
The Effect of Food Deprivation on Foraging Behavior and Digestive and Metabolic Capacities of the Chinese Mitten Crab, Eriocheir sinensis
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
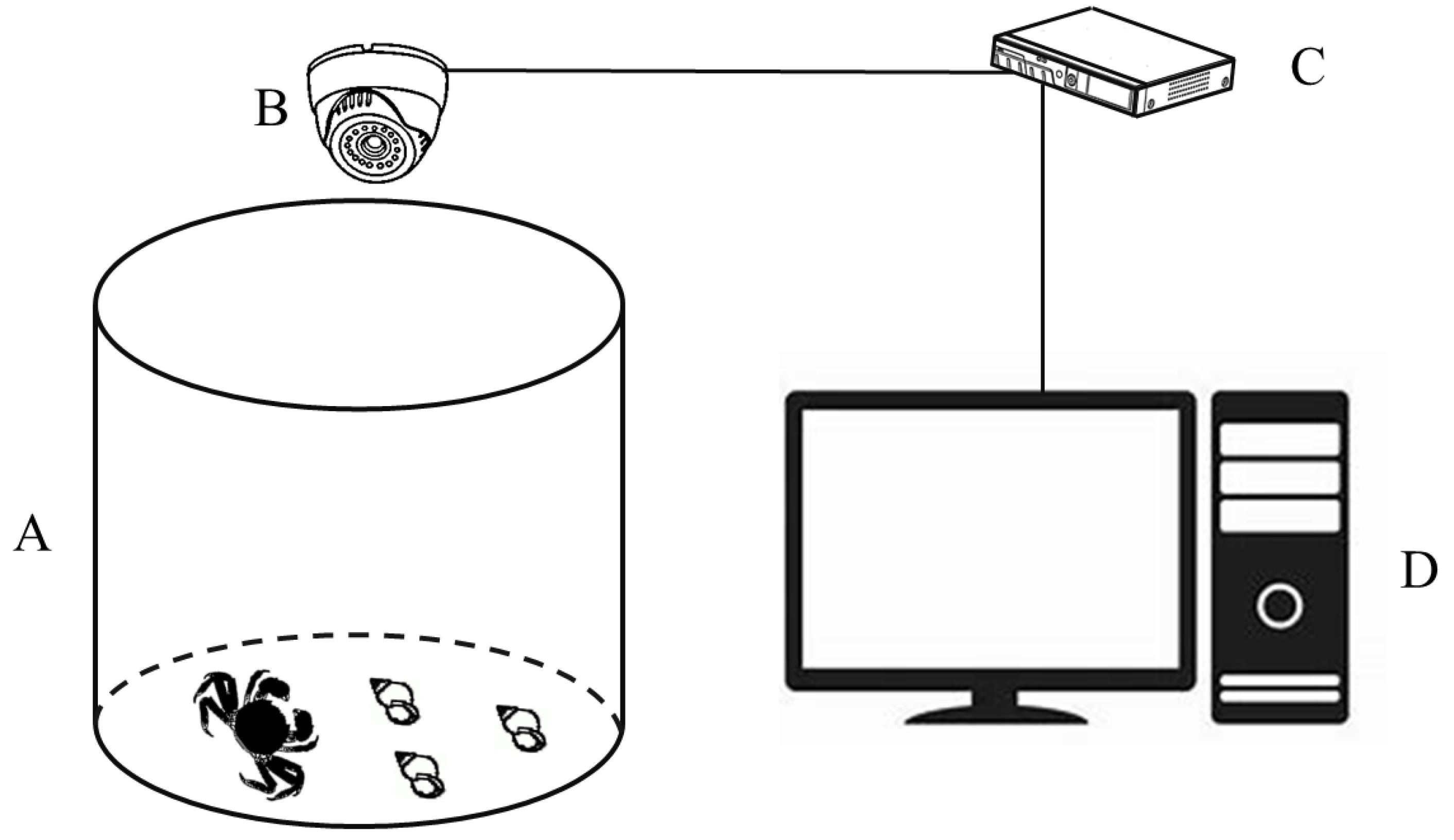
2.1. Resources and Temporary Culture of Experimental Crabs and Snails
2.2. Experimental Design and Procedures
2.3. Index and Determination Methods
2.3.1. Foraging Behavior Index
Pr (consumption/capture) = F3/F2
2.3.2. Shell-Breaking Techniques and the Proportion of Successful Snail Avoidance
2.3.3. Digestive and Metabolic Enzyme Activities
2.4. Statistical Analysis
3. Results
3.1. Average Predation Rate
3.2. Temporal Distribution of State Behavior
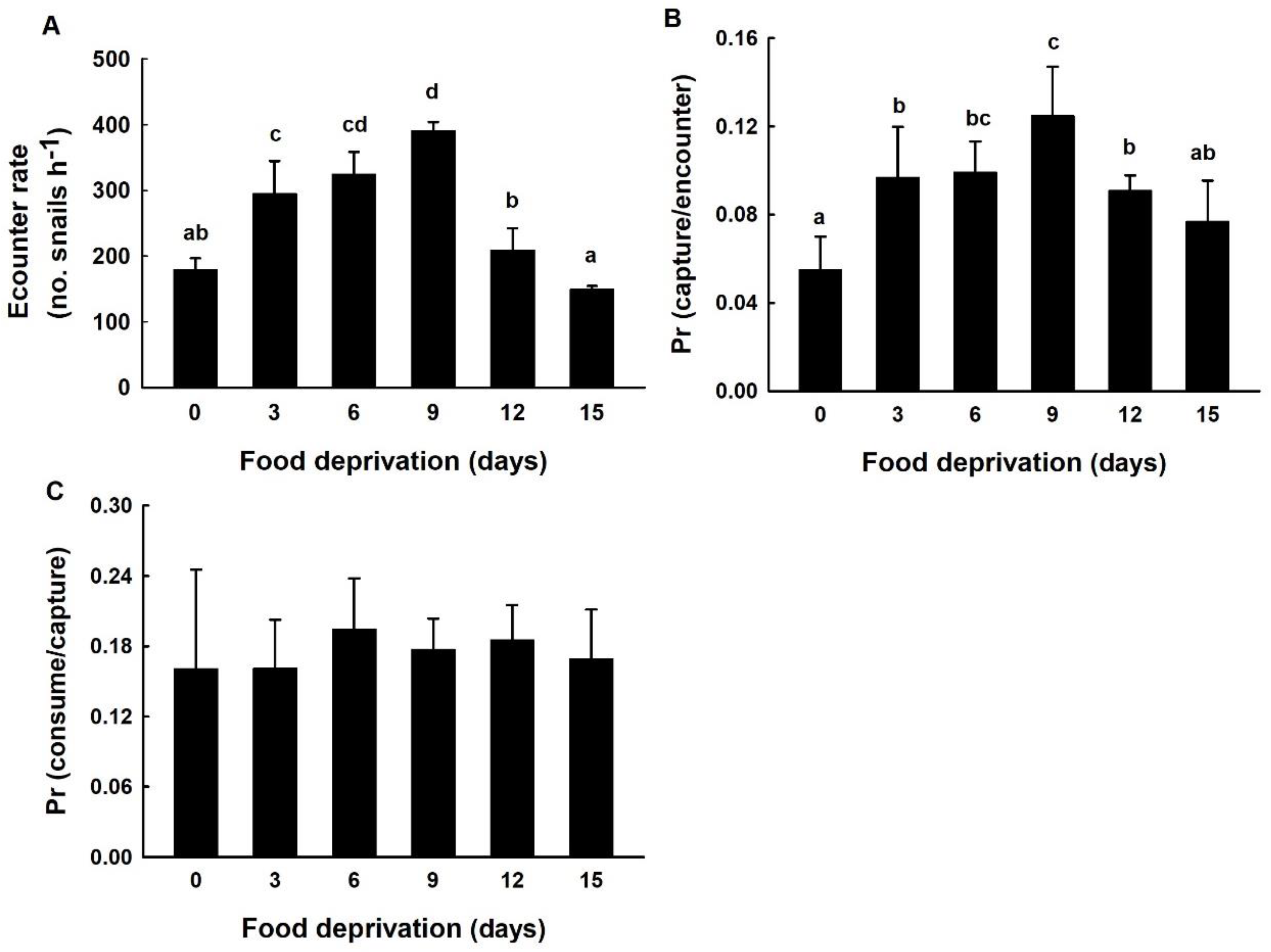
3.3. The Encounter Rate, the Probability of Capture upon Encounter, and the Probability of Consumption upon Capture
3.4. Handling Time per Snail, Proportion of Crushing Tactics, and Successful Avoidance
3.5. The Digestive and Metabolic Enzyme Activities
4. Discussion
4.1. Effects of Different Degrees of Food Deprivation on the Foraging Behavior of E. sinensis
4.2. Effects of Food Deprivation on the Shell-Breaking Techniques of E. sinensis
4.3. Effects of Food Deprivation on Crab Digestive and Metabolic Enzyme Activities
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- FDMA. China Fisheries Statistical Yearbook in 2020; China Agriculture Press: Beijing, China, 2020. [Google Scholar]
- Liu, J.; Sun, Y.; Han, W.; Cheng, F.; Chu, Y.; Yang, Z.; Cheng, Y. Effects of the size of Bellamya quadrata on the predation behavior of the Chinese mitten crab (Eriocheir sinensis). J. Shanghai Ocean Univ. 2022, 31, 140–150. [Google Scholar] [CrossRef]
- Wang, H.; Gin, M.; Shen, M.; Lu, C.; Shi, L.; Zhou, X.; Zhu, X. Effects of snails on yield and water quality of crab ponds and their suitable feeding amount. Fish. Sci. Technol. Inf. 2016, 43, 37–40. [Google Scholar] [CrossRef]
- Wang, H.; Shen, M.; Lu, C.; Shi, L.; Zhou, X.; Gin, M. Effect of replacement percentage of commercial feed by snails on yield and water quality in Chinese mitten handed crab culture. Fish. Sci. 2015, 34, 690–695. [Google Scholar] [CrossRef]
- Jin, G.; Xie, P.; Li, Z. Food habits of 2-year-old Chinese mitten crab, Eriocheir sinensis, stocked in lake bao’an. Acta Hydrobiol. Sin. 2003, 27, 140–146. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Geng, Z.; Feng, G.; Zhuang, P.; Wu, X.; Huang, X.; Shen, A.; Wang, H. Dietary and feeding intensity of female Eriocheir sinensis in the Yangtze River Estuary during migratory period. Mar. Fish. 2019, 41, 397–407. [Google Scholar] [CrossRef]
