplementary Material, Table S7). The most complete humerus shows a greatly developed epicondylar crest and a subcircular olecranian fossa; the radius has a subcircular, concave proximal facet. The preserved forefoot shows wide and short carpal bones, with flattened articular facets, and shortened metacarpals and phalanges. Both the dentition and bones can be clearly identified as the mastodon Notiomastodon platensis, a species broadly distributed in Argentina during the Upper Pleistocene.
4.4. Perissodactyla Owen, 1848
4.4.1. Equidae Gray, 1821
Equinae Steinman and Doderlein, 1890
- Referred material. MHIN-UNSL-GEO V 218a-b, two fragments of scapula, and V 468, a palatal fragment with incisors and canines, as well as maxillary fragments with molars, Barranquita Fm., Barranquita Creek. MHIN-UNSL-GEO V 227, isolated M1 and M2, Uspara Fm., Pasos Malos Creek (Merlo).
- Comments. The material from Barranquita Creek has a radiocarbon date of 11,870 ± 170 years BP [2]. The teeth show diagnostic characteristics of E. (A.) neogeus. They show projected parastyle and metastyle, limiting an angled concavity, a rounded metacone, a subtriangular protocone, a large hypocone, with a well-defined hypoconal sinus, wide fossettes, and a reduced pli caballin. The scapular fragments are not particularly diagnostic. Bargo et al. [17] mentioned both the scapula and the M1–2 as Equus (A.) cf. E. (A.) neogeus.
5. Final Remarks
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pascual, R.; Bondesio, P. Sedimentitas cenozoicas. Geología y Recursos Naturales de la Provincia de San Luis. In Proceedings of the Actas del VIII Congreso Geológico Argentino, San Luis, Argentina, 20–26 June 1981; pp. 117–154. [Google Scholar]
- Chiesa, J.O.; Lucero, N.P.; Basaez, A. Estratigrafía, paleontología y evidencias paleoambientales del Pleistoceno de San Luis. Op. Lilloana 2019, 52, 421–445. [Google Scholar]
- Chiesa, J.; Basaez, A.; Navío, J.; Lucero, N.; Ojeda, G.; Strasser, E. Neogene Stratigraphy of San Luis province, Argentina. In Cenozoic geology of the Central Andes of Argentina; Salfity, J.A., Marquillas, R.A., Eds.; SCS Publisher: Salta, Argentina, 2011; pp. 75–89. [Google Scholar]
- Miño-Boilini, A.R.; Carlini, A.A.; Chiesa, J.O.; Lucero, N.P.; Zurita, A.E. First record of Scelidodon chiliense (Lydekker) (Phyllophaga, Scelidotheriinae) from the Lujanian Stage (Late Pleistocene–Early Holocene) of Argentina. N. Jb. Geol. Paläont. Abh. 2009, 253, 373–381. [Google Scholar] [CrossRef]
- Basaez, A.; Chiesa, J.O.; Lucero, N.P. Estratigrafía y paleontología del sector medio de la cuenca Río Quinto (Mioceno superior-Pleistoceno tardío), San Luis, Argentina. Acta Geol. Lilloana 2013, 25, 37–48. [Google Scholar]
- Latrubesse, E.; Ramonell, C. Unidades litoestratigráficas del Cuaternario en la provincia de San Luis, Argentina. In Proceedings of the Actas del XI Congreso Geológico Argentino, San Juan, Argentina, 17–21 September 1990; pp. 109–112. [Google Scholar]
- De Moussy, J. Description Geographique et Statistique de la Confederacion Argentina; Firmin Didot Frères, Fils et Co.: Paris, France, 1860; Volume 1, pp. 1–582. [Google Scholar]
- Ave Lallemant, G. Apuntes sobre la geognosia de la sierra de San Luis. Acad. Nac. Ciencias 1875, 1, 106–108. [Google Scholar]
- Adaro, D. Fósiles y prehistoria; Conferencia en el Centro Ameghino. Escuela Normal de San Luis; Ministerio de Educación, Dirección de Cultura, Gobierno de San Luis: San Luis, Argentina, 1917; pp. 1–37. [Google Scholar]
