
The Ongoing Epidemic of West Nile Virus in Greece: The Contribution of Biological Vectors and Reservoirs and the Importance of Climate and Socioeconomic Factors Revisited

Abstract
:1. West Nile Virus: Pathogenicity and Virulence
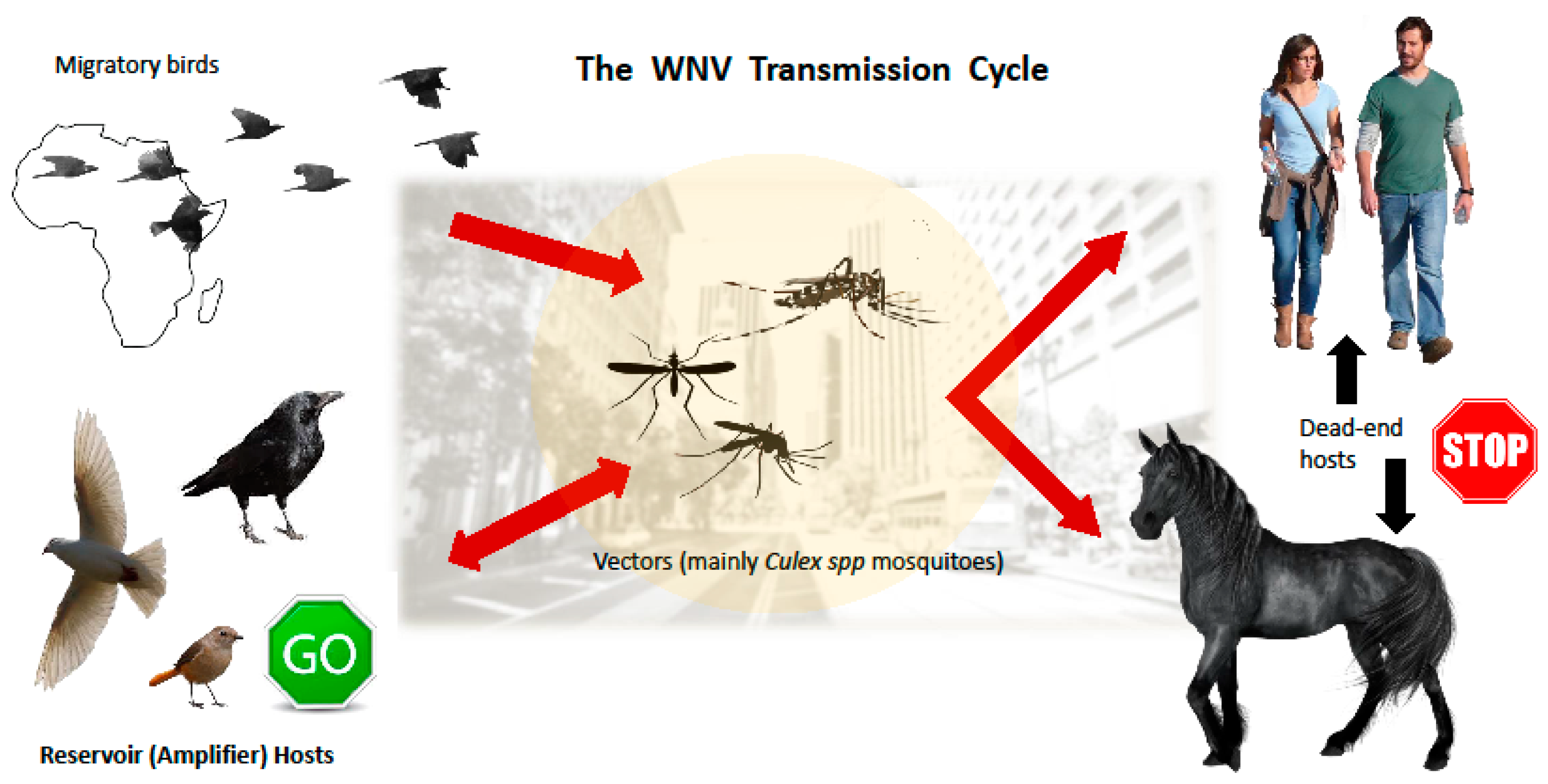
2. Transmission Vectors, Life Cycle and Animal Hosts of WNV
3. WNV Disease: An Emerging or Re-Emerging Infectious Disease?
4. Climate Factor Deviations and Mosquito-Borne Disease Expansion
5. Contribution of the Newly Translocated Arthropod Vectors to WNV Outbreaks in Europe
6. The Impact of Socioeconomic Factors on Vector-Borne Diseases
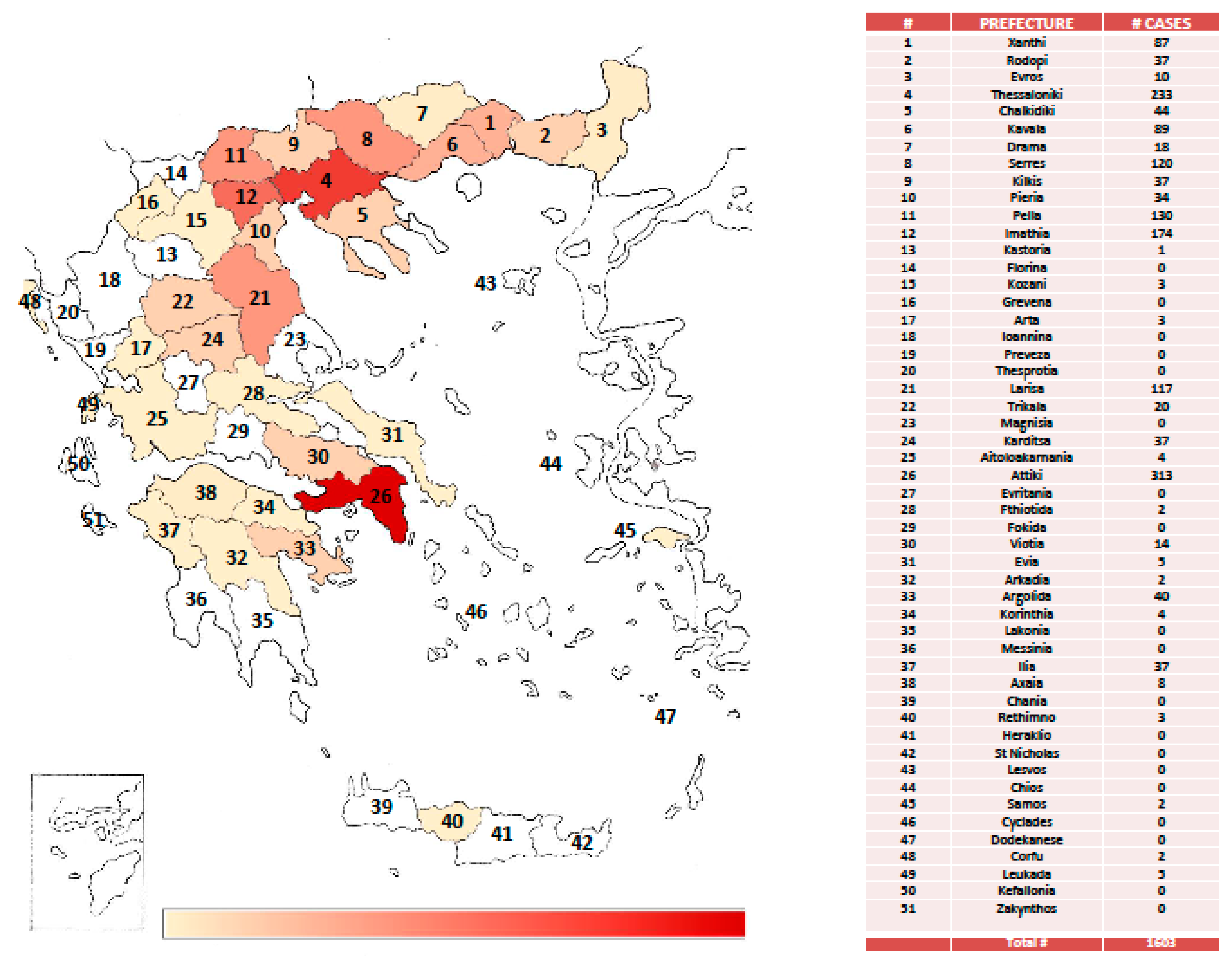
7. West Nile Virus: An Endemic Pathogen in Greece
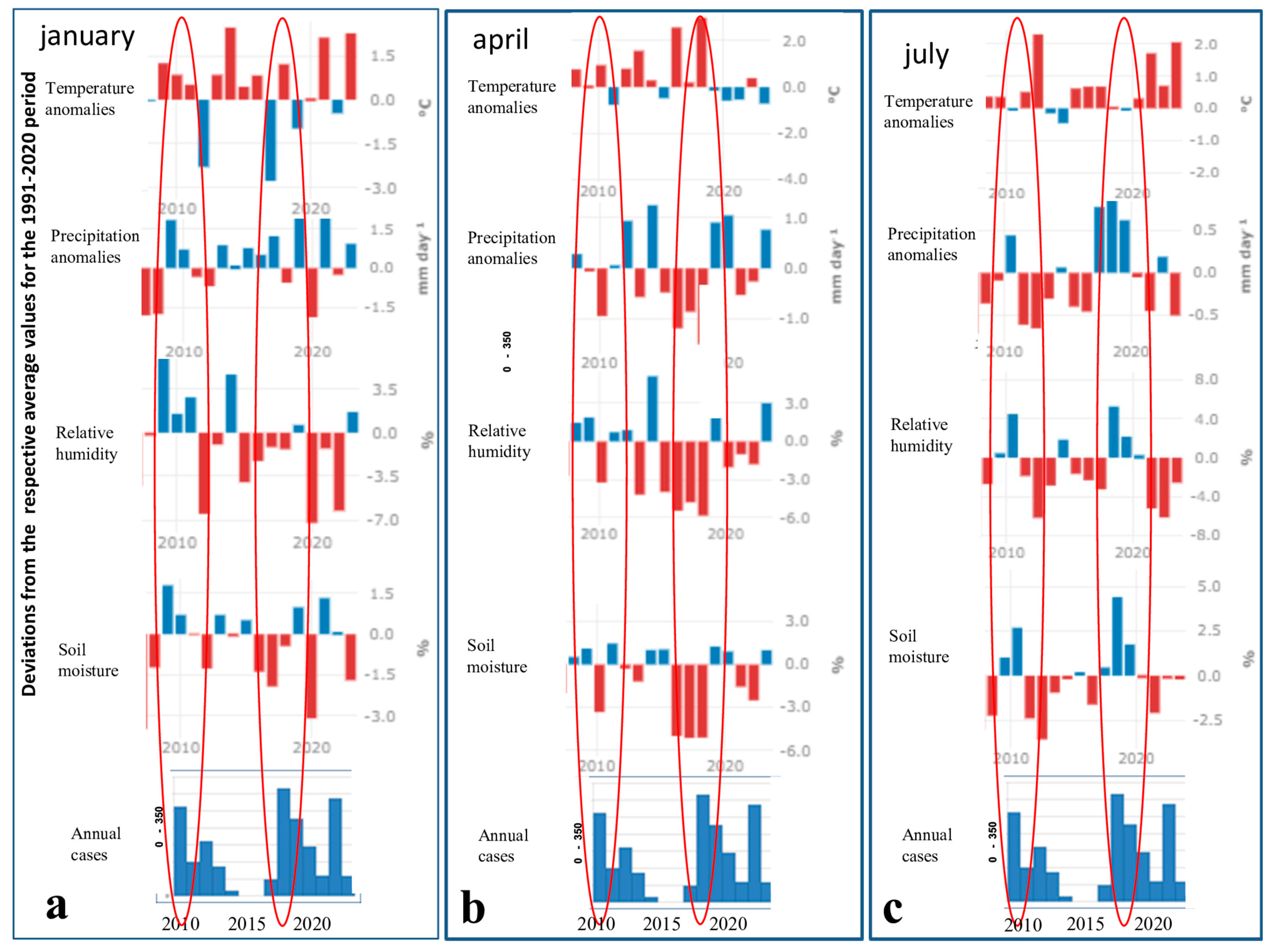
8. Comparison of Climatic Anomalies, Hydrological Variables, and Annual WNF Outbreaks in Greece
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Rossi, S.L.; Ross, T.M.; Evans, J.D. West Nile Virus. Clin. Lab. Med. 2010, 30, 47–65. [Google Scholar] [CrossRef] [PubMed]
- Brinton, M.A. The molecular biology of West Nile Virus: A new invader of the western hemisphere. Annu. Rev. Microbiol. 2002, 56, 371–402. [Google Scholar] [CrossRef] [PubMed]
- Saiz, J.-C.; Martín-Acebes, M.A.; Blázquez, A.B.; Escribano-Romero, E.; Poderoso, T.; de Oya, N.J. Pathogenicity and virulence of West Nile virus revisited eight decades after its first isolation. Virulence 2021, 12, 1145–1173. [Google Scholar] [CrossRef] [PubMed]
- Chancey, C.; Grinev, A.; Volkova, E.; Rios, M. The global ecology and epidemiology of West Nile virus. Biomed. Res. Int. 2015, 2015, 376230. [Google Scholar] [CrossRef] [PubMed]
- Hernández-Triana, L.M.; Jeffries, C.L.; Mansfield, K.L.; Carnell, G.; Fooks, A.R.; Johnson, N. Emergence of west nile virus lineage 2 in europe: A review on the introduction and spread of a mosquito-borne disease. Front. Public Health 2014, 2, 271. [Google Scholar] [CrossRef]
