
There Was No Mesozoic Marine Revolution †


1
New Mexico Museum of Natural History, 1801 Mountain Road N. W., Albuquerque, NM 87104, USA
2
Flying Heritage and Combat Armor Museum, 3407 109th St SW, Everett, WA 98204, USA
*
Author to whom correspondence should be addressed.
†
Presented at the 4th International Electronic Conference on Geosciences, 1–15 December 2022; Available online: https://sciforum.net/event/IECG2022 .
Proceedings 2023, 87(1), 87; https://doi.org/10.3390/IECG2022-14819
Published: 21 July 2023
(This article belongs to the Proceedings of The 4th International Electronic Conference on Geosciences)
{kind=link}
Abstract
:The phrase “Mesozoic marine revolution” refers to the Mesozoic origin of durophagous predators and the co-evolutionary response of their prey as well as an increase in infaunalization. However, using “revolution” for a process that takes many tens of millions of years is semantically improper hyperbole. Durophagous predators and their prey began to co-evolve by the Devonian, continued into the late Cenozoic and encompassed many distinct and convergent evolutionary events. Infaunalization has a similar prolonged and complex history. Identifying a single “revolution” confounds understanding of the multiple events and evolutionary convergences that took place, so “Mesozoic marine revolution” should be abandoned.
1. Introduction
Vermeij [1] introduced the phrase “Mesozoic marine revolution” (MMR) to refer primarily to the Cretaceous origin of durophagous (“shell-destroying”) predators such as teleost fishes and decapod crustaceans and the co-evolutionary response of their prey (primarily gastropods) by increasing shell sturdiness. He also drew attention to a perceived simultaneous increase in benthic grazing and a tendency toward infaunalization. Indeed, the MMR has come to be seen as a sweeping restructuring of benthic communities and one of the most significant paleoecological events of the Mesozoic. This “revolution” thus is identified as the origin of significant durophagous predators (such as shell-boring gastropods) and an intensification in grazing that resulted in a substantial increase in the sturdiness of bivalve shells, and the turnover from marine benthic communities dominated by epifaunal (surface-dwelling) or semi-infaunal animals to a more infaunal benthos [1,2,3].
Subsequent work, however, pushed the onset of these changes back into the Triassic or Jurassic, so that the MMR is now perceived to have taken place during most or all of the Mesozoic. Furthermore, durophagous predators had already evolved during the Paleozoic, in the Middle Devonian, and their prey items may have also responded co-evolutionarily to increase shell sturdiness [4]. Infaunalization also began during the late Paleozoic.
Here, we question identifying a single “revolution” or “event” in the co-evolution of durophagous predators and their prey and the infaunalization of the marine benthos. We begin by noting that the use of “revolution” for an event that takes tens of millions of years is simply semantically improper. We then discuss the evidence that durophagous predators and their prey began to co-evolve in the Devonian (if not earlier), and that such co-evolution continued into the late Cenozoic. Durophagy evolved in diverse taxa that evolved a disparate range of anatomical structures for durophagy. Infaunalization also has a similar prolonged and complex history. Identifying a single “revolution” thus confounds understanding of the multiple events and evolutionary convergences that actually took place. Therefore, the phrase and concept “Mesozoic marine revolution” needs to be abandoned. We also note that the concept of escalation, the co-evolution of predators and prey, is open to question, but discussion of this is beyond our scope here.
2. What Is a Revolution?
The term revolution is now popular in paleontology to refer to dramatic events/changes in the history of life. Thus, there is the Ordovician plankton revolution, the Paleozoic marine revolution, the Paleozoic phytoplankton revolution, the Mesozoic marine revolution and the Mesozoic terrestrial revolution, among others. Some similar “events” have different suffixes, such as the Cambrian explosion and the great Ordovician biodiversity event (which even has its own acronym, GOBE). Each of these “revolutions” and “events” took place over the course of millions or tens of millions of years.