- Frenguelli, J. Observaciones Geográficas y Geológicas en la Región de Sayapé (Pcia. de San Luis); Escuela Normal Superior ‘José María Torres’: Paraná, Argentina, 1931; pp. 1–68. [Google Scholar]
- Gez, J. Geografía de la Provincia de San Luis; Ediciones Peuser, Academia Nacional de Geografía: Buenos Aires, Argentina, 1938; Volume 1, pp. 1–68. [Google Scholar]
- Tapia, A.; Rigal, R. Geología del Paso de Las Carretas y sus alrededores. Bol. Direc. Nac. Minas y Geología 1933, 37, 1–56. [Google Scholar]
- Pastore, F.; Ruiz Huidobro, O. Descripción Geológica de la Hoja 24g. Saladillo (Provincia de San Luis). Bol. Direc. Nac. Minería 1952, 78, 1–63. [Google Scholar]
- Pastore, F.; González, R. Descripción geológica de la Hoja 23 g, San Francisco (San Luis). Bol. Direc. Nac. Minería 1954, 80, 1–61. [Google Scholar]
- Tognelli, G.; Chiesa, J.; Strasser, E.; Prado, J. Registro de mamíferos fósiles, cronología y ambiente sedimentario del Cuaternario de la sierra de San Luis y depresión de Conlara, San Luis, Argentina. In Proceedings of the Actas del IX Congreso Geológico Chileno, Puerto Varas, Chile, 31 July–4 August 2000; Volume 1, pp. 568–572. [Google Scholar]
- Miño-Boilini, A.R.; Carlini, A.A. The Scelidotheriinae Ameghino, 1904 (Phyllophaga, Xenarthra) from the Ensenadan–Lujanian Stage/Ages (Early Pleistocene to Early–Middle Pleistocene–Early Holocene) of Argentina. Quat. Intern. 2009, 210, 93–101. [Google Scholar] [CrossRef]
- Bargo, M.S.; Montalvo, C.I.; Chiesa, J.O.; Forasiepi, A.M.; Cerdeño, E.; Lucero, N.P.; Martinelli, A.G. El registro de mamíferos del Pleistoceno Tardío–Holoceno Temprano del centro oeste de Argentina. In Condiciones Paleoambientales y Ocupaciones Humanas del Centro-Oeste de Argentina durante el Pleistoceno Tardío-Holoceno y Holoceno; Zárate, M., Gil, A., Neme, G., Eds.; Publicaciones de la Sociedad Argentina de Antropología: Buenos Aires, Argentina, 2010; pp. 213–238. [Google Scholar]
- Lucero, N.P. Bioestratigrafía y Paleontología del Neógeno y Cuaternario de San Luis, Argentina. Ph.D. Thesis, Facultad de Ciencias Físico-Matemáticas y Naturales, Universidad Nacional de San Luis, San Luis, Argentina, 2016. [Google Scholar]
- Strasser, E.; Tognelli, G.; Chiesa, J.; Prado, J. Estratigrafía y Sedimentología de los depósitos eólicos del Pleistoceno tardío y Holoceno en el sector suroeste de la sierra de San Luis. In Proceedings of the Actas XIII Congreso Geológico Argentino and III Congreso de Exploración de Hidrocarburos, Buenos Aires, Argentina, 13–18 October 1996; Volume 4, pp. 73–83. [Google Scholar]
- Tripaldi, A.; Zárate, M. A review of late Quaternary inland dune systems of South America east of the Andes. Quat. Intern. 2016, 410, 96–110. [Google Scholar] [CrossRef]
- Iriondo, M. Climatic changes in the South American plains: Records of a continent-scale oscillation. Quat. Intern. 1999, 57–58, 93–112. [Google Scholar] [CrossRef]
- Zurita, A.E. Nuevo gliptodonte (Mammalia, Glyptodontoidea) del Cuaternario de la provincia de Chaco, Argentina. Ameghiniana 2002, 39, 175–182. [Google Scholar]
- Owen, R. Descriptive and Illustrated Catalogue of the Fossil Organic Remains of Mammalia and Aves Contained in the Museum of the Royal College of Surgeons of England; Legare Street Press: London, UK, 1845; pp. 1–391. [Google Scholar]
- Burmeister, G. Lista de los mamíferos fósiles del terreno diluviano. An. Mus. Públ. Buenos Aires 1866, 1, 121–232. [Google Scholar]
