

Detection of Anti-ZIKV NS1 IgA, IgM, and Combined IgA/IgM and Identification of IL-4 and IL-10 as Potential Biomarkers for Early ZIKV and DENV Infections in Hyperendemic Regions, Thailand
 , , , , ,
, , , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement, Collection of Human Specimens, and Clinical Data
2.2. Sample Classification and Study Design
2.3. RNA Extraction and Real-Time RT-PCR for ZIKV and DENV1-4 Diagnosis
2.4. ZIKV and DENV Serological Diagnosis
2.5. DENV1-4 NS1 IgG In-House ELISA
2.6. Anti-ZIKV NS1 IgA/M, IgM, and IgA ELISAs
2.7. Quantification of Cytokines Using Multiplex Immunoassay
2.8. Statistical Analysis
3. Results
3.1. Data of Participants and Samples
3.2. Detection of ZIKV IgA/M, IgM, and IgA in Confirmed ZIKV Cases in Thailand
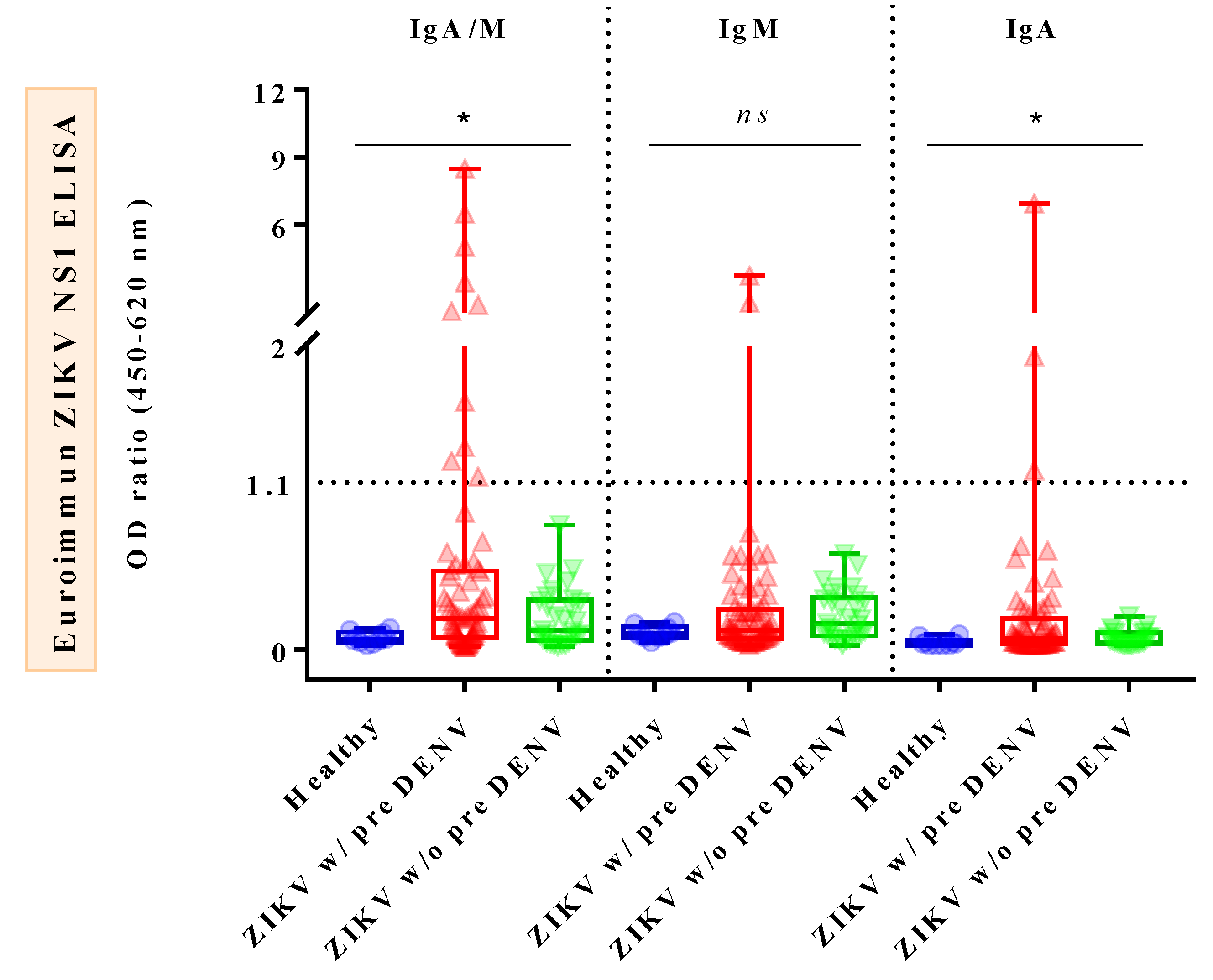
3.3. Comparison of Combined IgA/M, IgM, and IgA Reactivities against ZIKV NS1 ELISAs in Confirmed ZIKV Cases, Confirmed DENV Cases, and Healthy Donors
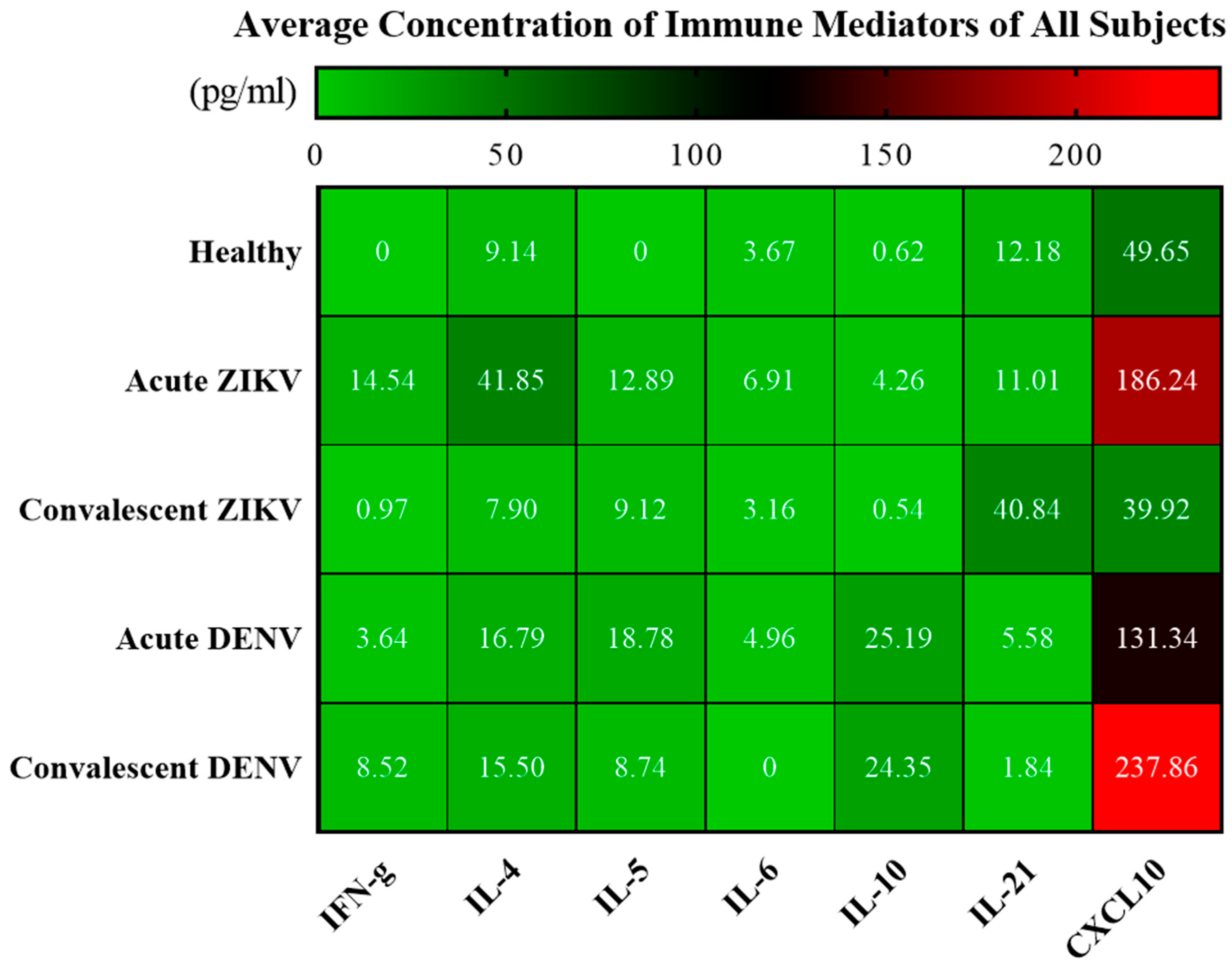
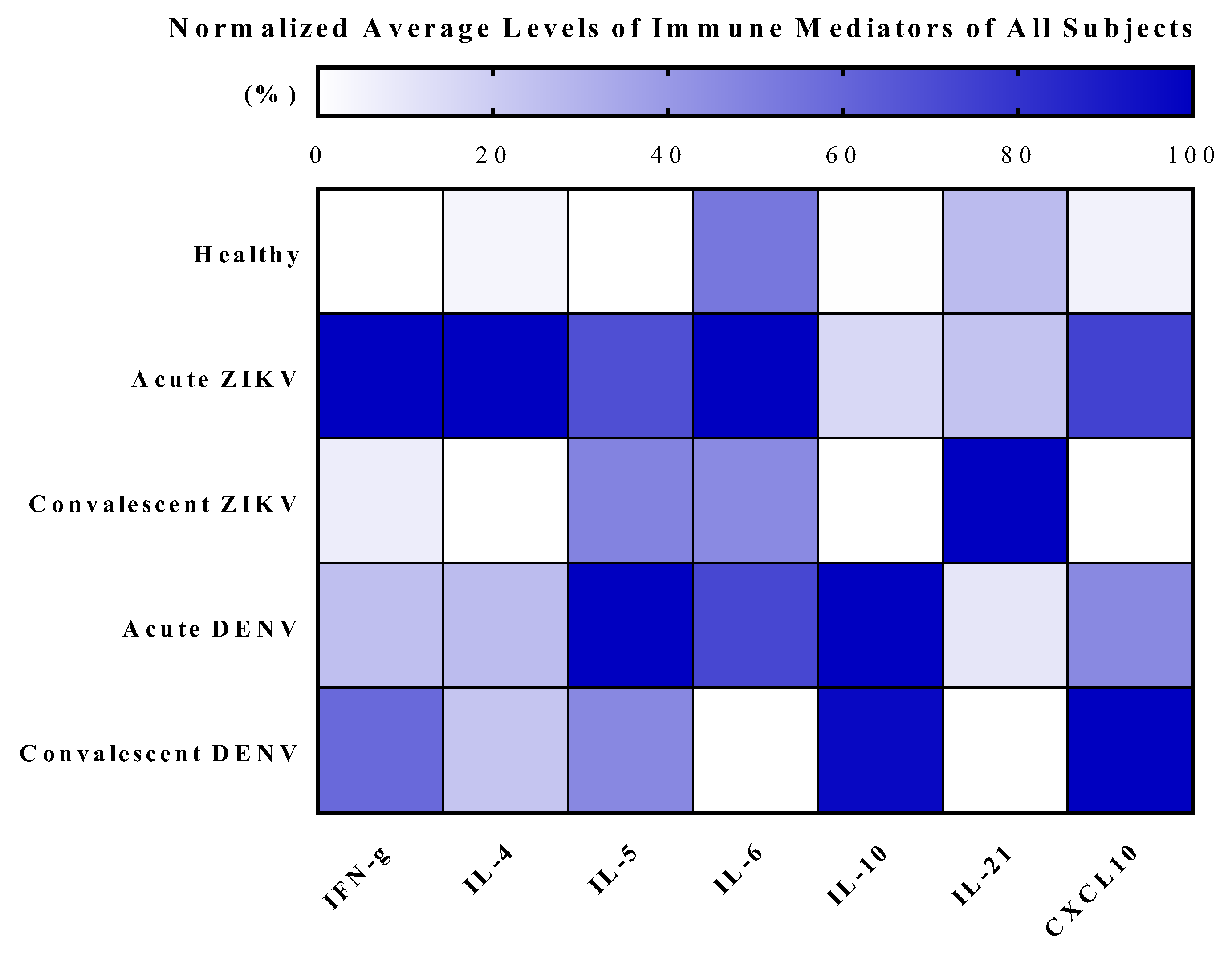
3.4. Cytokine Analysis of Zika and Dengue Patients in Endemic Country, Thailand
3.5. Association between Cytokines and Symptoms in Acute ZIKV-Infected Patients
3.6. Correlations among Cytokine Levels in Acute ZIKV-Infected Patients
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Heinz, F.X.; Stiasny, K. The Antigenic Structure of Zika Virus and Its Relation to Other Flaviviruses: Implications for Infection and Immunoprophylaxis. Microbiol. Mol. Biol. Rev. 2017, 81, e00055-16. [Google Scholar] [CrossRef] [PubMed]
- Shuaib, W.; Stanazai, H.; Abazid, A.G.; Mattar, A.A. Re-Emergence of Zika Virus: A Review on Pathogenesis, Clinical Manifestations, Diagnosis, Treatment, and Prevention. Am. J. Med. 2016, 129, e877–e879. [Google Scholar] [CrossRef] [PubMed]
- Relich, R.F.; Loeffelholz, M. Zika Virus. Clin. Lab. Med. 2017, 37, 253–267. [Google Scholar] [CrossRef] [PubMed]
- Hills, S.L.; Fischer, M.; Petersen, L.R. Epidemiology of Zika Virus Infection. J. Infect. Dis. 2017, 216, S868–S874. [Google Scholar] [CrossRef]
- Judice, C.C.; Tan, J.J.L.; Parise, P.L.; Kam, Y.W.; Milanez, G.P.; Leite, J.A.; Caserta, L.C.; Arns, C.W.; Resende, M.R.; Angerami, R.; et al. Efficient detection of Zika virus RNA in patients’ blood from the 2016 outbreak in Campinas, Brazil. Sci. Rep. 2018, 8, 4012. [Google Scholar] [CrossRef]
- L’Huillier, A.G.; Lombos, E.; Tang, E.; Perusini, S.; Eshaghi, A.; Nagra, S.; Frantz, C.; Olsha, R.; Kristjanson, E.; Dimitrova, K.; et al. Evaluation of Altona Diagnostics RealStar Zika Virus Reverse Transcription-PCR Test Kit for Zika Virus PCR Testing. J. Clin. Microbiol. 2017, 55, 1576–1584. [Google Scholar] [CrossRef]
- Munoz-Jordan, J.L. Diagnosis of Zika Virus Infections: Challenges and Opportunities. J. Infect. Dis. 2017, 216, S951–S956. [Google Scholar] [CrossRef]
- Cabral-Castro, M.J.; Cavalcanti, M.G.; Peralta, R.H.S.; Peralta, J.M. Molecular and serological techniques to detect co-circulation of DENV, ZIKV and CHIKV in suspected dengue-like syndrome patients. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2016, 82, 108–111. [Google Scholar] [CrossRef]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and serologic properties of Zika virus associated with an epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232–1239. [Google Scholar] [CrossRef]
- Tyson, J.; Tsai, W.Y.; Tsai, J.J.; Massgard, L.; Stramer, S.L.; Lehrer, A.T.; Nerurkar, V.R.; Wang, W.K. A high-throughput and multiplex microsphere immunoassay based on non-structural protein 1 can discriminate three flavivirus infections. PLoS Negl. Trop. Dis. 2019, 13, e0007649. [Google Scholar] [CrossRef]