- McMullen, A.R.; Albayrak, H.; May, F.J.; Davis, C.T.; Beasley, D.W.C.; Barrett, A.D.T. Molecular evolution of lineage 2 West Nile virus. J. Gen. Virol. 2013, 94, 318–325. [Google Scholar] [CrossRef]
- Campbell, G.L.; Marfin, A.A.; Lanciotti, R.S.; Gubler, D.J. West Nile virus. Lancet Infect. Dis. 2002, 9, 519–529. [Google Scholar] [CrossRef]
- Petersen, L.R.; Brault, A.C.; Nasci, R.S. West Nile virus: Review of the literature. JAMA 2013, 310, 308–315. [Google Scholar] [CrossRef]
- Pingen, M.; Schmid, M.A.; Harris, E.; McKimmie, C.S. Mosquito Biting Modulates Skin Response to Virus Infection. Trends Parasitol. 2017, 33, 645–657. [Google Scholar] [CrossRef]
- Cho, H.; Diamond, M.S. Immune responses to West Nile virus infection in the central nervous system. Viruses 2012, 4, 3812–3830. [Google Scholar] [CrossRef]
- Zou, S.; Foster, G.A.; Dodd, R.Y.; Petersen, L.R.; Stramer, S.L. West Nile fever characteristics among viremic persons identified through blood donor screening. J. Infect. Dis. 2010, 202, 1354–1361. [Google Scholar] [CrossRef] [PubMed]
- Hayes, E.B.; Sejvar, J.J.; Zaki, S.R.; Lanciotti, R.S.; Bode, A.V.; Campbell, G.L. Virology, Pathology, and Clinical Manifestations of West Nile Virus Disease. Emerg. Infect. Dis. 2005, 11, 1174–1179. [Google Scholar] [CrossRef] [PubMed]
- Suthar, M.S.; Diamond, M.S.; Gale, M., Jr. West Nile virus infection and immunity. Nat. Rev. Microbiol. 2013, 11, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Martín-Acebes, M.A.; Saiz, J.-C. West Nile virus: A re-emerging pathogen revisited. World J. Virol. 2012, 12, 51–70. [Google Scholar] [CrossRef] [PubMed]
- Kain, M.P.; Skinner, E.B.; Athni, T.S.; Ramirez, A.L.; Mordecai, E.A.; van den Hurk, A.F. Not all mosquitoes are created equal: A synthesis of vector competence experiments reinforces virus associations of Australian mosquitoes. PLoS Negl. Trop. Dis. 2022, 16, e0010768. [Google Scholar] [CrossRef] [PubMed]