- Owen, R. Fossil Mammalia. In The Zoology of the Voyage of HSM Beagle; Darwin, C., Ed.; Smith Elder and Co.: London, UK, 1838; pp. 1–40. [Google Scholar]
- Lydekker, R. Description of three species of Scelidotherium. Proc. Zool. Soc. 1886, 32, 491–498. [Google Scholar]
- Miño-Boilini, A.R.; Carlini, A.A.; Scillato-Yané, G.J. Revisión sistemática y taxonómica del género Scelidotherium Owen, 1839 (Xenarthra, Phyllophaga, Mylodontidae). Rev. Brasil. Paleontol. 2014, 17, 43–58. [Google Scholar] [CrossRef]
- Miño-Boilini, A.R.; Quiñones, S.L. Los perezosos Scelidotheriinae (Xenarthra, Folivora): Taxonomía, biocronología y biogeografía. Rev. Mus. Arg. Cs. Nat. 2020, 22, 201–218. [Google Scholar] [CrossRef]
- Cuvier, G. Notice sur le squelette d’une très-grande espèce de quadrupède inconnue jusqu’à présent trouvé au Paraguay, et déposé au cabinet d’histoire naturelle de Madrid. Mag. Encycl. J. Sci. Lett. Arts. 1796, 1, 303–310. [Google Scholar]
- Forasiepi, A.M.; Martinelli, A.; Blanco, J. Bestiario Fósil. Mamíferos del Pleistoceno de la Argentina; Editorial Albatros: Buenos Aires, Argentina, 2007; pp. 1–190. [Google Scholar]
- Scherer, C.S.; Pitana, V.G.; Ribeiro, A.M. Proterotheriidae and Macraucheniidae (Litopterna, Mammalia) from the Pleistocene of Rio Grande do Sul State, Brazil. Rev. Brasil. Paleontol. 2009, 12, 231–246. [Google Scholar] [CrossRef]
- Gelfo, J.N.; Fernández-Monescillo, M.; Flores-Aqueveque, V.; Martínez Rivera, I. Macrauchenia. Un Enigmático Animal Extinto de la Cuenca de Calama; Martínez Rivera, I., Rojas Mondaca, O., Eds.; Publicación Especial del Museo de Historia Natural y Cul-tural del Desierto de Atacama: Atacama, Chile, 2020; pp. 1–90. [Google Scholar]
- Guérin, C.; Faure, M. Macrauchenia patachonica Owen (Mammalia, Litopterna) de la région de São Raimundo Nonato (Piauí, Nordeste brésilien) et la diversité des Macraucheniidae pléistocènes. Geobios 2004, 37, 513–535. [Google Scholar] [CrossRef]
- Ameghino, F. Rápidas Diagnosis de Algunos Mamíferos Fósiles Nuevos de la República Argentina; P.E. Coni: Buenos Aires, Argentina, 1888; pp. 1–17. [Google Scholar]
- Lund, P. Nouvelles recherches sur la faune fossile du Brésil. Ann. Sci. Nat. 1840, 13, 310–319. [Google Scholar]
- Alberdi, M.T.; Prado, J.L. Caballos Fósiles de América del Sur. Una Historia de Tres Millones de Años; INCUAPA, Facultad de Ciencias Sociales: Olavarría, Argentina, 2004; pp. 1–269. [Google Scholar]
- Prado, J.L.; Alberdi, M.T. Fossil Horses of South America. Phylogeny, Systematics and Ecology, 1st ed.; The Latin American Studies Book Series; Springer: Cham, Switzerland, 2017; pp. 1–150. [Google Scholar]
- Prado, J.L.; Alberdi, M.T.; Martínez, G.; Gutiérrez, M.A. Equus (Amerhippus) neogeus Lund, 1840 (Equidae, Perissodactyla) at Paso Otero 5 site, Argentina: Its implications for the extinction of the South American horse. N. Jb. Geol. Paläont. Mh. 2005, 8, 449–468. [Google Scholar] [CrossRef]
- Tonni, E.P. Mamíferos del Holoceno en la provincia de Buenos Aires. Paula-Coutiana 1990, 4, 3–21. [Google Scholar]
- Zurita, A.E. Sistemática y Evolución de los Hoplophorini (Xenarthra, Glyptodontidae, Hoplophorinae. Mioceno Tardío-Holoceno Temprano). Importancia Bioestratigráfica, Paleobiogeográfica y Paleoambiental. Ph.D. Thesis, Universidad Nacional de La Plata, La Plata, Argentina, 2007. [Google Scholar]
- Zamorano, M.; Mones, A.; Scillato-Yané, G. Redescripción y designación de un neotipo de Panochthus tuberculatus (Owen) (Mammalia, Cingulata, Glyptodontidae). Rev. Brasil. Paleontol. 2012, 15, 113–122. [Google Scholar] [CrossRef]