So, there is a semantic problem that needs to be resolved here; namely, the proper and improper uses of the words “revolution” and “event”. The Merriam Webster English-language dictionary provides a general definition of a revolution as “a sudden, radical, or complete change”. Historical revolutions, such as the Bolshevik Revolution of 1917, are short-lived events that usually last much less than one year. Prolonged historical events that occur over several years, decades or longer, are usually not referred to as revolutions (an exception is the French Revolution). Merriam Webster defines the word event as “something that happens” or “a noteworthy happening”. Clearly, an event is a singular, one, “happening”, and it is not applicable to a process that lasted for millions of years, which must encompass events (plural) and not an event (singular).
Therefore, in the fossil record, why would processes that took the entire Cretaceous Period (about 80 million years) or longer be referred to as either an “event” or a “revolution?” Furthermore, an “event” that lasted the entire Mesozoic, about 186 million years, also does not merit the appellation “revolution”. These improper uses of the word revolution and event are intended to draw attention to the events and, we believe, hyperbolize them. However, if a perceived evolutionary “process” is a series of events, many not related, over many millions of years, then the label “revolution” misleads and masks important evolutionary history.
Recognizing the semantic issues, Vermeij [1], p. 245, stated that the “revolutions” of the fossil record “are not the instantaneous take-overs or inventions with which we identify revolutions in human history, but lasted tens of millions of years; yet these episodes are short relative to the hundreds of millions of years when comparatively little fundamental change took place in community organization”. However, this statement assumes without justification that there were long periods of little change in community organization punctuated by shorter “fundamental” changes. We read the fossil record differently, as one in which the fundamental changes in community organization have been multiple, long term and/or ongoing, often cumulative, changes. This reading finds substantiation below.
3. How Long Was the Mesozoic Marine “Revolution?”
Sepkoski [5] introduced the idea of a Paleozoic Evolutionary Fauna (PEF) in which brachiopods and crinoids dominated seafloors, followed by the Modern Evolutionary Fauna (MEF) in which molluscs dominated seafloors. In the marine benthos, the shift from the PEF to the MEF was the transition from sedentary epifaunal suspension feeders to mobile, energetic infaunal suspension feeders, deposit feeders and predators [6]. The MMR has been seen as a critical step in the change from the PEF to MEF. However, it is clear that this change began in the late Paleozoic [6] and was heavily influenced by the end Permian extinctions [7]. For example, there were benthic molluscs (gastropods and bivalves) by the Pennsylvanian and Permian, and infaunalization, even in nonmarine settings, began during the late Paleozoic [8,9].
When Vermeij [1] proposed the MMR, the data available to him suggested it was primarily a Cretaceous event. However, subsequent data and analyses have pushed back the onset of the MMR to either the Triassic or Jurassic according to various workers, including Vermei [2]. And there is a so-called “precursor” in the middle Paleozoic.
4. Paleozoic Events
The oldest evidence of durophagy may be an early Cambrian trilobite with an injury to its carapace that it likely incurred during ontogeny [10]. A diversity of Paleozoic durophages includes Late Ordovician durophagous arthropods and an impressive radiation of mid-Paleozoic durophagous predators (especially fishes) that took place over at least 25 Ma during the Devonian–Carboniferous [4].
Signor and Brett [4] analyzed changes in marine predation patterns during the Devonian–Carboniferous, and some credit them with the term “Mid-Palaeozoic Revolution” (MPR) [11] or “Middle Paleozoic Marine Revolution” (MPMR) [12], though they did not use either term. Brett et al. [13], p. 98, introduced the term Middle Paleozoic Revolution (MPR), and Baumiller and Gahn [14] used the term Middle Paleozoic Marine Revolution (MPMR). However, having taken at least 25 Ma, we reject referring to these “events” as a “revolution”.