- Wong, S.J.; Furuya, A.; Zou, J.; Xie, X.; Dupuis, A.P., 2nd; Kramer, L.D.; Shi, P.Y. A Multiplex Microsphere Immunoassay for Zika Virus Diagnosis. EBioMedicine 2017, 16, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Khalifian, S.; Raimondi, G.; Brandacher, G. The use of luminex assays to measure cytokines. J. Investig. Dermatol. 2015, 135, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Tyson, J.; Tsai, W.Y.; Tsai, J.J.; Brites, C.; Mässgård, L.; Ha Youn, H.; Pedroso, C.; Drexler, J.F.; Stramer, S.L.; Balmaseda, A.; et al. Combination of Nonstructural Protein 1-Based Enzyme-Linked Immunosorbent Assays Can Detect and Distinguish Various Dengue Virus and Zika Virus Infections. J. Clin. Microbiol. 2019, 57, e01464-18. [Google Scholar] [CrossRef] [PubMed]
- Steinhagen, K.; Probst, C.; Radzimski, C.; Schmidt-Chanasit, J.; Emmerich, P.; van Esbroeck, M.; Schinkel, J.; Grobusch, M.P.; Goorhuis, A.; Warnecke, J.M.; et al. Serodiagnosis of Zika virus (ZIKV) infections by a novel NS1-based ELISA devoid of cross-reactivity with dengue virus antibodies: A multicohort study of assay performance, 2015 to 2016. Euro Surveill. 2016, 21, 30426. [Google Scholar] [CrossRef]
- Balmaseda, A.; Guzmán, M.G.; Hammond, S.; Robleto, G.; Flores, C.; Téllez, Y.; Videa, E.; Saborio, S.; Pérez, L.; Sandoval, E.; et al. Diagnosis of dengue virus infection by detection of specific immunoglobulin M (IgM) and IgA antibodies in serum and saliva. Clin. Vaccine Immunol. 2003, 10, 317–322. [Google Scholar] [CrossRef]
- Warnecke, J.M.; Lattwein, E.; Saschenbrecker, S.; Stöcker, W.; Schlumberger, W.; Steinhagen, K. Added value of IgA antibodies against Zika virus non-structural protein 1 in the diagnosis of acute Zika virus infections. J. Virol. Methods 2019, 267, 8–15. [Google Scholar] [CrossRef]
- Morales, I.; Rosenberger, K.D.; Magalhaes, T.; Morais, C.N.L.; Braga, C.; Marques, E.T.A.; Calvet, G.A.; Damasceno, L.; Brasil, P.; Bispo de Filippis, A.M.; et al. Diagnostic performance of anti-Zika virus IgM, IgAM and IgG ELISAs during co-circulation of Zika, dengue, and chikungunya viruses in Brazil and Venezuela. PLoS Negl. Trop. Dis. 2021, 15, e0009336. [Google Scholar] [CrossRef]
- Hamel, R.; Dejarnac, O.; Wichit, S.; Ekchariyawat, P.; Neyret, A.; Luplertlop, N.; Perera-Lecoin, M.; Surasombatpattana, P.; Talignani, L.; Thomas, F.; et al. Biology of Zika Virus Infection in Human Skin Cells. J. Virol. 2015, 89, 8880–8896. [Google Scholar] [CrossRef]
- Mlakar, J.; Korva, M.; Tul, N.; Popovic, M.; Poljsak-Prijatelj, M.; Mraz, J.; Kolenc, M.; Resman Rus, K.; Vesnaver Vipotnik, T.; Fabjan Vodusek, V.; et al. Zika Virus Associated with Microcephaly. N. Engl. J. Med. 2016, 374, 951–958. [Google Scholar] [CrossRef]
- Medin, C.L.; Rothman, A.L. Zika Virus: The Agent and Its Biology, With Relevance to Pathology. Arch. Pathol. Lab. Med. 2017, 141, 33–42. [Google Scholar] [CrossRef]
- Noronha, L.; Zanluca, C.; Azevedo, M.L.; Luz, K.G.; Santos, C.N. Zika virus damages the human placental barrier and presents marked fetal neurotropism. Mem. Inst. Oswaldo Cruz 2016, 111, 287–293. [Google Scholar] [CrossRef]
- Kam, Y.W.; Leite, J.A.; Lum, F.M.; Tan, J.J.L.; Lee, B.; Judice, C.C.; Teixeira, D.A.T.; Andreata-Santos, R.; Vinolo, M.A.; Angerami, R.; et al. Specific Biomarkers Associated With Neurological Complications and Congenital Central Nervous System Abnormalities From Zika Virus-Infected Patients in Brazil. J. Infect. Dis. 2017, 216, 172–181. [Google Scholar] [CrossRef]
- Patro, A.R.K.; Mohanty, S.; Prusty, B.K.; Singh, D.K.; Gaikwad, S.; Saswat, T.; Chattopadhyay, S.; Das, B.K.; Tripathy, R.; Ravindran, B. Cytokine Signature Associated with Disease Severity in Dengue. Viruses 2019, 11, 34. [Google Scholar] [CrossRef]
- Puc, I.; Ho, T.C.; Yen, K.L.; Vats, A.; Tsai, J.J.; Chen, P.L.; Chien, Y.W.; Lo, Y.C.; Perng, G.C. Cytokine Signature of Dengue Patients at Different Severity of the Disease. Int. J. Mol. Sci. 2021, 22, 2879. [Google Scholar] [CrossRef] [PubMed]
