
The Fox from Bajiazui (Qingyang, Central China) and an Update on Early Pleistocene Foxes from China
 ,
,
Abstract
:1. Introduction
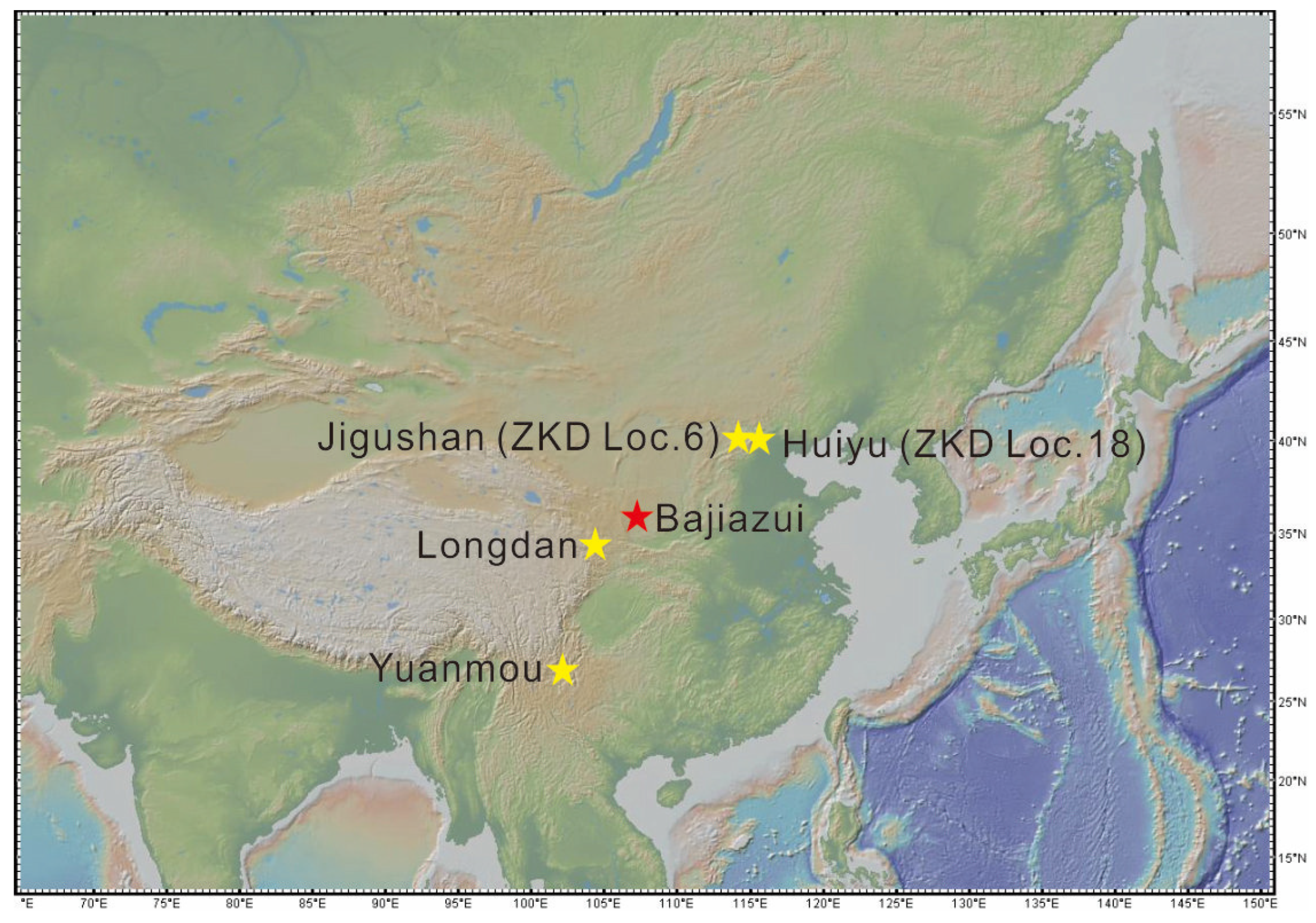
2. Geological and Geographical Background
3. Material and Methods
3.1. Abbreviations
3.1.1. Site and Institutional Abbreviations
3.1.2. Measurement Abbreviations
4. Systematic Paleontology
4.1. Referred Specimens
4.2. Description
4.3. Morphological Comparisons
4.4. Morphometric Comparisons
5. Discussion
Taxonomical Attribution of the Bajiazui Specimens and the Status of V. chikushanensis
6. Concluding Remarks
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lindblad-Toh, K.; Wade, C.M.; Mikkelsen, T.S.; Karlsson, E.K.; Jaffe, D.B.; Kamal, M.; Clamp, M.; Chang, J.L.; Kulbokas, E.J.; Zody, M.C.; et al. Genome Sequence, Comparative Analysis and Haplotype Structure of the Domestic Dog. Nature 2005, 438, 803–819. [Google Scholar] [CrossRef]
- Castelló, J.R. Canids of the World; Princeton University Press: Princeton, NJ, USA, 2018; ISBN 978-0-691-17685-7. [Google Scholar]
- Nowak, R.M. Walker’s Carnivores of the World; JHU Press: Baltimore, MD, USA, 2005; ISBN 0-8018-8032-7. [Google Scholar]
- Kaya, F.; Bibi, F.; Žliobaitė, I.; Eronen, J.T.; Hui, T.; Fortelius, M. The Rise and Fall of the Old World Savannah Fauna and the Origins of the African Savannah Biome. Nat. Ecol. Evol. 2018, 2, 241–246. [Google Scholar] [CrossRef]
- Tedford, R.H.; Wang, X.; Taylor, B.E. Phylogenetic Systematics of the North American Fossil Caninae (Carnivora: Canidae). Bull. Am. Mus. Nat. Hist. 2009, 325, 1–218. [Google Scholar] [CrossRef]
- Bartolini Lucenti, S.; Madurell-Malapeira, J. Unraveling the Fossil Record of Foxes: An Updated Review on the Plio-Pleistocene Vulpes spp. from Europe. Quat. Sci. Rev. 2020, 236, 106296. [Google Scholar] [CrossRef]
- De Bonis, L.; Peigné, S.; Likius, A.; Mackaye, H.T.; Vignaud, P.; Brunet, M. The Oldest African Fox (Vulpes riffautae n. sp., Canidae, Carnivora) Recovered in Late Miocene Deposits of the Djurab Desert, Chad. Naturwissenschaften 2007, 94, 575–580. [Google Scholar] [CrossRef]
- Wang, X.; Tseng, Z.J.; Li, Q.; Takeuchi, G.T.; Xie, G. From “third Pole” to North Pole: A Himalayan Origin for the Arctic Fox. Proc. R. Soc. Biol. Sci. 2014, 281, 20140893. [Google Scholar] [CrossRef]
- Qiu, Z.; Tedford, R.H. A Pliocene Species of Vulpes from Yushe, Shanxi. Vertebr. Palasiat. 1990, 28, 245–258. [Google Scholar]
