
A Histone Deacetylase Inhibitor Manifests Synergistic Interaction with Artesunate by Suppressing DNA Repair Activity

Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Cell Culture and Drug Treatment
2.3. Assessment of Cell Viability
2.4. HDAC Inhibition Assay
2.5. Propidium Iodide (PI) Apoptosis Assay
2.6. Fluoremetric Analyses of Apoptotic Cells Using PE Annexin V and 7-AAD
2.7. Combination Index (CI) Curve and Isobologram Analyses
2.8. Intracellular ROS Measurement
2.9. Cell Cycle Analysis
2.10. Relative mRNA Expression Analysis
2.11. In Vivo Study
2.12. Dalton’s Ascites Lymphoma Development in Mice
2.13. In Vivo Experimental Design
2.14. Histopathological Analysis
2.15. Cell Viability Assay
2.16. Statistical Analysis
3. Results
3.1. 4-PB Cytotoxicity on MCF-7 and MDA-MB-231 Cells as a Consequence of HDAC Activity Inhibition
3.2. 4-PB Enhanced the Dose-Dependent Cytotoxicity of ART
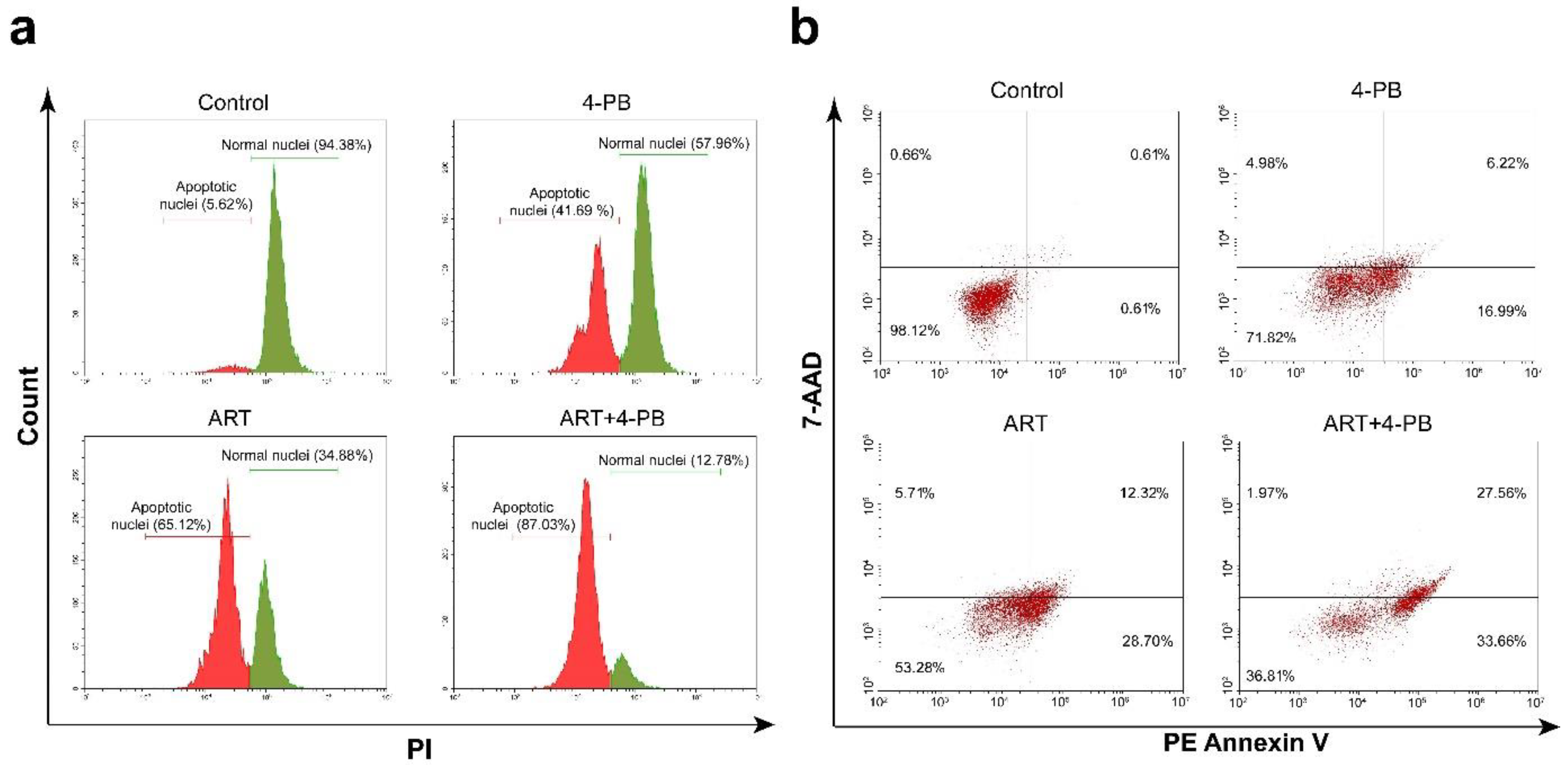
3.3. Induction of Apoptosis by ART and/or 4-PB
3.4. Isobologram Depicted the Interaction of 4-PB with ART Is Synergistic
3.5. 4-PB and ART Increased Oxidative Stress in MCF7 Cells
3.6. Cell Cycle Perturbation after Treatment with ART and/or 4-PB
3.7. Mechanism Underlying 4-PB Mediated Enhanced ART DNA Damaging Effect
3.8. Therapeutic Effectivity of Combinated Drusg in a Murine Model
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethics Approval
References
- Sakuma, T.; Uzawa, K.; Onda, T.; Shiiba, M.; Yokoe, H.; Shibahara, T.; Tanzawa, H. Aberrant expression of histone deacetylase 6 in oral squamous cell carcinoma. Int. J. Oncol. 2006, 29, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Attrish, D.; Srivastava, A.; Banerjee, J.; Tripathi, M.; Chandra, P.S.; Dixit, A.B. Non-histone substrates of histone deacetylases as potential therapeutic targets in epilepsy. Expert Opin. Ther. Targets 2021, 25, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Stypula-Cyrus, Y.; Damania, D.; Kunte, D.P.; Cruz, M.D.; Subramanian, H.; Roy, H.K.; Backman, V. HDAC up-regulation in early colon field carcinogenesis is involved in cell tumorigenicity through regulation of chromatin structure. PLoS ONE 2013, 8, e64600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peart, M.J.; Smyth, G.K.; Van Laar, R.K.; Bowtell, D.D.; Richon, V.M.; Marks, P.A.; Holloway, A.J.; Johnstone, R.W. Identification and functional significance of genes regulated by structurally different histone deacetylase inhibitors. Proc. Natl. Acad. Sci. USA 2005, 102, 3697–3702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, W.K.; O’Connor, O.A.; Marks, P.A. Histone deacetylase inhibitors: From target to clinical trials. Expert Opin. Investig. Drugs 2002, 11, 1695–1713. [Google Scholar] [CrossRef]