- Vasić, A.; Oșlobanu, L.E.; Marinov, M.; Crivei, L.A.; Rățoi, I.A.; Aniță, A.; Aniță, D.; Doroșencu, A.; Alexe, V.; Răileanu, S.; et al. Evidence of West Nile Virus (WNV) Circulation in Wild Birds and WNV RNA Negativity in Mosquitoes of the Danube Delta Biosphere Reserve, Romania, 2016. Trop Med. Infect. Dis. 2019, 4, 116. [Google Scholar] [CrossRef]
- Jansen, S.; Heitmann, A.; Lühken, R.; Leggewie, M.; Helms, M.; Badusche, M.; Rossini, G.; Schmidt-Chanasit, J.; Tannich, E. Culex torrentium: A Potent Vector for the Transmission of West Nile Virus in Central Europe. Viruses 2019, 11, 492. [Google Scholar] [CrossRef]
- Ciota, A.T. West Nile virus and its vectors. Curr. Opin. Insect. Sci. 2017, 22, 28–36. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Kramer, L.D.; Jones, M.J.; Marra, P.P.; Daszak, P. West Nile virus epidemics in North America are driven by shifts in 810 mosquito feeding behavior. PLoS Biol. 2006, 4, e82. [Google Scholar] [CrossRef]
- Vogels, C.B.; Göertz, G.P.; Pijlman, G.P.; Koenraadt, C.J. Vector competence of European mosquitoes for West Nile virus. Emerg. Microbes Infect. 2017, 6, e96. [Google Scholar] [CrossRef]
- Bardos, V.; Adamcova, J.; Dedei, S.; Gjini, N.; Rosicky, B.; Simkova, A. Neutralizing antibodies against some neurotropic viruses determined in human sera in Albania. J. Hyg. Epidemiol. Microbiol. Immunol. 1959, 3, 277–282. [Google Scholar]
- Joubert, L.; Oudar, J.; Hannoun, C.; Beytout, D.; Corniou, B.; Guillon, J.C.; Panthier, R. Epidemiology of the West Nile virus: Study of a focus in Camargue. IV. Meningo-encephalomyelitis of the horse. Ann. Inst. Pasteur. 1970, 118, 239–247. [Google Scholar]