- Ferrero, B.S.; Noriega, J.I.; Brunetto, E.; Otaño, N.N. Vertebrate continental assemblage from the Last Interglacial in southern South America (Entre Ríos, Argentina). Biostratigraphy and paleoenvironment. Palaeogeogr. Palaeoclim. Palaeoecol. 2017, 466, 89–99. [Google Scholar] [CrossRef]
- Ferrero, B.S.; Patterer, N.I.; Brunetto, E.; Ramos, R.S.; Noriega, J.I.; Moya, E.; Peralta, M.J.; Zucol, A.F.; Franco, M.J.; Brea, M. El registro paleontológico del Pleistoceno de la provincia de Entre Ríos (Argentina). Op. Lilloana 2019, 52, 490–519. [Google Scholar]
- Forasiepi, A.M.; Martinelli, A.G.; Gil, A.; Neme, G.; Cerdeño, E. Fauna extinta y ocupaciones humanas en el Pleistoceno final–Holoceno temprano del centro occidental argentino. In Zooarqueología a Principios del siglo XXI. Aportes Teóricos, Metodológicos y Casos de Studio; Gutiérrez, M.A., De Negris, M., Fernández, P.M., Giardina, M., Gil, A., Izeta, A., Neme, G., Yacobaccio, H., Eds.; Ediciones del Espinillo: Buenos Aires, Argentina, 2010; pp. 219–229. [Google Scholar]
- Carlini, A.A.; Zurita, A.E.; Miño-Boilini, A.R. Reseña paleobiogeográfica de los Xenarthra (Mammalia) del Pleistoceno tardío de la región Mesopotámica (Argentina). INSUGEO Miscelánea 2008, 17, 259–270. [Google Scholar]
- Montalvo, C.I.; Zárate, M.A.; Bargo, M.S.; Mehl, A. Registro faunístico y paleoambientes del Cuaternario Tardío, provincia de La Pampa, Argentina. Ameghiniana 2013, 50, 554–570. [Google Scholar] [CrossRef]
- Chimento, N.R.; Agnolín, F.L. Mamíferos del Pleistoceno Superior de Santiago del Estero (Argentina) y sus afinidades paleobiogeográficas. Papéis Av. Zool. 2011, 51, 83–100. [Google Scholar] [CrossRef]
- Pascual, R. Paleontografía Bonaerense. IV. Vertebrata; Borrello, A.V., Ed.; Comisión de Investigación Científica: Buenos Aires, Argentina, 1966; pp. 1–202. [Google Scholar]
- Zetti, J. El hallazgo de un Megatheriidae en el “médano invasor” del SW de Toay, provincia de La Pampa. Ameghiniana 1964, 3, 257–265. [Google Scholar]
- Rusconi, C. Sobre un fémur de megaterio descubierto en El Borbollón, Mendoza. Rev. Mus. Hist. Nat. 1947, 1, 60–64. [Google Scholar]
- Bond, M. Quaternary native ungulates of Southern South America. A synthesis. In Quaternary of South America and Antarctic Peninsula; Tonni, E.P., Cione, A.L., Eds.; A.A. Balkema: Rotterdam, The Netherlands, 1999; Volume 12, pp. 177–205. [Google Scholar]
- Alberdi, M.T.; Prado, J.L.; Cartelle, C. El registro de Stegomastodon (Mammalia, Gomphotheriidae) en el Pleistoceno superior de Brasil. Rev. Esp. Paleontol. 2002, 17, 217–236. [Google Scholar] [CrossRef]
- Alberdi, M.T.; Cerdeño, E.; Prado, J.L. Stegomastodon platensis (Proboscidea, Gomphotheriidae) en el Pleistoceno de Santiago del Estero, Argentina. Ameghiniana 2008, 45, 257–271. [Google Scholar]
- Prado, J.L.; Alberdi, M.T. Global evolution of Equidae and Gomphotheriidae from South America. Integrative Zool. 2014, 9, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Mothé, D.; Avilla, L.A.; Cozzuol, M.; Winck, G.R. Taxonomic revision of the Quaternary gomphotheres (Mammalia: Proboscidea: Gomphotheriidae) from the South American lowlands. Quat. Intern. 2012, 276–277, 2–6. [Google Scholar] [CrossRef]
- Prado, J.L.; Alberdi, M.T.; Gómez, G. Late Pleistocene gomphotheres (Proboscidea) from the Arroyo Tapalqué locality (Buenos Aires, Argentina) and their taxonomic and biogeographic implication. N. Jb. Geol. Paläont. Abh. 2002, 225, 275–296. [Google Scholar] [CrossRef]