- Brusilow, S.W.; Finkelstien, J. Restoration of nitrogen homeostasis in a man with ornithine transcarbamylase deficiency. Metabolism 1993, 42, 1336–1339. [Google Scholar] [CrossRef]
- Rubenstein, R.C.; Zeitlin, P.L. A Pilot Clinical Trial of Oral Sodium 4-Phenylbutyrate (Buphenyl) in Δ F508-Homozygous Cystic Fibrosis Patients: Partial Restoration of Nasal Epithelial CFTR function. Am. J. Respir. Crit. Care Med. 1998, 157, 484–490. [Google Scholar] [CrossRef] [PubMed]
- Munshi, A.; Kurland, J.F.; Nishikawa, T.; Tanaka, T.; Hobbs, M.L.; Tucker, S.L.; Ismail, S.; Stevens, C.; Meyn, R.E. Histone deacetylase inhibitors radiosensitize human melanoma cells by suppressing DNA repair activity. Clin. Cancer Res. 2005, 11, 4912–4922. [Google Scholar] [CrossRef] [Green Version]
- Rosenthal, P.J. Artesunate for the treatment of severe falciparum malaria. N. Engl. J. Med. 2008, 358, 1829–1836. [Google Scholar] [CrossRef] [Green Version]
- Crespo-Ortiz, M.P.; Wei, M.Q. Antitumor activity of artemisinin and its derivatives: From a well-known antimalarial agent to a potential anticancer drug. J. Biomed. Biotechnol. 2012, 2012, 247597. [Google Scholar] [CrossRef] [Green Version]
- Raza, A.; Ghoshal, A.; Chockalingam, S.; Ghosh, S.S. Connexin-43 enhances tumor suppressing activity of artesunate via gap junction-dependent as well as independent pathways in human breast cancer cells. Sci. Rep. 2017, 7, 7580. [Google Scholar] [CrossRef] [PubMed]
- Hosoya, N.; Miyagawa, K. Targeting DNA damage response in cancer therapy. Cancer Sci. 2014, 105, 370–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gordi, T.; Lepist, E.-I. Artemisinin derivatives: Toxic for laboratory animals, safe for humans? Toxicol. Lett. 2004, 147, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Raza, A.; Kohila, V.; Ghosh, S.S. Redesigned Escherichia coli cytosine deaminase: A new facet of suicide gene therapy. J. Gene Med. 2015, 17, 132–139. [Google Scholar] [CrossRef]
- Raza, A.; Ghosh, S.S. Connexin-43 Enhances the Redesigned Cytosine Deaminase Activity for Suicide Gene Therapy in Human Breast Cancer Cells. Biochem. Insights 2019, 12, 1178626418818182. [Google Scholar] [CrossRef]