- Winter, P.M.; Dung, N.M.; Loan, H.T.; Kneen, R.; Wills, B.; Thu le, T.; House, D.; White, N.J.; Farrar, J.J.; Hart, C.A.; et al. Proinflammatory cytokines and chemokines in humans with Japanese encephalitis. J. Infect. Dis. 2004, 190, 1618–1626. [Google Scholar] [CrossRef]
- Kalita, J.; Srivastava, R.; Mishra, M.K.; Basu, A.; Misra, U.K. Cytokines and chemokines in viral encephalitis: A clinicoradiological correlation. Neurosci. Lett. 2010, 473, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Abhishek, K.S.; Chakravarti, A.; Baveja, C.P.; Kumar, N.; Siddiqui, O.; Kumar, S. Association of interleukin-2, -4 and -10 with dengue severity. Indian J. Pathol. Microbiol. 2017, 60, 66–69. [Google Scholar] [CrossRef]
- Gowri Sankar, S.; Alwin Prem Anand, A. Cytokine IP-10 and GM-CSF are prognostic biomarkers for severity in secondary dengue infection. Hum. Immunol. 2021, 82, 438–445. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.H.; Leong, W.Y.; Wilder-Smith, A. Markers of dengue severity: A systematic review of cytokines and chemokines. J. Gen. Virol. 2016, 97, 3103–3119. [Google Scholar] [CrossRef]
- Tsai, J.J.; Liu, W.L.; Lin, P.C.; Huang, B.Y.; Tsai, C.Y.; Chou, P.H.; Lee, F.C.; Ping, C.F.; Lee, P.A.; Liu, L.T.; et al. An RT-PCR panel for rapid serotyping of dengue virus serotypes 1 to 4 in human serum and mosquito on a field-deployable PCR system. PLoS ONE 2019, 14, e0214328. [Google Scholar] [CrossRef]
- Sriburin, P.; Sittikul, P.; Kosoltanapiwat, N.; Sirinam, S.; Arunsodsai, W.; Sirivichayakul, C.; Limkittikul, K.; Chatchen, S. Incidence of Zika Virus Infection from a Dengue Epidemiological Study of Children in Ratchaburi Province, Thailand. Viruses 2021, 13, 1802. [Google Scholar] [CrossRef] [PubMed]
- Al-Qahtani, A.A.; Nazir, N.; Al-Anazi, M.R.; Rubino, S.; Al-Ahdal, M.N. Zika virus: A new pandemic threat. J. Infect. Dev. Ctries. 2016, 10, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Priyamvada, L.; Quicke, K.M.; Hudson, W.H.; Onlamoon, N.; Sewatanon, J.; Edupuganti, S.; Pattanapanyasat, K.; Chokephaibulkit, K.; Mulligan, M.J.; Wilson, P.C.; et al. Human antibody responses after dengue virus infection are highly cross-reactive to Zika virus. Proc. Natl. Acad. Sci. USA 2016, 113, 7852–7857. [Google Scholar] [CrossRef]
- Saiz, J.C.; Vazquez-Calvo, A.; Blazquez, A.B.; Merino-Ramos, T.; Escribano-Romero, E.; Martin-Acebes, M.A. Zika Virus: The Latest Newcomer. Front. Microbiol. 2016, 7, 496. [Google Scholar] [CrossRef]
- Hamer, D.H.; Wilson, M.E.; Jean, J.; Chen, L.H. Epidemiology, Prevention, and Potential Future Treatments of Sexually Transmitted Zika Virus Infection. Curr. Infect. Dis. Rep. 2017, 19, 16. [Google Scholar] [CrossRef]
- Kikuti, M.; Tauro, L.B.; Moreira, P.S.S.; Campos, G.S.; Paploski, I.A.D.; Weaver, S.C.; Reis, M.G.; Kitron, U.; Ribeiro, G.S. Diagnostic performance of commercial IgM and IgG enzyme-linked immunoassays (ELISAs) for diagnosis of Zika virus infection. Virol. J. 2018, 15, 108. [Google Scholar] [CrossRef] [PubMed]
- Bozza, F.A.; Moreira-Soto, A.; Rockstroh, A.; Fischer, C.; Nascimento, A.D.; Calheiros, A.S.; Drosten, C.; Bozza, P.T.; Souza, T.M.L.; Ulbert, S.; et al. Differential Shedding and Antibody Kinetics of Zika and Chikungunya Viruses, Brazil. Emerg. Infect. Dis. 2019, 25, 311–315. [Google Scholar] [CrossRef] [PubMed]
- Shu, P.Y.; Chen, L.K.; Chang, S.F.; Yueh, Y.Y.; Chow, L.; Chien, L.J.; Chin, C.; Lin, T.H.; Huang, J.H. Dengue NS1-specific antibody responses: Isotype distribution and serotyping in patients with Dengue fever and Dengue hemorrhagic fever. J. Med. Virol. 2000, 62, 224–232. [Google Scholar] [CrossRef]