Signor and Brett [4], p. 229, observed that “the mid-Paleozoic [Devonian] was punctuated by a rapid radiation of durophagous (shell-crushing) predators” and noted that there was also “an increase in the frequency of predator-resistant morphologies in a variety of marine invertebrate taxa”. Indeed, identifying a marked increase in durophagous fish during the later Devonian and Carboniferous [4], Figure 1, is reinforced by an increase in the record of angular digestilites of invertebrates in the Mississippian [12,15].
Signor and Brett [4] called this mid-Paleozoic diversification of durophages and the inferred co-evolutionary response a “Paleozoic precursor” to the MMR. However, a precursor means a “forerunner, harbinger or herald”, so there is some causal connection between two events. But how can these Paleozoic events, in totally different clades than the Mesozoic counterparts and separated by more than tens of millions of years from the Mesozoic, be “precursors?” Thus, as Buatois et al. [3], p. 61, stated, the mid-Paleozoic “event” is “a distinct episode, whose link with the subsequent MMR is tenuous at best”. Nevertheless, it shows the possible co-evolution of durophages and their prey has a much older history than the Mesozoic, as does the record of infaunalization [8,16].
5. Triassic Events
Saloman et al. [17] described regurgitalites from the Middle Triassic Gogolin Formation in Southern Poland that include angular bivalve shell fragments with sharp, non-abraded, margins and crinoid ossicles with many breaks. These are inferred to be the regurgitalites of durophagous vertebrates [17]. Saloman et al. [17] concluded that these regurgitalites indicate that the MMR may have begun by the Middle Triassic, and Early Triassic coprolites have been used to suggest that the MMR began even earlier in the Triassic [18,19,20], as is also suggested by evidence from the body fossils of marine reptiles [21,22]. The record of crinoids also suggests significant predation by sea urchins began during the Triassic [23,24]. Indeed, there were diverse Early–Middle Triassic durophages, including the first lobsters, decapod crustaceans, various fishes and marine reptiles.
Triassic ichnofaunas show limited infaunalization and relatively simple tiering structure. So, “….the Triassic is better regarded as a prelude to the MMR, rather than part of this evolutionary breakthrough” [3], p. 63. However, we read the trace fossil record as indicating infaunalization began in the Paleozoic, experienced a major reset by the end Permian extinctions, followed by increasing infaunalization throughout the Mesozoic and much of the Cenozoic, which is a process that took place in diverse clades employing disparate behavioral strategies over many tens of million years—not a revolution.
6. Jurassic-Cretaceous Events
Hunt and Lucas [15] summarized evidence from ichnology, including regurgitalites [11,25], consumulites [26,27] and digestilites [28], for increased durophagous vertebrate predation on invertebrates in the Jurassic and Cretaceous. However, ichthyosaur consumulites demonstrate that some feeding changes already took place during the Triassic, as Early and Middle Triassic specimens contain only cephalopod hooklets, whereas Late Triassic consumulites include both vertebrate remains and mollusk shell fragments [29,30,31,32,33,34]. This corresponds to an ecomorphological diversification of marine vertebrate durophages also documented by skeletal material [22].
According to Buatois and Mángano [35] (also see [3]), the MMR began in the Jurassic with an increase in durophagous predation and infaunalization, though some relevant innovation began in the Late Triassic [2,3,36,37]. Finnegan et al. [36] concluded that the energy budgets of marine ecosystems increased during the Jurassic–Cretaceous, though their own data show that the increases began in the Middle–Late Triassic. Vermeij [1] acknowledged that infaunalization of the soft-bottom benthos began during the Paleozoic but claimed that what happened in the Cretaceous was dramatic: a Mesozoic community reorganization. However, that reorganization began in the Triassic (if not before), continued through the Neogene [3], and there is no discrete infaunalization event that could be called a “revolution”.
7. Cenozoic Events
The Cenozoic saw the appearance of new clades of more powerful durophages such as crabs, rays and certain teleosts. The Neogene saw the establishment of modern marine ecosystems and more complex trace fossil tiering [3,16]. Processes deemed characteristic of the MMR thus continued and culminated during the late Cenozoic.