- Goswami, U.; Dutta, A.; Raza, A.; Kandimalla, R.; Kalita, S.; Ghosh, S.S.; Chattopadhyay, A. Transferrin–copper nanocluster–doxorubicin nanoparticles as targeted theranostic cancer Nanodrug. ACS Appl. Mater. Interfaces 2018, 10, 3282–3294. [Google Scholar] [CrossRef]
- Raza, A.; Singh, A.; Amin, S.; Spallholz, J.E.; Sharma, A.K. Identification and biotin receptor-mediated activity of a novel seleno-biotin compound that inhibits viability of and induces apoptosis in ovarian cancer cells. Chem.-Biol. Interact. 2022, 365, 110071. [Google Scholar] [CrossRef]
- Ghoshal, A.; Goswami, U.; Raza, A.; Chattopadhyay, A.; Ghosh, S.S. Recombinant sFRP4 bound chitosan–alginate composite nanoparticles embedded with silver nanoclusters for Wnt/β-catenin targeting in cancer theranostics. RSC Adv. 2016, 6, 85763–85772. [Google Scholar] [CrossRef]
- Gore, S.D.; Carducci, M.A. Modifying histones to tame cancer: Clinical development of sodium phenylbutyrate and other histone deacetylase inhibitors. Expert Opin. Investig. Drugs 2000, 9, 2923–2934. [Google Scholar] [CrossRef]
- Benakis, A.; Paris, M.; Loutan, L.; Plessas, C.T.; Plessas, S.T. Pharmacokinetics of artemisinin and artesunate after oral administration in healthy volunteers. Am. J. Trop. Med. Hyg. 1997, 56, 17–23. [Google Scholar] [CrossRef]
- Fishwick, J.; McLean, W.G.; Edwards, G.; Ward, S.A. The toxicity of artemisinin and related compounds on neuronal and glial cells in culture. Chem.-Biol. Interact. 1995, 96, 263–271. [Google Scholar] [CrossRef]
- Batshaw, M.L.; MacArthur, R.B.; Tuchman, M. Alternative pathway therapy for urea cycle disorders: Twenty years later. J. Pediatrics 2001, 138, S46–S55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ververis, K.; Hiong, A.; Karagiannis, T.C.; Licciardi, P.V. Histone deacetylase inhibitors (HDACIs): Multitargeted anticancer agents. Biol. Targets Ther. 2013, 7, 47. [Google Scholar]
- Smith, K.T.; Workman, J.L. Histone deacetylase inhibitors: Anticancer compounds. Int. J. Biochem. Cell Biol. 2009, 41, 21–25. [Google Scholar] [CrossRef] [PubMed]
- Blagosklonny, M.V.; Robey, R.; Sackett, D.L.; Du, L.; Traganos, F.; Darzynkiewicz, Z.; Fojo, T.; Bates, S.E. Histone deacetylase inhibitors all induce p21 but differentially cause tubulin acetylation, mitotic arrest, and cytotoxicity. Mol. Cancer Ther. 2002, 1, 937–941. [Google Scholar] [PubMed]