- Bartolini-Lucenti, S.; Madurell-Malapeira, J. Ancestral Foxes at the Gates of Europe: The Pliocene Fox from Çalta-1 (Turkey) and their Relationships with Asian and European Plio-Pleistocene Foxes. Comptes Rendus Palevol 2021, 20, 619–626. [Google Scholar] [CrossRef]
- Bartolini-Lucenti, S. A New Large-Sized Pliocene Fox (Carnivora, Canidae) from Yushe Basin (Shanxi, China). Riv. Ital. Paleontol. Stratigr. 2021, 127, 133–147. [Google Scholar] [CrossRef]
- Odintzov, I.A. Vulpes praecorsac Kormos from Pliocene Deposits of Odessa. Paleontol. Sb. L’vov Univ. 1965, 2, 57–64. [Google Scholar]
- Spassov, N. Biochronology and Zoogeographic Affinities of the Villafranchian Faunas of Bulgaria and South Europe. Hist. Nat. Bulg. 2000, 12, 89–128. [Google Scholar]
- Teilhard de Chardin, P. The Fossils from Locality 18 near Peking. Palaeontol. Sin. New Ser. C 1940, 9, 1–95. [Google Scholar]
- Qiu, Z.; Deng, T.; Wang, B. Early Pleistocene Mammalian Fauna from Longdan, Dongxiang, Gansu, China. Palaeontol. Sin. New Ser. C 2004, 191, 1–245. [Google Scholar]
- Ginsburg, L. Le Gisement de Vertébrés Pliocenes de Çalta, Ankara, Turquie. 5. Carnivores. Geodiversitas 1998, 20, 379–396. [Google Scholar]
- Rodríguez, J.; Rodríguez-Gómez, G.; Martín-González, J.A.; Goikoetxea, I.; Mateos, A. Predator–Prey Relationships and the Role of Homo in Early Pleistocene Food Webs in Southern Europe. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 365–366, 99–114. [Google Scholar] [CrossRef]
- Lazarev, S.; Kuiper, K.F.; Oms, O.; Bukhsianidze, M.; Vasilyan, D.; Jorissen, E.L.; Bouwmeester, M.J.; Aghayeva, V.; van Amerongen, A.J.; Agustí, J.; et al. Five-Fold Expansion of the Caspian Sea in the Late Pliocene: New and Revised Magnetostratigraphic and 40Ar/39Ar Age Constraints on the Akchagylian Stage. Glob. Planet. Chang. 2021, 206, 103624. [Google Scholar] [CrossRef]
- Pei, W.-C. On the Carnivora from Locality 1 of Choukoutien. Palaeontol. Sin. Ser. C 1934, 8, 1–216. [Google Scholar]
- Li, Y.; Zhang, Y.; Wu, X.; Ao, H.; Li, L.; An, Z. Mammalian Evolution in Asia Linked to Climate Changes. In Late Cenozoic Climate Change in Asia; Developments in Paleoenvironmental Research; Springer: Dordrecht, The Netherlands, 2014; Volume 16, pp. 435–490. ISBN 978-94-007-7816-0. [Google Scholar]
- Sher, A.V. Olyorian Land Mammal Age of Northeastern Siberia. Palaeontogr. Ital. 1986, 74, 97–112. [Google Scholar]
- Deng, T.; Xue, X.-X. Equus qingyangensis sp. nov. (Perissodactyla, Equidae) from the Early Pleistocene of Qingyang, Gansu, China. Vertebr. Palasiat. 1999, 37, 62–74. [Google Scholar]