- Chaskopoulou, A.; Dovas, C.I.; Chaintoutis, S.C.; Bouzalas, I.; Ara, G.; Papanastassopoulou, M. Evidence of enzootic circulation of West Nile virus (Nea Santa-Greece-2010, lineage 2), Greece, May to July 2011. Eur. Surveill. 2011, 16, 19933. [Google Scholar] [CrossRef]
- Malkinson, M.; Banet, C. The role of birds in the ecology of West Nile virus in Europe and Africa. Curr. Top Microbiol. Immunol. 2002, 267, 309–322. [Google Scholar] [PubMed]
- Watts, M.J.; Monteys, V.S.I.; Mortyn, P.G.; Kotsila, P. The rise of West Nile Virus in Southern and Southeastern Europe: A spatial-temporal analysis investigating the combined effects of climate, land use and economic changes. One Health 2021, 13, 100315. [Google Scholar] [CrossRef] [PubMed]
- Beermann, S.; Dobler, G.; Faber, M.; Frank, C.; Habedank, B.; Hagedorn, P.; Kampen, H.; Kuhn, C.; Nygren, T.; Schmidt-Chanasit, J.; et al. Impact of climate change on vector- and rodent-borne infectious diseases. J. Health Monit. 2023, 8, 33–61. [Google Scholar]
- Rezza, G.; Nicoletti, L.; Angelini, R.; Romi, R.; Finarelli, A.C.; Panning, M. Infection with chikungunya virus in Italy: An outbreak in a temperate region. Lancet 2007, 370, 1840–1846. [Google Scholar] [CrossRef] [PubMed]
- Emmanouil, M.; Evangelidou, M.; Papa, A.; Mentis, A. Importation of dengue, Zika and chikungunya infections in Europe: The current situation in Greece. New Microbes New Infect. 2020, 35, 100663. [Google Scholar] [CrossRef]
- Cochet, A.; Calba, C.; Jourdain, F.; Grard, G.; Durand, G.A.; Guinard, A.; Investigation Team; Noël, H.; Paty, M.C.; Franke, F. Autochthonous dengue in mainland France, 2022: Geographical extension and incidence increase. Eur. Surveill. 2022, 27, 2200818. [Google Scholar] [CrossRef]
- European Center for Disease Control (ECDC), Stockholm, Sweden. Zika Virus Disease in Var Department, France. ECDC. 2019. Available online: https://www.ecdc.europa.eu/sites/default/files/documents/RRA-Zika-France-16-Oct-2019-corrected.pdf (accessed on 16 October 2022).