- Vázquez, S.; Pérez, A.B.; Ruiz, D.; Rodríguez, R.; Pupo, M.; Calzada, N.; González, L.; González, D.; Castro, O.; Serrano, T.; et al. Serological markers during dengue 3 primary and secondary infections. J. Clin. Virol. Off. Publ. Pan Am. Soc. Clin. Virol. 2005, 33, 132–137. [Google Scholar] [CrossRef]
- Groen, J.; Velzing, J.; Copra, C.; Balentien, E.; Deubel, V.; Vorndam, V.; Osterhaus, A.D. Diagnostic value of dengue virus-specific IgA and IgM serum antibody detection. Microbes Infect. 1999, 1, 1085–1090. [Google Scholar] [CrossRef]
- Zhao, L.Z.; Hong, W.X.; Wang, J.; Yu, L.; Hu, F.Y.; Qiu, S.; Yin, C.B.; Tang, X.P.; Zhang, L.Q.; Jin, X.; et al. Kinetics of antigen-specific IgM/IgG/IgA antibody responses during Zika virus natural infection in two patients. J. Med. Virol. 2019, 91, 872–876. [Google Scholar] [CrossRef]
- Yamanaka, A.; Matsuda, M.; Okabayashi, T.; Pitaksajjakul, P.; Ramasoota, P.; Saito, K.; Fukasawa, M.; Hanada, K.; Matsuura, T.; Muramatsu, M.; et al. Seroprevalence of Flavivirus Neutralizing Antibodies in Thailand by High-Throughput Neutralization Assay: Endemic Circulation of Zika Virus before 2012. mSphere 2021, 6, e0033921. [Google Scholar] [CrossRef]
- Barros, J.B.S.; da Silva, P.A.N.; Koga, R.C.R.; Gonzalez-Dias, P.; Carmo Filho, J.R.; Nagib, P.R.A.; Coelho, V.; Nakaya, H.I.; Fonseca, S.G.; Pfrimer, I.A.H. Acute Zika Virus Infection in an Endemic Area Shows Modest Proinflammatory Systemic Immunoactivation and Cytokine-Symptom Associations. Front. Immunol. 2018, 9, 821. [Google Scholar] [CrossRef]
- Rathakrishnan, A.; Wang, S.M.; Hu, Y.; Khan, A.M.; Ponnampalavanar, S.; Lum, L.C.; Manikam, R.; Sekaran, S.D. Cytokine expression profile of dengue patients at different phases of illness. PLoS ONE 2012, 7, e52215. [Google Scholar] [CrossRef]
- Sánchez-Arcila, J.C.; Badolato-Correa, J.; de Souza, T.M.A.; Paiva, I.A.; Barbosa, L.S.; Nunes, P.C.G.; Lima, M.; Dos Santos, F.B.; Damasco, P.V.; da Cunha, R.V.; et al. Clinical, Virological, and Immunological Profiles of DENV, ZIKV, and/or CHIKV-Infected Brazilian Patients. Intervirology 2020, 63, 33–45. [Google Scholar] [CrossRef] [PubMed]
- Le Page, C.; Génin, P.; Baines, M.G.; Hiscott, J. Interferon activation and innate immunity. Rev. Immunogenet. 2000, 2, 374–386. [Google Scholar] [PubMed]
- Liu, M.; Guo, S.; Hibbert, J.M.; Jain, V.; Singh, N.; Wilson, N.O.; Stiles, J.K. CXCL10/IP-10 in infectious diseases pathogenesis and potential therapeutic implications. Cytokine Growth Factor Rev. 2011, 22, 121–130. [Google Scholar] [CrossRef]
- Wei, L.; Laurence, A.; Elias, K.M.; O’Shea, J.J. IL-21 is produced by Th17 cells and drives IL-17 production in a STAT3-dependent manner. J. Biol. Chem. 2007, 282, 34605–34610. [Google Scholar] [CrossRef] [PubMed]
- Wan, Z.; Zhou, Z.; Liu, Y.; Lai, Y.; Luo, Y.; Peng, X.; Zou, W. Regulatory T cells and T helper 17 cells in viral infection. Scand. J. Immunol. 2020, 91, e12873. [Google Scholar] [CrossRef]
- Priyadarshini, D.; Gadia, R.R.; Tripathy, A.; Gurukumar, K.R.; Bhagat, A.; Patwardhan, S.; Mokashi, N.; Vaidya, D.; Shah, P.S.; Cecilia, D. Clinical findings and pro-inflammatory cytokines in dengue patients in Western India: A facility-based study. PLoS ONE 2010, 5, e8709. [Google Scholar] [CrossRef] [PubMed]
- Pandey, N.; Jain, A.; Garg, R.K.; Kumar, R.; Agrawal, O.P.; Lakshmana Rao, P.V. Serum levels of IL-8, IFNγ, IL-10, and TGF β and their gene expression levels in severe and non-severe cases of dengue virus infection. Arch. Virol. 2015, 160, 1463–1475. [Google Scholar] [CrossRef]
- Bhatt, P.; Sabeena, S.P.; Varma, M.; Arunkumar, G. Current Understanding of the Pathogenesis of Dengue Virus Infection. Curr. Microbiol. 2021, 78, 17–32. [Google Scholar] [CrossRef]