8. One Event or Multiple Events?
The preceding brief review indicates that the co-evolution of durophages and their prey and major changes in benthic communities took place as multiple events in diverse clades from the Devonian (or earlier) to the Neogene [38,39,40,41]. Thus, “in actuality, the Mesozoic and Cenozoic evolution of predators involved a series of episodes” [16], p. 119. Indeed, Vermeij [1] listed various modes of molluscivory including crushing (first seen in Devonian dipnoans) and extraoral/intraoral digestions by asteroids that began in the Late Ordovician. So, the MMR is singling out part of what was a lengthy and complex process, including the independent origins of many diverse and disparate durophages over hundreds of millions of years and the inferred co-evolutionary response of their supposed prey items.
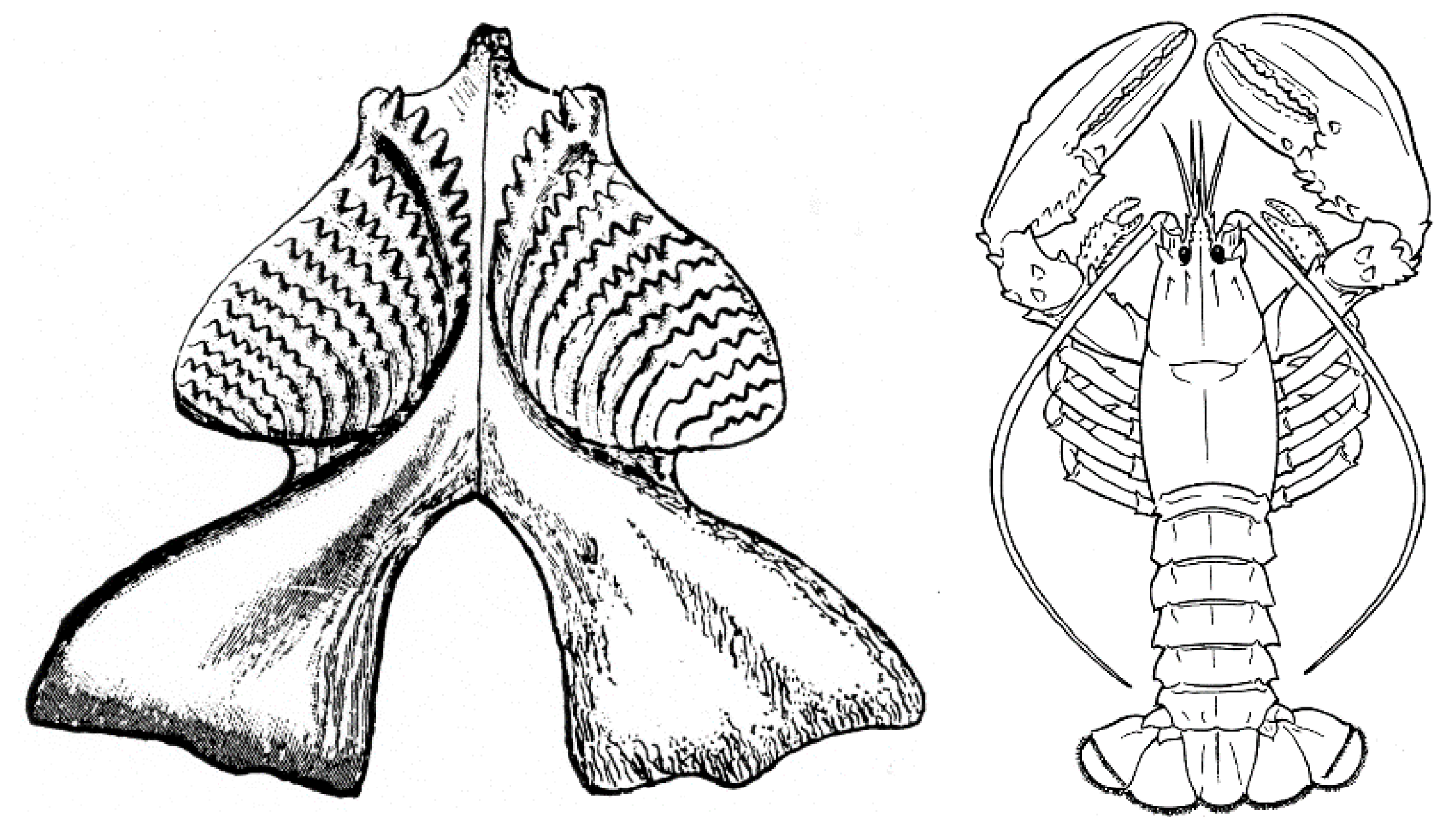
Many of the durophagous vertebrates convergently evolved broad, flat or rounded dental pavements for crushing (Figure 1), but other means of crushing evolved; for example, the massive jaws of some turtles, which lack teeth. And, among crustaceans, other methods of crushing, such as claws, evolved (Figure 1). To combine all of these durophages into a single “revolution” conflates multiple evolutionary events in diverse clades with disparate morphological structures for durophagy into a single event of tens of millions of years in duration.
Figure 1.
Two contrasting solutions to durophagy. Left, Carboniferous lungfish lower jaw and toothplate from [42], which is used to crush food, in occlusal view. Right, extant lobster, which uses its claws to crush food (from Wikimedia Commons).
Figure 1.
Two contrasting solutions to durophagy. Left, Carboniferous lungfish lower jaw and toothplate from [42], which is used to crush food, in occlusal view. Right, extant lobster, which uses its claws to crush food (from Wikimedia Commons).

According to Walker and Brett [16], p. 119, the evolution of durophagous predators during the Mesozoic and Cenozoic involved “a series of episodes”, which they described as “episodic but generally increasing predation pressure on marine organisms through the Mesozoic-Cenozoic interval”. They show many phases of “escalation” [16], Figure 14, and refer to Cambrian, mid-Paleozoic, Mesozoic and Cenozoic marine revolutions, each of which took tens to millions of years. However, we question calling these revolutions for reasons already stated.