- Sauer, F.G.; Timmermann, E.; Lange, U.; Lühken, R.; Kiel, E. Effects of Hibernation Site, Temperature, and Humidity on the Abundance and Survival of Overwintering Culex pipiens pipiens and Anopheles messeae (Diptera: Culicidae). J. Med. Entomol. 2022, 59, 2013–2021. [Google Scholar] [CrossRef]
- Reisen, W.K.; Wheeler, S.S. Overwintering of West Nile Virus in the United States Review. J. Med. Entomol. 2019, 56, 1498–1507. [Google Scholar] [CrossRef] [PubMed]
- Marini, G.; Manica, M.; Delucchi, L.; Pugliese, A.; Rosà, R. Spring temperature shapes West Nile virus transmission in Europe. Acta Trop. 2021, 215, 105796. [Google Scholar] [CrossRef] [PubMed]
- Marcantonio, M.; Rizzoli, A.; Metz, M.; Rosà, R.; Marini, G.; Chadwick, E.; Neteler, M. Identifying the Environmental Conditions Favouring West Nile Virus Outbreaks in Europe. PLoS ONE 2015, 10, e0121158. [Google Scholar] [CrossRef] [PubMed]
- Ciota, A.T.; Keyel, A.C. The Role of Temperature in Transmission of Zoonotic Arboviruses. Viruses 2019, 11, 1013. [Google Scholar] [CrossRef]
- Farooq, Z.; Sjödin, H.; Semenza, J.C.; Tozan, Y.; Sewe, M.O.; Wallin, J.; Rocklöva, J. European projections of West Nile virus transmission under climate change scenarios. One Health 2023, 16, 100509. [Google Scholar] [CrossRef]
- Liu, Z.; Zhang, Z.; Lai, Z.; Zhou, T.; Jia, Z.; Gu, J.; Wu, K.; Chen, X.G. Temperature Increase Enhances Aedes albopictus Competence to Transmit Dengue Virus. Front. Microbiol. 2017, 8, 2337. [Google Scholar] [CrossRef]
- Reiter, P. Climate change and mosquito-borne disease. Rev. Environ. Health Perspect. 2001, 109, 141–161. [Google Scholar]
- Brown, J.J.; Pascual, M.; Wimberly, M.C.; Johnson, L.R.; Murdock, C.C. Humidity—The overlooked variable in the thermal biology of mosquito-borne disease. Ecol. Lett. 2023, 26, 1029–1049. [Google Scholar] [CrossRef]
- Cai, X.; Zhao, J.; Deng, H.; Xiao, J.; Liu, T.; Zeng, W.; Li, X.; Hu, J.; Huang, C.; Zhu, G.; et al. Effects of temperature, relative humidity, and illumination on the entomological parameters of Aedes albopictus: An experimental study. Int. J. Biometeorol. 2023, 67, 687–694. [Google Scholar] [CrossRef]
- Lindsay, S.W.; Wilkins, H.A.; Zieler, H.A.; Daly, R.J.; Petrarca, V.; Byass, P. Ability of Anopheles gambiae mosquitoes to transmit malaria during the dry and wet seasons in an area of irrigated rice cultivation in The Gambia. J. Trop. Med. Hyg. 1991, 94, 313–324. [Google Scholar]
- Chow, W.K.; Beebe, N.W.; Ambrose, L.; Pickering, P.; Cooper, R.D. Seasonal assessment on the effects of time of night, temperature and humidity on the biting profile of Anopheles farauti in north Queensland, Australia using a population naive to malaria vector control pressures. Malar. J. 2023, 22, 85. [Google Scholar] [CrossRef] [PubMed]
- Keyel, A.C.; Timm, O.E.; Backenson, P.B.; Prussing, C.; Quinones, S.; McDonough, K.A.; Vuille, M.; Conn, J.E.; Armstrong, P.M.; Andreadis, T.G.; et al. Seasonal temperatures and hydrological conditions improve the prediction of West Nile virus infection rates in Culex mosquitoes and human case counts in New York and Connecticut. PLoS ONE 2019, 14, e0217854. [Google Scholar] [CrossRef] [PubMed]
- Gorris, M.E.; Randerson, J.T.; Coffield, S.R.; Treseder, K.K.; Zender, C.S.; Xu, C.; Manore, C.A. Assessing the Influence of Climate on the Spatial Pattern of West Nile Virus Incidence in the United States. Environ. Health Perspect. 2023, 131, 047016. [Google Scholar] [CrossRef]
- Tran, A.; Sudre, S.; Paz, S.; Rossi, M.; Desbrosse, A.; Chevalier, V.; Semenza, J.C. Environmental predictors of West Nile fever risk in Europe. Int. J. Health Geogr. 2014, 13, 26. [Google Scholar] [CrossRef] [PubMed]
- Cotar, A.I.; Falcuta, E.; Prioteasa, L.F.; Dinu, S.; Ceianu, C.S.; Paz, S. Transmission Dynamics of the West Nile Virus in Mosquito Vector Populations under the Influence of Weather Factors in the Danube Delta, Romania. EcoHealth 2016, 13, 796–807. [Google Scholar] [CrossRef]
- Paz, S.; Albersheim, I. Influence of warming tendency on Culex Pipiens population abundance and on the probability of West Nile Fever outbreaks (Israeli Case Study: 2001–2005). EcoHealth 2008, 5, 40–48. [Google Scholar] [CrossRef]
- Korytár, L.; Peňazziová, K.; Pistl, J.; Tichá, E.; Čabanová, V.; Csank, T. Retrospective review and current knowledge on the occurrence of West Nile virus in mosquito vectors, reservoirs and hosts in Slovakia (Central Europe). Acta Virol. 2020, 64, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Benelli, G.; Wilke, A.B.B.; Beier, J.C. Aedes albopictus (Asian tiger mosquito). Trends Parasitol. 2020, 36, 942–943. [Google Scholar] [CrossRef]
- Oliveira, S.; Rocha, J.; Sousa, C.A.; Capinha, C. Wide and increasing suitability for Aedes albopictus in Europe is congruent across distribution models. Sci. Rep. 2021, 11, 9916. [Google Scholar] [CrossRef]
- European Center for Disease Control (ECDC), Stockholm, Sweden. Aedes Albopictus—Factsheet for Experts, 20 December 2016. Available online: https://www.ecdc.europa.eu/en/disease-vectors/facts/mosquito-factsheets/aedes-albopictus (accessed on 20 August 2023).