- Tappe, D.; Pérez-Girón, J.V.; Zammarchi, L.; Rissland, J.; Ferreira, D.F.; Jaenisch, T.; Gómez-Medina, S.; Günther, S.; Bartoloni, A.; Muñoz-Fontela, C.; et al. Cytokine kinetics of Zika virus-infected patients from acute to reconvalescent phase. Med. Microbiol. Immunol. 2016, 205, 269–273. [Google Scholar] [CrossRef]
- Restrepo, B.N.; Ramirez, R.E.; Arboleda, M.; Alvarez, G.; Ospina, M.; Diaz, F.J. Serum levels of cytokines in two ethnic groups with dengue virus infection. Am. J. Trop. Med. Hyg. 2008, 79, 673–677. [Google Scholar] [CrossRef] [PubMed]
- Subramaniam, K.S.; Lant, S.; Goodwin, L.; Grifoni, A.; Weiskopf, D.; Turtle, L. Two Is Better Than One: Evidence for T-Cell Cross-Protection Between Dengue and Zika and Implications on Vaccine Design. Front. Immunol. 2020, 11, 517. [Google Scholar] [CrossRef]
- Elong Ngono, A.; Shresta, S. Cross-Reactive T Cell Immunity to Dengue and Zika Viruses: New Insights Into Vaccine Development. Front. Immunol. 2019, 10, 1316. [Google Scholar] [CrossRef]
- Gourinat, A.C.; O’Connor, O.; Calvez, E.; Goarant, C.; Dupont-Rouzeyrol, M. Detection of Zika virus in urine. Emerg. Infect. Dis. 2015, 21, 84–86. [Google Scholar] [CrossRef] [PubMed]
- Screaton, G.; Mongkolsapaya, J.; Yacoub, S.; Roberts, C. New insights into the immunopathology and control of dengue virus infection. Nat. Rev. Immunol. 2015, 15, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Rivino, L.; Kumaran, E.A.; Thein, T.L.; Too, C.T.; Gan, V.C.; Hanson, B.J.; Wilder-Smith, A.; Bertoletti, A.; Gascoigne, N.R.; Lye, D.C.; et al. Virus-specific T lymphocytes home to the skin during natural dengue infection. Sci. Transl. Med. 2015, 7, 278ra235. [Google Scholar] [CrossRef]
- Lee, A.; Park, S.P.; Park, C.H.; Kang, B.H.; Park, S.H.; Ha, S.J.; Jung, K.C. IL-4 Induced Innate CD8+ T Cells Control Persistent Viral Infection. PLoS Pathog. 2015, 11, e1005193. [Google Scholar] [CrossRef]
- Ozaki, K.; Spolski, R.; Feng, C.G.; Qi, C.F.; Cheng, J.; Sher, A.; Morse, H.C., 3rd; Liu, C.; Schwartzberg, P.L.; Leonard, W.J. A critical role for IL-21 in regulating immunoglobulin production. Science 2002, 298, 1630–1634. [Google Scholar] [CrossRef] [PubMed]
- Konforte, D.; Simard, N.; Paige, C.J. IL-21: An executor of B cell fate. J. Immunol. 2009, 182, 1781–1787. [Google Scholar] [CrossRef]
- Rojas, J.M.; Avia, M.; Martín, V.; Sevilla, N. IL-10: A Multifunctional Cytokine in Viral Infections. J. Immunol. Res. 2017, 2017, 6104054. [Google Scholar] [CrossRef]
- Pérez, A.B.; García, G.; Sierra, B.; Alvarez, M.; Vázquez, S.; Cabrera, M.V.; Rodríguez, R.; Rosario, D.; Martínez, E.; Denny, T.; et al. IL-10 levels in Dengue patients: Some findings from the exceptional epidemiological conditions in Cuba. J. Med. Virol. 2004, 73, 230–234. [Google Scholar] [CrossRef]
- Stewart, C.A.; Metheny, H.; Iida, N.; Smith, L.; Hanson, M.; Steinhagen, F.; Leighty, R.M.; Roers, A.; Karp, C.L.; Müller, W.; et al. Interferon-dependent IL-10 production by Tregs limits tumor Th17 inflammation. J. Clin. Investig. 2013, 123, 4859–4874. [Google Scholar] [CrossRef] [PubMed]
- Brandt, E.B.; Sivaprasad, U. Th2 Cytokines and Atopic Dermatitis. J. Clin. Cell. Immunol. 2011, 2, 110. [Google Scholar] [CrossRef]
- Geha, R.S.; Jabara, H.H.; Brodeur, S.R. The regulation of immunoglobulin E class-switch recombination. Nat. Rev. Immunol. 2003, 3, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Yao, J.; Li, B. Expression of TSLP and Downstream Molecules IL-4, IL-5, and IL-13 on the Eye Surface of Patients with Various Types of Allergic Conjunctivitis. J. Ophthalmol. 2016, 2016, 5072781. [Google Scholar] [CrossRef] [PubMed]
- Katial, R.K.; Bensch, G.W.; Busse, W.W.; Chipps, B.E.; Denson, J.L.; Gerber, A.N.; Jacobs, J.S.; Kraft, M.; Martin, R.J.; Nair, P.; et al. Changing Paradigms in the Treatment of Severe Asthma: The Role of Biologic Therapies. J. Allergy Clin. Immunol. Pr. 2017, 5, S1–S14. [Google Scholar] [CrossRef]