- Ramsay, K.; Kaiser, M.J.; Hughes, R.N. A field study of intraspecific competition for food in hermit crabs (Pagurus bernhardus). Estuar. Coast. Shelf Sci. 1997, 44, 213–220. [Google Scholar] [CrossRef]
- Zhou, S.; Shirley, T.C.; Kruse, G.H. Feeding and Growth of the Red King Crabs Paralithodes Camtschaticus Under Laboratory Conditions. J. Crustac. Biol. 1998, 18, 337–345. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Su, X.; Wang, F.; Zhong, D.; Sun, Y.; Zhang, D. Starvation intensifies the impacts of interspecific interactions on foraging behavior of swimming crab (Portunus trituberculatus). Aquaculture 2019, 504, 22–29. [Google Scholar] [CrossRef]
- Moir, F.; Weissburg, M.J. Cautious cannibals: Behavioral responses of juvenile and adult blue crabs to the odor of injured conspecifics. J. Exp. Mar. Biol. Ecol. 2009, 369, 87–92. [Google Scholar] [CrossRef]
- Curtis, D.L.; McGaw, I.J. Salinity and thermal preference of Dungeness crabs in the lab and in the field: Effects of food availability and starvation. J. Exp. Mar. Biol. Ecol. 2012, 413, 113–120. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, F.; Dong, S. A comparative study of the effect of starvation regimes on the foraging behavior of Portunus trituberculatus and Charybdis japonica. Physiol. Behav. 2015, 151, 168–177. [Google Scholar] [CrossRef]
- Liu, W.; Liu, B.; Liu, G.; Yin, X. Effects of Fasting Time on Food Selectivity of Water Fleas Daphnia magna. J. Dalian Ocean Univ. 2014, 29, 582–587. [Google Scholar] [CrossRef]
- Bardera, G.; Owen, M.A.G.; Pountney, D.; Alexander, M.E.; Sloman, K.A. The effect of short-term feed-deprivation and moult status on feeding behaviour of the Pacific white shrimp (Litopenaeus vannamei). Aquaculture 2019, 511, 734222. [Google Scholar] [CrossRef]
- Wang, X.; Wen, H.; Zhang, M.; Li, J.; Chen, S.; Zhang, K.; Liu, Y.; Tian, Y.; Chang, Z.; Wang, X. Cannibalism among Lateolabrax maculatus juveniles and its control under artificial culture conditions. Period. Ocean Univ. China 2019, 49, 16–22. [Google Scholar]
- Genodepa, J.G.; Zeng, C.; Southgate, P.C. Changes in digestive enzyme activities and nutrient utilization during embryonic development and starvation of newly hatched larvae of the mud crab, Scylla serrata. Aquaculture 2018, 493, 137–143. [Google Scholar] [CrossRef]