Buatois et al. [3], Figure 9.36, illustrated “representative tiering structures and ichnoguilds from selected case studies illustrating infaunal colonization during and in the aftermath of the MMR”. These are Triassic, Jurassic, Cretaceous, Eocene and Miocene examples of tiering, each with varied taxa and ichnoguilds in diverse facies, and the only apparent trend is the increasing numbers of taxa per ichnoguild evident in the Miocene. Clearly, tiering/infaunalization has a complex history of diverse taxa colonizing varied environments in different ways at different times. Conflating these as a single MMR combines multiple events that began in the Paleozoic. Indeed, if we examine the compilations of Buatois et al. [3] of “common bioturbating ichnogenera” (Table 9.1) and of “bioeroders” (Table 9.2), more than half of these ichnogenera (35 of 61 bioturbators, 14 of 26 bioeroders) have Ediacaran/Paleozoic first appearances. This does not support the conclusion that “the MMR strongly shaped the nature of animal–substrate interactions in the post-Paleozoic world” [3], p. 60.
9. Conclusions
No single event can be called the MMR. Levels of predation and infaunalization increased during the Mesozoic as part of a lengthy set of processes that began in the Paleozoic and, over many tens of millions of years by multiple convergent evolutionary events, restructured benthic communities. Durophagy first evolved in the Paleozoic and continued to appear in diverse taxa that evolved disparate morphological adaptations to durophagy through the Neogene. When you combine multiple events into one, as the concept of the MMR does, you conceal complexity. Thus, not only is the term MMR simply semantically misleading, it also embodies a view of the history of the marine biota that is not matched by the fossil record.
Supplementary Materials
The poster presentation can be downloaded at: https://www.mdpi.com/article/10.3390/IECG2022-14819/s1.