- Laporta, G.Z.; Potter, A.M.; Oliveira, J.F.A.; Bourke, B.P.; Pecor, D.B.; Linton, Y.-M. Global Distribution of Aedes aegypti and Aedes albopictus in a Climate Change Scenario of Regional Rivalry. Insects 2023, 14, 49. [Google Scholar] [CrossRef]
- Montecino-Latorre, D.; Barker, C.M. Overwintering of West Nile virus in a bird community with a communal crow roost. Sci. Rep. 2018, 8, 6088. [Google Scholar] [CrossRef] [PubMed]
- Kampen, H.; Tews, B.A.; Werner, D. First Evidence of West Nile Virus Overwintering in Mosquitoes in Germany. Viruses 2021, 13, 2463. [Google Scholar] [CrossRef] [PubMed]
- Kjellstrom, T.; Friel, S.; Dixon, J.; Corvalan, C.; Rehfuess, E.; Campbell-Lendrum, D.; Gore, F.; Bartram, J. Urban environmental health hazards and health equity. J. Urban Health 2007, 84, 86–97. [Google Scholar] [CrossRef]
- Kondilis, E.; Giannakopoulos, S.; Gavana, M.; Ierodiakonou, I.; Waitzkin, H.; Benos, A. Economic crisis, restrictive policies, and the population’s health and health care: The Greek case. Am. J. Public Health 2013, 103, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Paz, S.; Malkinson, D.; Green, M.S.; Tsioni, G.; Papa, A.; Danis, K.; Sirbu, A.; Ceianu, C.; Katalin, K.; Ferenczi, E.; et al. Permissive summer temperatures of the 2010 European West Nile fever upsurge. PLoS ONE 2013, 8, e56398. [Google Scholar] [CrossRef]
- Gratz, N.G. Critical review of the vector status of Aedes albopictus. Med. Vet. Entomol. 2004, 18, 215–227. [Google Scholar] [CrossRef]
- Giunti, G.; Becker, N.; Benelli, G. Invasive mosquito vectors in Europe: From bioecology to surveillance and management. Acta Trop. 2023, 239, 106832. [Google Scholar] [CrossRef]
- Miranda, M.M.; Barceló, C.; Arnoldi, D.; Augsten, X.; Bakran-Lebl, K.; Balatsos, G.; Bengoa, M.; Bindler, P.; Boršová, K.; Bourquia, M.; et al. Consortium AIM-COST/AIM-Surv AIMSurv: First pan-European harmonized surveillance of Aedes invasive mosquito species of relevance for human vector-borne diseases. GigaByte 2022, 2022, gigabyte57. [Google Scholar]
- Wint, W.; Jones, P.; Kraemer, M.; Alexander, N.; Schaffnerd, F. Past, present and future distribution of the yellow fever mosquito Aedes aegypti: The European paradox. Sci. Total Environ. 2022, 847, 157566. [Google Scholar] [CrossRef]
- Trájer, A.J. Aedes aegypti in the Mediterranean container ports at the time of climate change: A time bomb on the mosquito vector map of Europe. Heliyon 2021, 7, e07981. [Google Scholar] [CrossRef]
- Eritja, R.; Escosa, R.; Lucientes, J.; Marques, E.; Roiz, D.; Ruiz, S. Worldwide invasion of vector mosquitoes: Present European distribution and challenges in Spain. Biol Invasions 2005, 7, 87–97. [Google Scholar] [CrossRef]
- Mavridis, K.; Fotakis, E.A.; Kioulos, I.; Mpellou, S.; Konstantas, S.; Varela, E.; Gewehr, S.; Diamantopoulos, V.; Vontas, J. Detection of West Nile virus—Lineage 2 in Culex pipiens mosquitoes, associated with disease outbreak in Greece, 2017. Acta Trop. 2018, 182, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Patsoula, E.; Vakali, A.; Balatsos, G.; Pervanidou, D.; Beleri, S.; Tegos, N.; Baka, A.; Spanakos, G.; Georgakopoulou, T.; Tserkezou, P.; et al. West Nile virus circulation in mosquitoes in Greece (2010–2013). Biomed. Res. Int. 2016, 2016, 2450682. [Google Scholar] [CrossRef] [PubMed]
- Giesen, C.; Herrador, Z.; Fernandez-Martinez, B.; Figuerola, J.; Gangoso, L.; Vazquez, A.; Gómez-Barroso, D. A systematic review of environmental factors related to WNV circulation in European and Mediterranean countries. One Health 2023, 16, 100478. [Google Scholar] [CrossRef] [PubMed]
- Cuthbert, R.N.; Darriet, F.; Chabrerie, O.; Lenoir, J.; Courchamp, F.; Claeys, C.; Robert, V.; Jourdain, F.; Ulmer, R.; Diagne, C.; et al. Invasive hematophagous arthropods and associated diseases in a changing world. Parasit. Vectors 2023, 16, 291. [Google Scholar] [CrossRef]
- Harrigan, R.J.; Thomassen, H.A.; Buermann, W.; Cummings, R.F.; Kahn, M.E.; Smith, T.B. Economic conditions predict prevalence of West Nile virus. PLoS ONE 2010, 5, e15437. [Google Scholar] [CrossRef]
- Kouamé, R.M.A.; Guglielmo, F.; Abo, K.; Ouattara, A.F.; Chabi, J.; Sedda, L.; Donnelly, M.J.; Edi, C. Education and So-cio-economic status are key factors influencing use of insecticides and malaria knowledge in rural farmers in Southern Côte d’Ivoire. BMC Public Health 2022, 22, 2443. [Google Scholar] [CrossRef]