- Hiraoka, N.; Yokote, T.; Nakayama-Ichiyama, S.; Takayama, A.; Iwaki, K.; Kobayashi, K.; Oka, S.; Miyoshi, T.; Akioka, T.; Takubo, T.; et al. High fever and shock induced by interferon-γ and interleukin-6 produced by adult T-cell leukaemia/lymphoma cells. Leuk. Res. 2010, 34, e290–e291. [Google Scholar] [CrossRef]
- Fajgenbaum, D.C.; June, C.H. Cytokine Storm. N. Engl. J. Med. 2020, 383, 2255–2273. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics of Samples | Microbiology and Immunology | Tropical Pediatrics | Total |
|---|---|---|---|
| Subject number (n) | 71 | 30 | 101 |
| Male:Female | 0.82:1 | 1:1 | 0.87:1 |
| Age (years old) * | 36 | 9 | 27.41 ± 17.37 (28, 4–68) |
| Day of fever * | 3 | 2 | 2.90 ± 1.49 (3, 1–7) |
| Clinical symptoms (n = 70): | |||
| - Fever | 58 (83%) | NA | 58 (83%) |
| - Sore throat | 18 (26%) | NA | 18 (26%) |
| - Cough | 14 (20%) | NA | 14 (20%) |
| - Headache | 20 (29%) | NA | 20 (29%) |
| - Exanthema | 59 (84%) | NA | 59 (84%) |
| - Conjunctivitis | 19 (27%) | NA | 19 (27%) |
| - Arthralgia | 14 (20%) | NA | 14 (20%) |
| - Myalgia | 23 (33%) | NA | 23 (33%) |
| Sample types: | |||
| Acute sample | 71 | 30 ** | 101 |
| Convalescent sample | 0 | 30 ** | 30 |
| Dengue exposure cases ***: | |||
| Dengue IgG+ | 55 (77%) | 17 (57%) | 72 (71%) |
| Dengue IgG− | 16 (23%) | 13 (43%) | 29 (29%) |
| Characteristics of Samples | Dengue Patients * | Healthy Donors (n = 10) | |
|---|---|---|---|
| Acute (n = 5) | Convalescent (n = 10) | ||
| Dengue IgM+ | 5 (100%) | 10 (100%) | NA |
| Dengue IgM− | 0 | 0 | NA |
| Dengue IgG+ | 0 | 10 (100%) | NA |
| Dengue IgG− | 5 (100%) | 0 | NA |
| Flavivirus IgM+ | NA | NA | 0 |
| Flavivirus IgM− | NA | NA | 10 (100%) |
| Flavivirus IgG+ | NA | NA | 0 |
| Flavivirus IgG− | NA | NA | 10 (100%) |
| Type of Samples/Cases | DENV IgG | Euroimmun ZIKV NS1 ELISAs | ||
|---|---|---|---|---|
| IgA+/IgM+ | IgM+ | IgA+ | ||
| Acute ZIKV samples | Positive (n = 72) | 10/72 (14%) | 2/72 (3%) | 3/69 (4%) |
| Negative (n = 29) | 0/29 (0%) | 0/29 (0%) | 0/27 (0%) | |
| Total (n = 101) | 10/101 (10%) | 2/101 (2%) | 3/96 * (3%) | |
| Convalescent ZIKV samples | Positive (n = 17) | 0/17 (0%) | 0/17 (0%) | 0/17 (0%) |
| Negative (n = 13) | 1/13 (8%) | 0/13 (0%) | 1/13 (8%) | |
| Total (n = 30) | 1/30 (3%) | 0/30 (0%) | 1/30 (3%) | |
| ZIKV patients ** | Positive (n = 72) | 10/72 (14%) | 2/72 (3%) | 3/69 (4%) |
| Negative (n = 29) | 1/29 (3%) | 0/29 (0%) | 1/27 (4%) | |
| Total (n = 101) | 11/101 (11%) | 2/101 (2%) | 4/96 * (4%) | |
| Acute DENV samples | Total (n = 5) | 0/5 (0%) | 0/5 (0%) | NA |
| Convalescent DENV samples | Total (n = 10) | 0/10 (0%) | 0/10 (0%) | NA |
| Healthy samples | Total (n = 10) | 0/10 (0%) | 0/10 (0%) | 0/10 (0%) |
| Cytokine (pg/mL) | Type of Serum Samples | ||||
|---|---|---|---|---|---|
| Acute ZIKV (n = 70) | Convalescent ZIKV (n = 18) | Acute DENV (n = 5) | Convalescent DENV (n = 5) | Healthy Donor (n = 8) | |
| IFN-γ | 14.54 ± 50.87/6.08 5.32 (0–422.70) 2.41, 26.67 | 1.13 ± 1.65/0.39 0 (0–4.49) 0.31, 1.95 | 3.64 ± 3.18/1.42 1.53 (1.10–7.82) −0.31, 7.59 | 8.52 ± 13.99/6.25 3.81 (0–33.15) −8.85, 25.88 | 0 ± 0/0 0 (0–0) 0, 0 |
| IL-4 | 41.85 ± 24.85/2.97 40.44 (0–106.30) 35.93, 47.78 | 7.90 ± 8.77/2.07 4.41 (0–28.81) 3.54, 12.26 | 16.79 ± 1.54/0.69 17.28 (14.60–18.42) 14.87, 18.70 | 15.50 ± 7.66/3.43 19.43 (7.08–22.72) 5.99, 25.02 | 9.14 ± 6.22/2.20 10.88 (0–17.39) 3.94, 14.33 |
| IL-5 | 12.90 ± 12.03/1.44 9.69 (0–58.09) 10.03, 15.76 | 9.12 ± 13.54/3.19 0 (0–44.85) 2.39, 15.85 | 18.78 ± 11.80/5.28 18.59 (0–31.60) 4.12, 33.43 | 8.74 ± 12.18/5.45 0 (0–25.10) −6.39, 23.87 | 0 ± 0/0 0 (0–0) 0, 0 |
| IL-6 | 6.91 ± 12.02/1.44 2.89 (0–77.54) 4.05, 9.78 | 3.16 ± 5.60/1.32 0 (0–18.06) 0.38, 5.94 | 4.96 ± 9.68/4.33 0.60 (0–22.21) −7.06, 16.97 | 0 ± 0/0 0 (0–0) 0, 0 | 3.67 ± 4.42/1.56 2.70 (0–11.93) −0.03, 7.37 |