- Peter, M.; Lv, H.; Jiang, X.; Liu, Y.; Hur, J.-W.; Esbon, M.; Samwel, C.; Superius, E.; Khalfan, Z.; Gao, Y.; et al. Effects of starvation on enzyme activities and intestinal microflora composition in loach (Paramisgurnus dabryanus). Aquac. Rep. 2020, 18, 100467. [Google Scholar] [CrossRef]
- Li, Z.; Xie, S.; Wang, J.; Chen, D. Effects of intermittent starvation on growth and some digestive enzymes in the shrimp Macrobrachium nipponense. J. Fish. China 2007, 31, 42–48. [Google Scholar]
- Lu, G.; Liu, G.; Chen, X.; Li, J.; Wei, W.; Tang, J. Effects of starvation on digestive enzyme and antioxidant enzyme activities of Procambarus clarkii. J. Yangzhou Univ. Agric. Life Sci. Ed. 2012, 33, 33–38. [Google Scholar]
- Wang, Y.; Hu, M.; Wu, F.; Storch, D.; Pörtner, H.O. Elevated pCO2 affects feeding behavior and acute physiological response of the Brown crab Cancer pagurus. Front. Physiol. 2018, 9, 1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, M.C.; Barbeau, M.A. Effects of substrate on interactions between juvenile sea scallops (Placopecten magellanicus Gmelin) and predatory sea stars (Asterias vulgaris Verrill) and rock crabs (Cancer irroratus Say). J. Exp. Mar. Biol. Ecol. 2003, 287, 155–178. [Google Scholar] [CrossRef]
- Sun, Y.; Wang, F.; Su, X.; Liu, D.; Dong, S. Size matters for predator-prey relationships: The case of swimming crab Portunus trituberculatus preying on Manila clam Ruditapes philippinarum. Aquaculture 2017, 479, 638–644. [Google Scholar] [CrossRef]
- Barbeau, M.A.; Scheibling, R.E. Temperature effects on predation of juvenile sea scallops [Placopecten magellanicus (Gmelin)] by sea stars (Asterias vulgaris Verrill) and crabs (Cancer irroratus Say). J. Exp. Mar. Biol. Ecol. 1994, 182, 27–47. [Google Scholar] [CrossRef]
- Mills, C.D.; Clark, P.F.; Morritt, D. Flexible prey handling, preference and a novel capture technique in invasive, sub-adult Chinese mitten crabs. Hydrobiologia 2016, 773, 135–147. [Google Scholar] [CrossRef]
- Shigemiya, Y. Does the handedness of the pebble crab Eriphia smithii influence its attack success on two dextral snail species? J. Zool. 2003, 260, 259–265. [Google Scholar] [CrossRef]
- Alexander, J.E.; Covich, A.P. Predator avoidance by the freshwater snail Physella virgata in response to the crayfish Procambarus simulans. Oecologia 1991, 87, 435–442. [Google Scholar] [CrossRef]
- Wallace, J.C. Feeding, starvation and metabolic rate in the shore crab Carcinus maenas. Mar. Biol. 1973, 20, 277–281. [Google Scholar] [CrossRef]