- Savage, H.M.; Anderson, M.; Gordon, E.; McMillen, L.; Colton, L.; Delorey, M.; Sutherland, G.; Aspen, S.; Charnetzky, D.; Burkhalter, K. Host-seeking heights, host-seeking activity patterns, and West Nile virus infection rates for members of the Culex pipiens complex at different habitat types within the hybrid zone, Shelby County, TN, 2002 (Diptera: Culicidae). J Med. Entomol. 2008, 45, 276–288. [Google Scholar] [CrossRef]
- Bonovas, S.; Nikolopoulos, G. High-burden epidemics in Greece in the era of economic crisis. Early signs of a public health tragedy. J. Prev. Med. Hyg. 2012, 3, 169–171. [Google Scholar]
- Hubalek, Z. European Experience with the West Nile Virus Ecology and Epidemiology: Could It Be Relevant for the New World? Viral Immunol. 2000, 13, 415–426. [Google Scholar] [CrossRef]
- Antoniadis, A.; Alexiou-Daniel, S.; Malissiovas, N.; Doutsos, J.; Polyzoni, T.; LeDuc, J.W.; Peters, C.J.; Saviolakis, G. Seroepidemiological survey for antibodies to arboviruses in Greece. In Archives of Virology Supplementum; Springer: Vienna, Austria, 1990; Suppl. 1; pp. 277–285. [Google Scholar]
- Valiakos, G.; Touloudi, A.; Athanasiou, L.V.; Giannakopoulos, A.; Iacovakis, C.; Birtsas, P.; Spyrou, V.; Dalabiras, Z.; Petrovska, L.; Billinis, C. Serological and molecular investigation into the role of wild birds in the epidemiology of West Nile virus in Greece. Virol. J. 2012, 9, 266. [Google Scholar] [CrossRef] [PubMed]
- Papa, A.; Perperidou, P.; Tzouli, A.; Castilletti, C. West Nile virus—Neutralizing antibodies in humans in Greece. Vector Borne Zoonotic Dis. 2010, 10, 655–658. [Google Scholar] [CrossRef] [PubMed]
- Danis, K.; Papa, A.; Theocharopoulos, G.; Dougas, G.; Athanasiou, M.; Detsis, M.; Baka, A.; Lytras, T.; Mellou, K.; Bonovas, S.; et al. Outbreak of West Nile virus infection in Greece, 2010. Emerg. Infect. Dis. 2011, 10, 1868–1872. [Google Scholar]
- Available online: https://eody.gov.gr/en/epidemiological-statistical-data/annual-epidemiological-data/ (accessed on 23 August 2023).
- Ladbury, G.A.F.; Gavana, M.; Danis, K.; Papa, A.; Papamichail, D.; Mourelatos, S.; Gewehr, S.; Theocharopoulos, G.; Bonovas, S.; Benos, A.; et al. Population seroprevalence study after a West Nile virus lineage 2 epidemic, Greece, 2010. PLoS ONE 2013, 8, e80432. [Google Scholar] [CrossRef] [PubMed]
- Pervanidou, D.; Kefaloudi, C.N.; Vakali, A.; Tsakalidou, O.; Karatheodorou, M.; Tsioka, K.; Evangelidou, M.; Mellou, K.; Pappa, S.; Stoikou, K.; et al. The 2022 West Nile Virus Season in Greece; A Quite Intense Season. Viruses 2023, 15, 1481. [Google Scholar] [CrossRef] [PubMed]
- Gossner, C.M.; Marrama, L.; Carson, M.; Allerberger, F.; Calistri, P.; Dilaveris, D.; Lecollinet, S.; Morgan, D.; Nowotny, N.; Paty, M.-C.; et al. West Nile virus surveillance in Europe: Moving towards an integrated animal-human-vector approach. Eur. Surveill. 2017, 22, 30526. [Google Scholar] [CrossRef]
- Bakonyi, T.; Ivanics, E.; Erdelyi, K.; Ursu, K.; Ferenczi, E.; Weissenböck, H.; Nowotny, R. Lineage 1 and 2 strains of encephalitic West Nile virus, central Europe. Emerg. Infect. Dis. 2006, 12, 618–623. [Google Scholar] [CrossRef]
- Papa, A.; Papadopoulou, E.; Chatzixanthouliou, C.; Glouftsios, P.; Pappa, S.; Pervanidou, D.; Georgiou, L. Emergence of West Nile Virus Lineage 2 Belonging to the Eastern European Subclade, Greece. Arch. Virol. 2019, 164, 1673–1675. [Google Scholar] [CrossRef]
- Magombedze, G.; Ferguson, N.M.; Ghani, A.C. A trade-off between dry season survival longevity and wet season high net reproduction can explain the persistence of anopheles mosquitoes. Parasites Vectors 2018, 11, 576. [Google Scholar] [CrossRef]
- Chowdhury, F.R.; Ibrahim, Q.S.U.; Bari, M.S.; Alam, M.M.J.; Dunachie, S.J.; Rodriguez-Morales, A.J.; Patwary, M.I. The associ-ation between temperature, rainfall and humidity with common climate-sensitive infectious diseases in Bangladesh. PLoS ONE 2018, 13, e0199579. [Google Scholar] [CrossRef]
- Klingelhöfer, D.; Braun, M.; Kramer, I.M.; Reuss, F.; Müller, R.; Groneberg, D.A.; Brüggmann, D. A virus becomes a global concern: Research activities on West-Nile virus. Emerg. Microbes Infect. 2023, 12, 2256424. [Google Scholar] [CrossRef] [PubMed]
- Fagre, A.C.; Lyons, S.; Staples, J.E.; Lindsey, N. West Nile Virus and Other Nationally Notifiable Arboviral Diseases—United States, 2021. MMWR Morb. Mortal Wkly. Rep. 2023, 72, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Papa, A. Emerging arboviral human diseases in Southern Europe. J. Med. Virol. 2017, 89, 1315–1322. [Google Scholar] [CrossRef]