| IL-10 | 4.26 ± 7.06/0.84 2.44 (0.24–55.16) 2.58, 5.95 | 0.54 ± 0.63/0.15 0.35 (0–2.64) 0.23, 0.85 | 25.20 ± 24.77/11.08 14.73 (5.47–64.38) −5.56, 55.95 | 24.35 ± 23.99/10.73 13.08 (3.13–56.09) −5.43, 54.14 | 0.63 ± 0.68/0.24 0.55 (0–2.18) 0.05, 1.20 |
| IL-21 | 11.01 ± 32.27/3.86 0 (0–184.3) 3.32, 18.71 | 40.84 ± 97.16/22.90 2.07 (0–347.10) −7.48, 89.16 | 5.58 ± 10.76/4.81 0 (0–24.66) −7.78, 18.93 | 1.84 ± 2.69/1.20 0 (0–5.95) −1.51, 5.18 | 12.18 ± 23.18/8.20 0 (0–66.86) −7.20, 31.56 |
| CXCL10 | 186.2 ± 170.2/20.35 135.2 (13.53–852.4) 145.60, 226.80 | 39.92 ± 27.69/6.53 29.13 (14.82–124.1) 26.15, 53.69 | 131.3 ± 33.16/14.83 137.2 (93.19–167.8) 90.17, 172.50 | 237.9 ± 132.7/59.34 293.0 (95.63–372.6) 73.11, 402.60 | 49.65 ± 17.52/6.19 51.05 (13.72–75.47) 35.00, 64.29 |
| Cytokine | p-Value a Derived from the Comparison between Each Group of Serum Samples | |||||
|---|---|---|---|---|---|---|
| Acute ZIKV vs. Healthy Donor | Convalescent ZIVK vs. Healthy Donor | Acute DENV vs. Healthy Donor | Convalescent DENV vs. Healthy Donor | Acute ZIKV vs. Acute DENV | Convalescent ZIKV vs. Convalescent DENV | |
| IFN-γ | <0.0001; **** | 0.0721; ns | 0.0008; *** | 0.0350; * | 0.3361; ns | 0.1381; ns |
| IL-4 | <0.0001; **** | 0.6726; ns | 0.0109; * | 0.2688; ns | 0.0176; * | 0.0909; ns |
| IL-5 | <0.0001; **** | 0.0696; ns | 0.0070; ** | 0.1282; ns | 0.2132; ns | 0.9026; ns |
| IL-6 | 0.5928; ns | 0.6948; ns | >0.9999; ns | 0.1608; ns | 0.7391; ns | 0.2006; ns |
| IL-10 | <0.0001; **** | 0.4187; ns | 0.0008; *** | 0.0008; *** | 0.0003; *** | <0.0001; **** |
| IL-21 | 0.9495; ns | 0.7600; ns | >0.9999; ns | 0.6954; ns | 0.8012; ns | 0.3794; ns |
| CXCL10 | 0.0002; *** | 0.1285; ns | 0.0016; ** | 0.0016; ** | 0.9099; ns | 0.0002; *** |
| Cytokine (pg/mL) p-Value a | Cytokine Levels among Individuals with the Absence or Presence of Clinical Symptoms | |||||||
|---|---|---|---|---|---|---|---|---|
| Fever | Sore Throat | Cough | Headache | Exanthema | Conjunctivitis | Arthralgia | Myalgia | |
| Absence (n = 18) vs. Presence (n = 50) | Absence (n = 55) vs. Presence (n = 13) | Absence (n = 56) vs. Presence (n = 12) | Absence (n = 49) vs. Presence (n = 19) | Absence (n = 19) vs. Presence (n = 49) | Absence (n = 52) vs. Presence (n = 16) | Absence (n = 55) vs. Presence (n = 13) | Absence (n = 49) vs. Presence (n = 19) | |
| IFN-γ | 0.77 (0–10.83) vs. 5.97 (0–422.70) <0.0001; **** | 3.61 (0–422.7) vs. 6.42 (0–35.88) 0.2585; ns | 3.81 (0–422.7) vs. 3.58 (0–48.29) 0.9081; ns | 3.61 (0–56.64) vs. 7.83 (0–422.7) 0.1010; ns | 0.67 (0–422.7) vs. 5.63 (0–50.66) 0.0017; ** | 3.41 (0–422.7) vs. 8.23 (0.9–29.94) 0.0306; * | 3.41 (0–422.7) vs. 6.42 (0–29.94) 0.2455; ns | 3.61 (0–422.7) vs. 6.12 (0–56.64) 0.1131; ns |
| IL-4 | 15.18 (0–69.94) vs. 45.1 (4–106.3) 0.0106; * | 26.49 (0–106.3) vs. 55.78 (6.0–78.7) 0.0671; ns | 39.29 (0–106.3) vs. 31.4 (3.9–85.1) 0.9079; ns | 38.34 (0–106.3) vs. 38.82 (6.3–92.84) 0.6473; ns | 17.43 (0–106.3) vs. 49.77 (6.0–92.84) 0.0018; ** | 23.6 (0–106.3) vs. 57.24 (11–92.84) 0.0112; * | 29.94 (0–106.3) vs. 57.81 (3.89–92.84) 0.0388; * | 28.58 (0–92.84) vs. 46.89 (4–106.3) 0.1381; ns |
| IL-5 | 0 (0–25.10) vs. 10.87 (0–58.09) 0.0018; ** | 8.53 (0–58.09) vs. 9.70 (0–31.60) 0.2189; ns | 8.53 (0–58.09) vs. 10.28 (0–38.34) 0.9841; ns | 8.53 (0.0–58.09) vs. 12.03 (0–38.34) 0.0496; * | 0 (0–31.60) vs. 10.87 (0–58.09) 0.0047; ** | 8.53 (0–44.85) vs. 10.28 (0–58.09) 0.2223; ns | 8.53 (0–58.09) vs. 12.02 (0–38.34) 0.3942; ns | 8.53 (0–44.85) vs. 12.02 (0–58.09) 0.1299; ns |