- Comoglio, L.; Smolko, L.; Amin, O. Effects of starvation on oxygen consumption, ammonia excretion and biochemical composition of the hepatopancreas on adult males of the false southern king crab Paralomis granulosa (Crustacea, Decapoda). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2005, 140, 411–416. [Google Scholar] [CrossRef] [PubMed]
- Weisheng, H.; Chunyan, M.; Lingbo, M.; Fengying, Z.; Ming, Z.; Wei, W. Effects of starvation on molting and growth of the megalopae and first juvenile crab of Scylla paramamosain. Mar. Fish. 2022, 44, 54–66. [Google Scholar] [CrossRef]
- Xugan, W.; Rongbing, F.; Yongxu, C.; Silin, C.; Xiaozhen, Y.; Chulin, W.; Dongfa, Z.; Haizhong, L. Effects of starvation on the survival and the mainly biochemical composition of swimming crab (Portunus trituberculatus) freshly hatched larvae(Z1). Chin. J. Zool. 2006, 41, 7–13. [Google Scholar] [CrossRef]
- Jie, H.; Chun, W.; Luchang, S.; Yuhang, H.; Yi, L.; Yang, Z.; Yongxu, C. Impacts of starvation on the juveniles of Chinese mitten crab Eriocheir sinensis. Chin. J. Ecol. 2013, 32, 2077–2084. [Google Scholar]
- Lee, P.G.; Meyers, S.P. Chemoattraction and feeding stimulation in crustaceans. Aquacult. Nutr. 1996, 2, 157–164. [Google Scholar] [CrossRef]
- Micheli, F. Behavioural plasticity in prey-size selectivity of the blue crab Callinectes sapidus feeding on bivalve prey. J. Anim. Ecol. 1995, 64, 63–74. [Google Scholar] [CrossRef]
- Henan, Y.; Xiaotao, L.; Zhongneng, X.; Xiaozhuang, Z. Effects of starvation stress on behavior in Litopenaeus vannamei. J. Jinan Univ. (Nat. Sci.) 2010, 31, 100–104. [Google Scholar] [CrossRef]
- Weissburg, M.; Poulin, R.X.; Kubanek, J. You Are What You Eat: A Metabolomics Approach to Understanding Prey Responses to Diet-Dependent Chemical Cues Released by Predators. J. Chem. Ecol. 2016, 42, 1037–1046. [Google Scholar] [CrossRef] [PubMed]
- Schindler, D.E.; Johnson, B.M.; Mackay, N.A.; Bouwes, N.; Kitchell, J.F. Crab: Snail size-structured interactions and salt marsh predation gradients. Oecologia 1994, 97, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Bentley, M.G. The Global Spread of the Chinese Mitten Crab Eriocheir sinensis. In In the Wrong Place—Alien Marine Crustaceans: Distribution, Biology and Impacts; Springer: Dordrecht, The Netherlands, 2011; pp. 107–127. [Google Scholar]
- Calderwood, J.; O’Connor, N.E.; Roberts, D. Breaking and entering: Examining the role of stress and aerial exposure in predator–prey relationships between the common shore crab (Carcinus maenas) and cultivated blue mussels (Mytilus edulis). Aquaculture 2016, 452, 217–223. [Google Scholar] [CrossRef]