- Wang, J. Early Pleistocene Mammalian Fauna from Bajiazui, Qingyang, Gansu, China and Their Environment; Northwest University: Xi’an, China, 2006; pp. 1–88. [Google Scholar]
- Xue, X.-X.; Zhang, Y.-X.; Yue, L.-P. Paleoenvironments indicated by the fossil mammalian assemblages from red clay-loess sequence in the Chinese Loess Plateau since 8.0 Ma B.P. Sci. China Ser. D 2006, 49, 518–530. [Google Scholar] [CrossRef]
- Young, C.C. On the Mammalian Remains from Chi Ku Shan. Palaeontol. Sin. 1930, 8, 5–19. [Google Scholar]
- Rabeder, G. Die Carnivoren (Mammalia) Aus Dem Altpleistozän Mit Beiträgen Zur Systematik Einiger Musteliden Und Caniden. Beitr. Paläontol. Osterr. 1976, 1, 5–119. [Google Scholar]
- Garrido, G. El registro de Vulpes alopecoides (Forsyth-Major, 1877), Canis etruscus Forsyth-Major, 1877 y Canis cf. falconeri Forsyth-Major, 1877 (Canidae, Carnivora, Mammalia) en Fonelas P-1 (Cuenca de Guadix, Granada). Vertebr. Plioceno Super. Termin. El Suroeste Eur. Fonelas P-1 El Proy. Fonelas 2008, 10, 159–186. [Google Scholar]
- Koufos, G.D. The Villafranchian Carnivoran Guild of Greece: Implications for the Fauna, Biochronology and Paleoecology. Integr. Zool. 2014, 9, 444–460. [Google Scholar] [CrossRef]
- Bartolini-Lucenti, S.; Spassov, N. Cave Canem! The Earliest Canis (Xenocyon) (Canidae, Mammalia) of Europe: Taxonomic Affinities and Paleoecology of the Fossil Wild Dogs. Quat. Sci. Rev. 2022, 276, 107315. [Google Scholar] [CrossRef]
- Ebbestad, J.O.R.; Romgard, J. Otto Zdansky the Scientist Who Discovered Peking Man and Explored China’s Fossil Past; Acta Universitatis Upsaliensis: Uppsala, Sweden, 2021; ISBN 978-91-554-7182-8. [Google Scholar]
- Teilhard de Chardin, P.; Piveteau, J. Les Mammifères Fossiles de Nihowan (Chine). Ann. Paleontol. 1930, 19, 1–134. [Google Scholar]
- You, Y.; Qi, G. New Materials of Pleistocene Mammal Fossils in Yuanmou, Yunnan. Vertebr. Palasiat. 1973, 11, 66–85. [Google Scholar]
- Wang, X.; Flynn, L.J.; Fortelius, M. Fossil Mammals of Asia; Columbia University Press: New York, NY, USA, 2013; pp. 1–732. [Google Scholar]
- Gimranov, D.O.; Kosintsev, P.A.; Gasilin, V.V. Species Diagnostics of Corsac (Vulpes Corsac), Fox (Vulpes Vulpes), and Arctic Fox (Vulpes Lagopus) According to Ontological Characteristics of Mandible Teeth. Zool. Zhurnal 2015, 94, 1338–1350. [Google Scholar] [CrossRef]
- Gimranov, D.O. Species Diagnostics of the Corsac (Vulpes Corsac), Fox (Vulpes Vulpes) and Arctic Fox (Vulpes Lagopus, Carnivora, Canidae) Using the Upper Teeth. Zool. Journal 2017, 96, 684–697. [Google Scholar] [CrossRef]