| IL-6 | 2.52 (0–11.93) vs. 2.13 (0–77.54) 0.5793; ns | 1.63 (0–77.54) vs. 5.04 (0–20.89) 0.5512; ns | 2.13 (0–77.54) vs. 3.61 (0–20.89) 0.5558; ns | 1.41 (0–37.86) vs. 3.62 (0–77.54) 0.4603; ns | 5.40 (0–77.54) vs. 1.63 (0–28.84) 0.414; ns | 0 (0–77.54) vs. 4.57 (0–28.84) 0.2589; ns | 1.13 (0–77.54) vs. 5.04 (0–28.84) 0.5250; ns | 1.63 (0–77.54) vs. 3.62 (0–37.86) 0.6509; ns |
| IL-10 | 1.35 (0–4.43) vs. 3.35 (0.2–55.2) 0.0004; *** | 2.18 (0–55.16) vs. 3.92 (1.7–10.7) 0.0857; ns | 2.44 (0–55.16) vs. 3.59 (0.3–12.1) 0.4476; ns | 2.55 (0–55.16) vs. 2.50 (0.24–16.5) 0.3001; ns | 2.28 (0–55.16) vs. 2.58 (0.24–16.50) 0.632; ns | 2.995 (0–55.16) vs. 1.87 (0.90–12.14) 0.5214; ns | 2.55 (0–55.16) vs. 1.61 (0.57–12.14) 0.5554; ns | 2.18 (0–16.5) vs. 4.44 (1.3–55.2) 0.0114; * |
| IL-21 | 0 (0–66.86) vs. 0.70 (0–138.90) 0.5181; ns | 0 (0–138.9) vs. 3.83 (0–18.51) 0.7609; ns | 0 (0–66.86) vs. 2.05 (0–138.9) 0.6518; ns | 2.32 (0–138.90) vs. 0 (0–10.58) 0.2113; ns | 0 (0–138.9) vs. 0 (0–46.8) 0.9795; ns | 0 (0–138.9) vs. 0 (0–46.80) 0.9228; ns | 0 (0–138.9) vs. 4.10 (0–21.63) 0.1638; ns | 0 (0–66.86) vs. 2.32 (0–138.9) 0.4379; ns |
| CXCL10 | 76.0 (14–525.6) vs. 140.9 (13.5–852) 0.0914; ns | 120.4 (14–852.4) vs. 163.0 (46–727.6) 0.2267; ns | 120.2 (14–852.4) vs. 169.3 (14–726.1) 0.4122; ns | 120.6 (14–852.4) vs. 126.0 (23–399.2) 0.8922; ns | 64.36 (13.5–852.4) vs. 151.2 (22.8–727.6) 0.0029; ** | 100.6 (13.5–852.4) vs. 156.2 (65.4–489.8) 0.0332; * | 120.0 (13.7–852.4) vs. 143.2 (13.5–489.8) 0.5169; ns | 115.7 (14–727.6) vs. 151.2 (14–852.4) 0.2649; ns |
| Correlations among Immune Mediators in Acute ZIKV Subjects (n = 70) | |||||||
|---|---|---|---|---|---|---|---|
| rs 95% CI p-Value a | IFN-γ | IL-4 | IL-5 | IL-6 | IL-10 | IL-21 | CXCL10 |
| IFN-γ | 1 | ||||||
| IL-4 | 0.6038 0.42, 0.74 <0.001; *** | 1 | |||||
| IL-5 | 0.4017 0.18, 0.59 <0.001; *** | 0.1923 −0.05, 0.41 0.111; ns | 1 | ||||
| IL-6 | 0.4282 0.21, 0.61 <0.001; *** | 0.4676 0.25, 0.64 <0.001; *** | 0.2123 −0.03, 0.43 0.078; ns | 1 | |||
| IL-10 | 0.5266 0.33, 0.68 <0.001; *** | 0.2271 −0.02, 0.44 0.059; ns | 0.3990 0.17, 0.58 <0.001; *** | 0.1961 −0.05, 0.42 0.104; ns | 1 | ||
| IL-21 | 0.2019 −0.04, 0.42 0.094; ns | 0.4730 0.26, 0.64 <0.001; *** | 0.2663 0.03, 0.48 0.026; * | 0.3873 0.16, 0.58 <0.001; *** | 0.1770 −0.07, 0.40 0.143; ns | 1 | |
| CXCL10 | 0.4175 0.20, 0.60 <0.001; *** | 0.6452 0.48, 0.77 <0.001; *** | 0.1417 −0.10, 0.37 0.242; ns | 0.3716 0.14, 0.56 0.002; ** | 0.3603 0.13, 0.55 0.002; ** | 0.4182 0.20, 0.60 <0.001; *** | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petphong, V.; Kosoltanapiwat, N.; Limkittikul, K.; Maneekan, P.; Chatchen, S.; Jittmittraphap, A.; Sriburin, P.; Chattanadee, S.; Leaungwutiwong, P. Detection of Anti-ZIKV NS1 IgA, IgM, and Combined IgA/IgM and Identification of IL-4 and IL-10 as Potential Biomarkers for Early ZIKV and DENV Infections in Hyperendemic Regions, Thailand. Trop. Med. Infect. Dis. 2023, 8, 284. https://doi.org/10.3390/tropicalmed8050284
Petphong V, Kosoltanapiwat N, Limkittikul K, Maneekan P, Chatchen S, Jittmittraphap A, Sriburin P, Chattanadee S, Leaungwutiwong P. Detection of Anti-ZIKV NS1 IgA, IgM, and Combined IgA/IgM and Identification of IL-4 and IL-10 as Potential Biomarkers for Early ZIKV and DENV Infections in Hyperendemic Regions, Thailand. Tropical Medicine and Infectious Disease. 2023; 8(5):284. https://doi.org/10.3390/tropicalmed8050284
Chicago/Turabian StylePetphong, Vajee, Nathamon Kosoltanapiwat, Kriengsak Limkittikul, Pannamas Maneekan, Supawat Chatchen, Akanitt Jittmittraphap, Pimolpachr Sriburin, Siriporn Chattanadee, and Pornsawan Leaungwutiwong. 2023. "Detection of Anti-ZIKV NS1 IgA, IgM, and Combined IgA/IgM and Identification of IL-4 and IL-10 as Potential Biomarkers for Early ZIKV and DENV Infections in Hyperendemic Regions, Thailand" Tropical Medicine and Infectious Disease 8, no. 5: 284. https://doi.org/10.3390/tropicalmed8050284