- Yu, X.; Zhang, X.; Yan, D.; Zhang, P.; Miao, Z. Effects of temperature, salinity, body length, and starvation on the critical swimming speed of whiteleg shrimp, Litopenaeus vannamei. Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2010, 157, 392–397. [Google Scholar] [CrossRef]
- Wang, Z.; Yang, Y. Survival and metamorphosis of the larvae Portunus trituberculatus treated by the short-period fasting. Acta Ocean. Sin. 2007, 29, 111–119. [Google Scholar] [CrossRef]
- Xu, D.; Wu, J.; Sun, L.; Qin, X.; Fan, X.; Zheng, X. Energy metabolism response of Litopenaeus vannamei to combined stress of acute cold exposure and waterless duration: Implications for physiological regulation and waterless live transport. J. Therm. Biol. 2022, 104, 103149. [Google Scholar] [CrossRef]
- Hu, M.; Wang, Y.; Tsang, S.T.; Cheung, S.G.; Shin, P.K.S. Effect of prolonged starvation on body weight and blood-chemistry in two horseshoe crab species: Tachypleus tridentatus and Carcinoscorpius rotundicauda (Chelicerata: Xiphosura). J. Exp. Mar. Biol. Ecol. 2010, 395, 112–119. [Google Scholar] [CrossRef]
- Zeng, L.; Li, F.; Li, X.; Cao, Z.; Fu, S.; Zhang, Y. The effects of starvation on digestive tract function and structure in juvenile southern catfish (Silurus meridionalis Chen). Comp. Biochem. Physiol. A Mol. Integr. Physiol. 2012, 162, 200–211. [Google Scholar] [CrossRef] [PubMed]
- Sacristan, H.J.; Ansaldo, M.; Franco-Tadic, L.M.; Gimenez, A.V.F.; Greco, L.S.L. Long-term starvation and posterior feeding effects on biochemical and physiological responses of midgut gland of Cherax quadricarinatus Juveniles (Parastacidae). PLoS ONE 2016, 11, e0150854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muhlia-Almazán, A.; García-Carreño, F. Influence of molting and starvation on the synthesis of proteolytic enzymes in the midgut gland of the white shrimp Penaeus vannamei. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2002, 133, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Luqing, P.; Hongyu, L.; Guoqiang, X. A review on digestive enzyme of crustacean larvae. J. Fish. Sci. China 2006, 13, 492–501. [Google Scholar] [CrossRef]
- Johnston, D.J.; Ritar, A.J.; Thomas, C.W. Digestive enzyme profiles reveal digestive capacity and potential energy sources in fed and starved spiny lobster (Jasus edwardsii) phyllosoma larvae. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2004, 138, 137–144. [Google Scholar] [CrossRef]