- European Food Safety Authority. European Centre for Disease Prevention and Control the European Union One Health 2021 Zoonoses Report. EFSA J. 2022, 20, e07666. [Google Scholar]
- Rocheleau, J.-P.; Kotchi, S.-O.; Arsenault, J. Can local risk of West Nile virus infection be predicted from previous cases? A descriptive study in Quebec, 2011–2016. Can. J. Public Health 2020, 111, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Casades-Martí, L.; Cuadrado-Matías, R.; Peralbo-Moreno, A.; Baz-Flores, S.; Fierro, Y.; Ruiz-Fonsac, F. Insights into the spatiotemporal dynamics of West Nile virus transmission in emerging scenarios. One Health 2023, 16, 100557. [Google Scholar] [CrossRef]
- Tsioka, K.; Gewehr, S.; Kalaitzopoulou, S.; Pappa, S.; Stoikou, K.; Mourelatos, S.; Papa, A. Detection and molecular characterization of West Nile virus in Culex pipiens mosquitoes in Central Macedonia, Greece, 2019–2021. Acta Trop. 2022, 230, 106391. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Environmental Factors | References | |||
|---|---|---|---|---|
| Mean Temperature | Winter | Spring | Summer | |
| Type of association | negative | positive | positive | [31,32,33,34,37,44] |
| Proposed mechanism | affects vector survival, depending on the vector | increases biting rate, increases host-to-vector transmission probability, accelerates mosquito viral incubation period, potentially enhances oviposition activity | affects distribution, behaviour and survival of the mosquito vector, reduces incubation period in the competent vectors and affects host distribution and behaviour very high temperatures reduce mosquito survival | |
| Relative Humidity/Precipitation | Winter | Spring | Summer | |
| Type of association | negative | positive/negative | negative | [34,36,45,46,47] |
| Proposed mechanism | affects vector survival | increases the standing water resources at the beginning of the hot season/consequence of favourable higher temperatures- drought creates stagnant water | drought leads to increases in mosquito numbers and increases biting rate | |
| Ground Moisture/Vegetation | Winter | Spring | Summer | |
| Type of association | positive (when followed by a decrease in spring/summer) | depending on mean temperatures | positive | [43,46] |
| Proposed mechanism | variations influence the presence and size of the WNV vector populations | inconclusive | favours vector abundance | [48] |
| Vector-Associated Factors | Current Situation in Europe | Effect on Potential Outbreak | ||
| Installation/availability of potent arthropod vectors | installation and endemicity in the Mediterranean Basin/Europe of Aedes mosquitoes, mainly albopictus | positive (pros: very potent vectors, are able to infestate urbanized areas, bite throughout the daytime, contras: preferentially bite humans) | [49,50,51,52] | |
| Hosts/reservoirs | climate change favours vector abundance, geographic location on the route of migratory birds from Africa | positive | [53] | |
| Vector and viral overwintering | evidence of enzootic circulation, viral overwintering in non-migratory birds, evidence of viral overwintering in mosquitoes | positive | [23,24,54,55] | |
| Socioeconomic Factors | Current Situation in Europe | Effect on Potential Outbreak | ||
| Economic crisis | coincided with the outbreak in 2010 | negative correlation of the Gross Domestic Product with WNV infections | [56] | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kouroupis, D.; Charisi, K.; Pyrpasopoulou, A. The Ongoing Epidemic of West Nile Virus in Greece: The Contribution of Biological Vectors and Reservoirs and the Importance of Climate and Socioeconomic Factors Revisited. Trop. Med. Infect. Dis. 2023, 8, 453. https://doi.org/10.3390/tropicalmed8090453
Kouroupis D, Charisi K, Pyrpasopoulou A. The Ongoing Epidemic of West Nile Virus in Greece: The Contribution of Biological Vectors and Reservoirs and the Importance of Climate and Socioeconomic Factors Revisited. Tropical Medicine and Infectious Disease. 2023; 8(9):453. https://doi.org/10.3390/tropicalmed8090453
Chicago/Turabian StyleKouroupis, Dimitrios, Konstantina Charisi, and Athina Pyrpasopoulou. 2023. "The Ongoing Epidemic of West Nile Virus in Greece: The Contribution of Biological Vectors and Reservoirs and the Importance of Climate and Socioeconomic Factors Revisited" Tropical Medicine and Infectious Disease 8, no. 9: 453. https://doi.org/10.3390/tropicalmed8090453