Author Contributions
Conceptualization, S.G.L. and A.P.H.; methodology, S.G.L. and A.P.H.; investigation, S.G.L. and A.P.H. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Vermeij, G.J. The Mesozoic marine revolution; evidence from snails, predators and grazers. Paleobiology 1977, 3, 245–258. [Google Scholar] [CrossRef]
- Vermeij, G.J. Escalation and its role in Jurassic biotic history. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 263, 3–8. [Google Scholar] [CrossRef]
- Buatois, L.A.; Carmona, N.B.; Curran, H.A.; Netto, R.G.; Mángano, M.G.; Wetzel, A. The Mesozoic marine revolution. In The Trace-Fossil Record of Major Evolutionary Events. Volume 2: Mesozoic and Cenozoic; Mángano, M.G., Buatois, L.A., Eds.; Topics in Geobiology 40; Springer: Dordrecht, The Netherlands, 2016; pp. 19–134. [Google Scholar]
- Signor, P.W., III; Brett, C.E. The mid-Paleozoic precursor to the Mesozoic marine revolution. Paleobiology 1984, 10, 229–245. [Google Scholar] [CrossRef]
- Sepkoski, J.J., Jr. A factor analytic description of the Phanerozoic marine fossil record. Paleobiology 1981, 7, 36–53. [Google Scholar] [CrossRef]
- Leighton, L.R.; Webb, A.E.; Sawyer, J.A. Ecological effects of the Paleozoic-Modern faunal transition: Comparing predation on Paleozoic brachiopods and molluscs. Geology 2013, 41, 275–278. [Google Scholar] [CrossRef]
- Wagner, P.J.; Kosnik, M.A.; Lidgard, S. Abundance distributions imply elevated complexity of post-Paleozoic marine ecosystems. Science 2006, 314, 1289–1292. [Google Scholar] [CrossRef]
- Ausich, W.I.; Bottjer, D.J. Tiering in suspension-feeding communities on soft substrata throughout the Phanerozoic. Science 1982, 216, 173–174. [Google Scholar] [CrossRef]
- Minter, N.J.; Buatois, L.A.; Mángano, M.G.; Davies, N.S.; Gibling, M.R.; Labandeira, C. The establishment of continental ecosystems. In The Trace-Fossil Record of Major Evolutionary Events. Volume 2: Mesozoic and Cenozoic; Mángano, M.G., Buatois, L.A., Eds.; Topics in Geobiology 40; Springer: Dordrecht, The Netherlands, 2016; pp. 205–324. [Google Scholar]
- Pocock, K.J. A unique case of teratology in trilobite segmentation. Lethaia 1974, 7, 63–66. [Google Scholar] [CrossRef]
- Borszcz, T.; Zatoń, M. The oldest record of predation on echinoids: Evidence from the Middle Jurassic of Poland. Lethaia 2013, 46, 141–145. [Google Scholar] [CrossRef]
- Salamon, M.A.; Gorzelak, P.; Niedźwiedzki, R.; Trzęsiok, D.; Baumiller, T.K. Trends in shell fragmentation as evidence of mid-Paleozoic changes in marine predation. Paleobiology 2014, 40, 14–23. [Google Scholar] [CrossRef]
- Brett, C.E.; Walker, S.E.; Kowalewski, M.; Kelley, P.H. Predators and predation in Paleozoic marine environments. Paleontol. Soc. Pap. 2002, 8, 93–118. [Google Scholar] [CrossRef]
- Baumiller, T.K.; Gahn, F.J. Testing predator-driven evolution with Paleozoic crinoid arm regeneration. Science 2004, 305, 1453–1455. [Google Scholar] [CrossRef]
- Hunt, A.P.; Lucas, S.G. The ichnology of vertebrate consumption: Dentalites, gastroliths and bromalites. New Mex. Mus. Nat. Hist. Sci. Bull. 2021, 87, 1–216. [Google Scholar]