- Calvo, N.S.; Stumpf, L.; Sacristán, H.J.; Greco, L.S.L. Energetic reserves and digestive enzyme activities in juveniles of the red claw crayfish Cherax quadricarinatus nearby the point-of-no-return. Aquaculture 2013, 416–417, 85–91. [Google Scholar] [CrossRef]
- Wen, X.; Chen, L.; Ai, C.; Zhou, Z. Starvation metabolism in parent Chinese mitten-handed crab (Eriocheir sinensis). Chin. J. Appl. Ecol. 2002, 13, 1441–1444. [Google Scholar]
- Sánchez-Paz, A.; García-Carreño, F.; Muhlia-Almazán, A.; Peregrino-Uriarte, A.B.; Hernández-López, J.; Yepiz-Plascencia, G. Usage of energy reserves in crustaceans during starvation: Status and future directions. Insect Biochem. Mol. Biol. 2006, 36, 241–249. [Google Scholar] [CrossRef]
- Cuzon, G.; Cahu, C.; Aldrin, J.F.; Messager, J.L.; Stephan, G.; Mével, M. Starvation effect on metabolism of Penaeus japonicus. J. Proc. World Maricul. Soc. 2010, 11, 410–423. [Google Scholar] [CrossRef]
- Sacristán, H.J.; Nolasco-Soria, H.; Greco, L.S.L. Effect of attractant stimuli, starvation period and food availability on digestive enzymes in the redclaw crayfish Cherax quadricarinatus (Parastacidae). Aquat. Biol. 2014, 23, 87–99. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behavior | Description |
|---|---|
| Stationary | Crab is in a steady position or grooming. |
| Moving | Crab movement forward or backward with position change without probing/sweeping. |
| Searching | Crab’s walking leg probes and sweeps over the floor of the aquarium. |
| Handling | Handling for crabs is the period from capture until the crab moves away after consuming the snails. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Long, Q.; Liu, J.; Sun, Y.; Yang, Z.; Tang, B.; Cheng, Y. The Effect of Food Deprivation on Foraging Behavior and Digestive and Metabolic Capacities of the Chinese Mitten Crab, Eriocheir sinensis. Fishes 2023, 8, 47. https://doi.org/10.3390/fishes8010047
Long Q, Liu J, Sun Y, Yang Z, Tang B, Cheng Y. The Effect of Food Deprivation on Foraging Behavior and Digestive and Metabolic Capacities of the Chinese Mitten Crab, Eriocheir sinensis. Fishes. 2023; 8(1):47. https://doi.org/10.3390/fishes8010047
Chicago/Turabian StyleLong, Qiaolin, Jian Liu, Yunfei Sun, Zhigang Yang, Boping Tang, and Yongxu Cheng. 2023. "The Effect of Food Deprivation on Foraging Behavior and Digestive and Metabolic Capacities of the Chinese Mitten Crab, Eriocheir sinensis" Fishes 8, no. 1: 47. https://doi.org/10.3390/fishes8010047