- Sánchez, B.; Prado, J.L.; Alberdi, M.T. Feeding ecology, dispersal, and extinction of South American Pleistocene gomphotheres (Gomphotheriidae, Proboscidea). Paleobiology 2004, 30, 146–161. [Google Scholar] [CrossRef]
- Rusconi, C. Restos de huesos fósiles en una cueva de Malalhue (Mendoza). An. Soc. Cient. Arg. 1946, 141, 241–249. [Google Scholar]
- Neme, G.; Gil, A. Biogeografía humana en los Andes meridionales: Tendencias arqueológicas en el sur de Mendoza. Chungara Rev. Antropol. Chilena 2008, 40, 5–18. [Google Scholar]
- Pesquín, P.B. Los mamíferos pleistocenos de Santiago del Estero (Argentina), con énfasis en los Toxodontidae (Notoungulata). Bachelor’s Thesis, Facultad de Química, Bioquímica y Farmacia, Universidad Nacional de San Luis, San Luis, Argentina, 2010. [Google Scholar]
- García, M.E.; Bonini, R.; Alberdi, M.T.; Prado, J.L. First record of Equus neogeus from Abaucán River (Catamarca, Argentina). Est. Geol. 2018, 74, e080. [Google Scholar] [CrossRef]
- Sánchez, B.; Prado, J.L.; Alberdi, M.T. Ancient feeding, ecology and extinction of Pleistocene horses from the Pampean Region (Argentina). Ameghiniana 2006, 43, 427–436. [Google Scholar]
- Miño Boilini, A.R.; Cerdeño, E.; Bond, M. Revisión del género Toxodon Owen, 1837 (Notoungulata: Toxodontidae) en el Pleistoceno de las provincias de Corrientes, Chaco y Santa Fe, Argentina. Rev. Esp. Paleontol. 2006, 21, 93–103. [Google Scholar] [CrossRef]
- Prado, J.L.; Martínez-Maza, C.; Alberdi, M.T. Megafauna extinction in South America: A new chronology for the Argentine Pampas. Palaeogeogr. Palaeoclim. Palaeoecol. 2015, 425, 41–49. [Google Scholar] [CrossRef]
- Prates, L.; Perez, S.I. Late Pleistocene South American megafaunal extinctions associated with rise of fishtail points and human population. Nat. Comm. 2021, 12, 2175. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxa | San Luis | Mza. | La Pampa | Bs.As. | Cba. | NOA | NEA | Chaco | Stgo. E. |
|---|---|---|---|---|---|---|---|---|---|
| Neosclerocalyptus paskoensis | X | X | X | X | X | X | X | ||
| Panochthus tuberculatus * | cf. | X | X | X | X | X | |||
| Panochthus sp. | X | X | |||||||
| Glyptodon reticulatus | X | X | X | X | |||||
| Glyptodon clavipes | X | ||||||||
| Glyptodon sp. | X | X | |||||||
| Scelidotherium leptocephalum | X | X | X | X | X | X | |||
| S. bravardi ** | X | ||||||||
| Scelidotherium sp. | X | ||||||||
| Scelidotheriinae indet. | X | X | |||||||
| Catonyx chiliensis | X | X | |||||||
| Megatherium americanum | X | X | X | ||||||
| Megatherium sp. | X | X | |||||||
| Macrauchenia patachonica | X | cf. | X | X | X | X | |||
| Notiomastodon platensis | X | X | X | X | X | ||||
| Equus (A.) neogeus | X | cf. | X | X | cf. | ||||
| Equus (A.) sp. | X | X |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cerdeño, E.; Lucero, N.P.; Chiesa, J.O. Quaternary Mammals from Central-Western Argentina in the Stratigraphic Context of Southern South America. Quaternary 2023, 6, 59. https://doi.org/10.3390/quat6040059
Cerdeño E, Lucero NP, Chiesa JO. Quaternary Mammals from Central-Western Argentina in the Stratigraphic Context of Southern South America. Quaternary. 2023; 6(4):59. https://doi.org/10.3390/quat6040059
Chicago/Turabian StyleCerdeño, Esperanza, Natalia P. Lucero, and Jorge O. Chiesa. 2023. "Quaternary Mammals from Central-Western Argentina in the Stratigraphic Context of Southern South America" Quaternary 6, no. 4: 59. https://doi.org/10.3390/quat6040059