- Walker, S.E.; Brett, C.E. Post-Palaeozoic patterns in marine predation: Was there a Mesozoic and Cenozoic marine predatory revolution? Palaeont. Soc. Pap. 2002, 8, 119–193. [Google Scholar] [CrossRef]
- Salamon, M.A.; Niedźwiedzki, R.; Gorzelak, P.; Lach, R.; Surmik, D. Bromalites from the Middle Triassic of Poland and the rise of the Mesozoic Marine Revolution. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2012, 321, 142–150. [Google Scholar] [CrossRef]
- Nakajima, Y.; Izumi, K. Coprolites from the upper Osawa Formation (upper Spathian), northeastern Japan: Evidence for predation in a marine ecosystem 5 Myr after the end-Permian mass extinction. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 414, 225–232. [Google Scholar] [CrossRef]
- Brachaniec, T.; Niedźwiedzki, R.; Surmik, D.; Krzykawski, T.; Szopa, K.; Gorzelak, P.; Salamon, M.A. Coprolites of marine vertebrate predators from the Lower Triassic of southern Poland. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2015, 435, 118–126. [Google Scholar] [CrossRef]
- Antczak, M.; Ruciński, M.R.; Stachacz, M.; Matysik, M.; Król, J.J. Diversity of vertebrate remains from the Lower Gogolin Beds (Anisian) of southern Poland. Ann. Soc. Geol. Pol. 2020, 90, 419–433. [Google Scholar] [CrossRef]
- Hu, S.X.; Zhang, Q.Y.; Chen, Z.Q.; Zhou, C.Y.; Lü, T.; Xie, T.; Wen, W.; Huang, J.Y.; Benton, M.J. The Luoping biota: Exceptional preservation, and new evidence on the Triassic recovery from end-Permian mass extinction. Proc. R. Soc. Lond. B 2011, 278, 2274–2282. [Google Scholar] [CrossRef] [PubMed]
- Stubbs, T.L.; Benton, M.J. Ecomorphological diversifications of Mesozoic marine reptiles: The roles of ecological opportunity and extinction. Paleobiology 2016, 42, 547–573. [Google Scholar] [CrossRef]
- Baumiller, T.K.; Salamon, M.A.; Gorzelak, P.; Mooi, R.; Messing, C.G.; Gahn, F.J. Post-Paleozoic crinoid radiation in response to benthic predation preceded the Mesozoic marine revolution. Proc. Nat. Acad. Sci. USA 2010, 107, 5893–5896. [Google Scholar] [CrossRef] [PubMed]
- Gorzelak, P.; Salamon, M.A.; Baumiller, T.K. Predator-induced macroevolutionary trends in Mesozoic crinoids. Proc. Nat. Acad. Sci. USA 2012, 109, 7004–7007. [Google Scholar] [CrossRef] [PubMed]
- Zatoń, M.; Villier, L.; Salamon, M.A. Signs of predation in the Middle Jurassic of south-central Poland: Evidence from echinoderm taphonomy. Lethaia 2007, 40, 139–151. [Google Scholar] [CrossRef]
- Pollard, J.E. The gastric contents of an ichthyosaur from the lower Lias of Lyme Regis, Dorset. Paleobiology 1968, 11, 376–388. [Google Scholar]
- Přikryl, T.; Košťk, M.; Mazuch, M.; Mikuláš, R. Evidence for fish predation on a coleoid cephalopod from the Lower Jurassic Posidonia Shale of Germany. Neues Jahrb. Geol. Palaontol.-Abh. 2012, 263, 25–33. [Google Scholar] [CrossRef]
- Oji, T.; Ogaya, C.; Sato, T. Increase of shell-crushing predation recorded in fossil shell fragmentation. Paleobiology 2003, 29, 520–526. [Google Scholar] [CrossRef]