- Kahlke, R.D.; García, N.; Kostopoulos, D.S.; Lacombat, F.; Lister, A.M.; Mazza, P.P.A.; Spassov, N.; Titov, V.V. Western Palaearctic Palaeoenvironmental Conditions during the Early and Early Middle Pleistocene Inferred from Large Mammal Communities, and Implications for Hominin Dispersal in Europe. Quat. Sci. Rev. 2011, 30, 1368–1395. [Google Scholar] [CrossRef]
- Head, M.J.; Gibbard, P.L. Early-Middle Pleistocene Transitions: Linking Terrestrial and Marine Realms. Quat. Int. 2015, 389, 7–46. [Google Scholar] [CrossRef]
- Madurell-Malapeira, J.; Ros-Montoya, S.; Espigares, M.P.; Alba, D.M.; Aurell-Garrido, J. Villafranchian Large Mammals from the Iberian Peninsula: Paleobiogeography, Paleoecology and Dispersal Events. J. Iber. Geol. 2014, 40, 167–178. [Google Scholar] [CrossRef]
- Rook, L.; Martínez-Navarro, B. Villafranchian: The Long Story of a Plio-Pleistocene European Large Mammal Biochronologic Unit. Quat. Int. 2010, 219, 134–144. [Google Scholar] [CrossRef]
- Antón, M.; Turner, A.; Salesa, M.J.; Morales, J. A Complete Skull of Chasmaporthetes Lunensis (Carnivora, Hyaenidae) from the Spanish Pliocene Site of La Puebla de Valverde (Teruel). Estud. Geol. 2006, 62, 375–388. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C L | C W | P1 L | P1 W | P2 L | P2 W | P3 L | P3 W | P4 L | P4 W | M1 L | M1 W | M2 L | M2 W | LMR | C-M2 L | P4-M2 L | GPW | ECW | |
| MLS028 | 5.10 | 3.40 | 3.80 | 2.30 | 6.80 | 2.60 | 7.7 | 3.2 | 12.2 | 6.3 | 8.2 | 10.9 | 5.7 | 8.5 | 13.65 | 55.90 | 23.40 | 37.70 | 19.00 |
| c L | c W | p2 L | p2 W | p3 L | p3 W | p4 L | p4 W | m1 L | m1 W | m1td L | m1td W | m1trd L | m2 L | m2 W | |||||
| M63022 | 5.6 | 3.4 | 6.8 | 2.6 | 7.3 | 2.8 | 8.2 | 3.4 | 14 | 5.2 | 4.2 | 5 | 9.8 | 6.5 | 5.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bartolini-Lucenti, S.; Jiangzuo, Q.; Jiang, H.; Li, Z.; Xie, K.; Madurell-Malapeira, J. The Fox from Bajiazui (Qingyang, Central China) and an Update on Early Pleistocene Foxes from China. Quaternary 2023, 6, 57. https://doi.org/10.3390/quat6040057
Bartolini-Lucenti S, Jiangzuo Q, Jiang H, Li Z, Xie K, Madurell-Malapeira J. The Fox from Bajiazui (Qingyang, Central China) and an Update on Early Pleistocene Foxes from China. Quaternary. 2023; 6(4):57. https://doi.org/10.3390/quat6040057
Chicago/Turabian StyleBartolini-Lucenti, Saverio, Qigao Jiangzuo, Hao Jiang, Zhaoyu Li, Kun Xie, and Joan Madurell-Malapeira. 2023. "The Fox from Bajiazui (Qingyang, Central China) and an Update on Early Pleistocene Foxes from China" Quaternary 6, no. 4: 57. https://doi.org/10.3390/quat6040057