- Goh, M.; Chen, F.; Paulsen, M.T.; Yeager, A.M.; Dyer, E.S.; Ljungman, M. Phenylbutyrate attenuates the expression of Bcl-X(L), DNA-PK, caveolin-1, and VEGF in prostate cancer cells. Neoplasia 2001, 3, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Berdelle, N.; Nikolova, T.; Quiros, S.; Efferth, T.; Kaina, B. Artesunate induces oxidative DNA damage, sustained DNA double-strand breaks, and the ATM/ATR damage response in cancer cells. Mol. Cancer Ther. 2011, 10, 2224–2233. [Google Scholar] [CrossRef] [Green Version]
- Li, P.C.; Lam, E.; Roos, W.P.; Zdzienicka, M.Z.; Kaina, B.; Efferth, T. Artesunate derived from traditional Chinese medicine induces DNA damage and repair. Cancer Res. 2008, 68, 4347–4351. [Google Scholar] [CrossRef] [Green Version]
- Haber, J.E. Partners and pathways: Repairing a double-strand break. Trends Genet. 2000, 16, 259–264. [Google Scholar] [CrossRef]
- Khanna, K.K.; Jackson, S.P. DNA double-strand breaks: Signaling, repair and the cancer connection. Nat. Genet. 2001, 27, 247–254. [Google Scholar] [CrossRef]
- Koiri, R.K.; Mehrotra, A.; Trigun, S.K. Dalton’s lymphoma as a murine model for understanding the progression and development of t-cell lymphoma and its role in drug discovery. Int. J. Immunother. Cancer Res. 2017, 3, 001–006. [Google Scholar]
- Choudhury, B.; Kandimalla, R.; Bharali, R.; Monisha, J.; Kunnumakara, A.B.; Kalita, K.; Kotoky, J. Anticancer activity of Garcinia morella on T-cell murine lymphoma via apoptotic induction. Front. Pharmacol. 2016, 7, 3. [Google Scholar] [CrossRef] [PubMed]
- Shanker, A.; Singh, S.M.; Sodhi, A. Ascitic growth of a spontaneous transplantable T cell lymphoma induces thymic involution. Tumor Biol. 2000, 21, 315–327. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. No | Treatment | Viable Cell Count (105 Cells/mL) |
|---|---|---|
| 1 | DLA + Saline | 418.4 ± 19.7 |
| 2 | DLA + 4-PB | 10.35 ± 1.5 * |
| 3 | DLA + Artesunate | 5.82 ± 1.4 * |
| 4 | DLA + 4-PB + Artesunate | 2.47 ± 0.8 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Raza, A.; Kandimalla, R.; Kalita, S.; Ghosh, S.S. A Histone Deacetylase Inhibitor Manifests Synergistic Interaction with Artesunate by Suppressing DNA Repair Activity. Sci 2022, 4, 41. https://doi.org/10.3390/sci4040041
Raza A, Kandimalla R, Kalita S, Ghosh SS. A Histone Deacetylase Inhibitor Manifests Synergistic Interaction with Artesunate by Suppressing DNA Repair Activity. Sci. 2022; 4(4):41. https://doi.org/10.3390/sci4040041
Chicago/Turabian StyleRaza, Asif, Raghuram Kandimalla, Sanjeeb Kalita, and Siddhartha Sankar Ghosh. 2022. "A Histone Deacetylase Inhibitor Manifests Synergistic Interaction with Artesunate by Suppressing DNA Repair Activity" Sci 4, no. 4: 41. https://doi.org/10.3390/sci4040041