- Rieber, H. Phragmoteuthis? ticinensis n. sp., ein Coleoidea-Rest aus der Grenzbitumenzone (Mittlere Trias) des Monte San Giorgio (Kt. Tessin, Schweiz). Paläontol. Z. 1970, 44, 32–40. [Google Scholar] [CrossRef]
- Camp, C.L. Large ichthyosaurs from the Upper Triassic of Nevada. Palaeontogr. Abt. A 1980, 170, 139–200. [Google Scholar]
- Brinkmann, W. Mixosaurier (Reptilia, Ichthyosauria) mit Quetschzähnen aus der Grenzbitumenzone (Mitteltrias) des Monte San Giorgio (Schweiz, Kanton Tessin). Schweiz. Paläontol. Abhand. 2004, 124, 1–84. [Google Scholar]
- Buchy, M.C.; Taugourdeau, P.; Janvier, P. Stomach contents of a Lower Triassic ichthyosaur from Spitzbergen. Oryctos 2004, 5, 47–55. [Google Scholar]
- Cheng, L.; Wings, O.; Chen, X.; Sander, P.M. Gastroliths in the Triassic ichthyosaur Panjiangsaurus from China. J. Paleontol. 2006, 80, 583–588. [Google Scholar]
- Druckenmiller, P.S.; Kelley, N.; Whalen, M.T.; McRoberts, C.; Carter, J.G. An Upper Triassic (Norian) ichthyosaur (Reptilia, Ichthyopterygia) from northern Alaska and dietary insight based on gut contents. J. Vert. Paleontol. 2014, 34, 1460–1465. [Google Scholar] [CrossRef]
- Buatois, L.; Mángano, M.G. The other biodiversity record: Innovation in animal-substrate interactions through geologic time. GSA Today 2018, 28, 4–10. [Google Scholar] [CrossRef]
- Finnegan, S.; McClain, C.M.; Kosnik, M.A.; Payne, J.L. Escargots through time: An energetic comparison of marine gastropod assemblages before and after the Mesozoic marine revolution. Paleobiology 2011, 37, 252–269. [Google Scholar] [CrossRef]
- Radley, J.D. Grazing erosion in Jurassic seas: A neglected factor in the Mesozoic marine revolution? Hist. Biol. 2010, 22, 387–393. [Google Scholar] [CrossRef]
- Thayer, C.W. Biological bulldozers and the evolution of marine benthic communities. Science 1979, 203, 458–461. [Google Scholar] [CrossRef]
- Bambach, R.K. Ecospace utilization and guilds in marine communities through the Phanerozoic. In Biotic Interactions in Recent and Fossil Benthic Communities; Tevesz, M.J.S., McCall, P.L., Eds.; Springer: Plenum, NY, USA, 1983; pp. 719–746. [Google Scholar]
- Bambach, R.K. Classes and adaptive variety: The ecology of diversification in marine faunas through the Phanerozoic. In Phanerozoic Diversity Patterns; Valentine, J.W., Ed.; Princeton University Press: Princeton, NJ, USA, 1985; pp. 191–253. [Google Scholar]
- Bush, A.M.; Bambach, R.K.; Daley, G.M. Changes in theoretical ecospace utilization in marine fossil assemblages between the mid-Paleozoic and late Cenozoic. Paleobiology 2007, 33, 76–97. [Google Scholar] [CrossRef]
- Peyer, B. Comparative Odontology; University of Chicago Press: Chicago, IL, USA, 1968; 347p. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lucas, S.G.; Hunt, A.P. There Was No Mesozoic Marine Revolution. Proceedings 2023, 87, 87. https://doi.org/10.3390/IECG2022-14819
AMA Style
Lucas SG, Hunt AP. There Was No Mesozoic Marine Revolution. Proceedings. 2023; 87(1):87. https://doi.org/10.3390/IECG2022-14819
Chicago/Turabian StyleLucas, Spencer G., and Adrian P. Hunt. 2023. "There Was No Mesozoic Marine Revolution" Proceedings 87, no. 1: 87. https://doi.org/10.3390/IECG2022-14819