
Effects of Water Temperature on the Growth, Antioxidant Capacity, and Gut Microbiota of Percocypris pingi Juveniles
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design and Sample Collection
2.2. Determination of Antioxidant Enzyme Activities in the Liver and Kidney
2.3. Gut Microbiota Structure Analyses
2.4. Data Analyses
3. Results
3.1. Effects of Water Temperature on the Growth of P. pingi
3.2. Effects of Water Temperature on the Activities of Three Antioxidant Enzymes in the Liver and Kidney of P. pingi
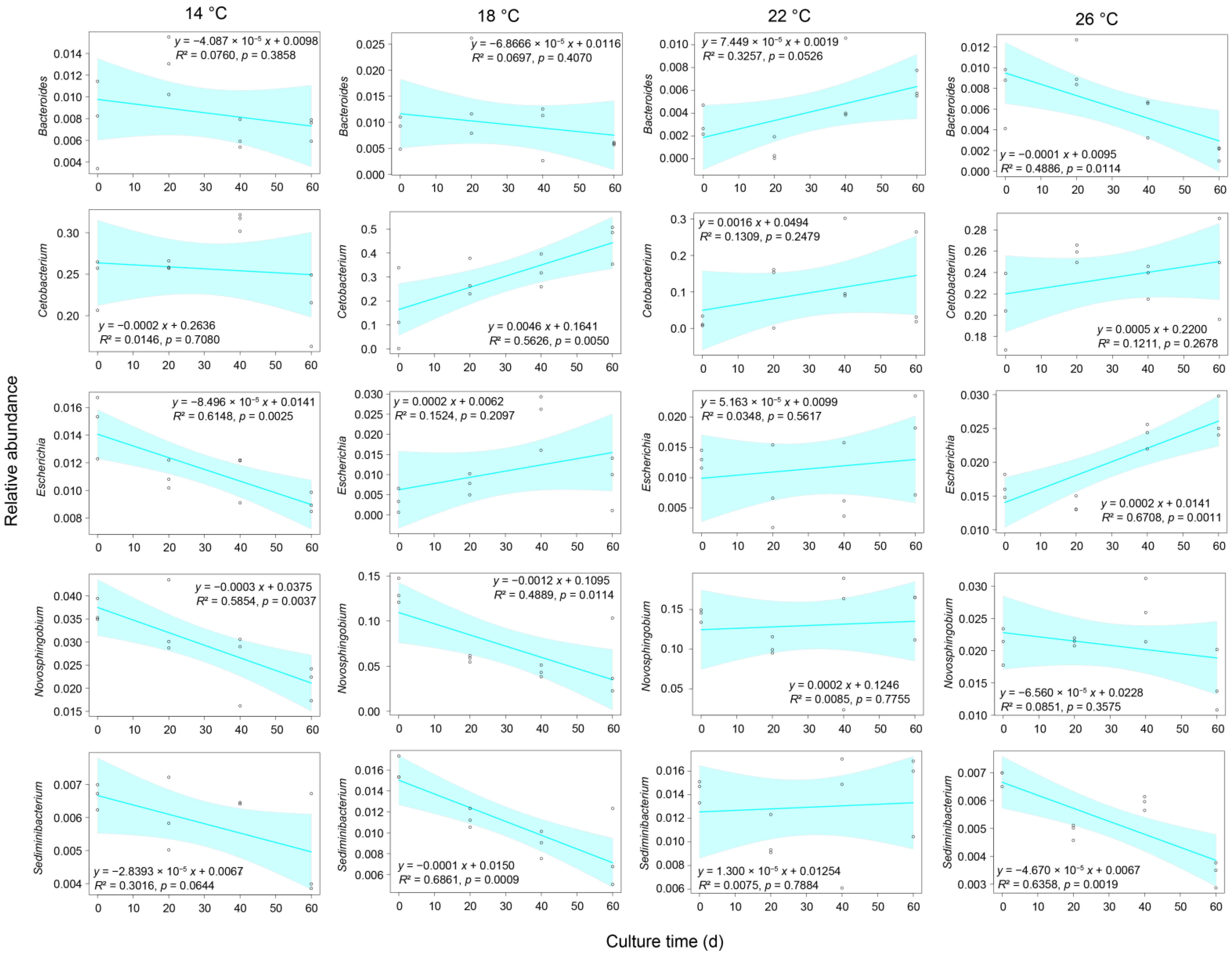
3.3. Effect of Water Temperature on the Gut Microbiota Structure of Percocypris pingi
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, Y.; Wang, J.; Peng, Z. The complete mitochondrial genome of Percocypris pingi (Teleostei, Cypriniformes). Mitochondrial. DNA 2013, 24, 40–42. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yue, X. Embryonic development of Percocypris pingi. Sichuan J. Zool. 2013, 32, 530–534. [Google Scholar] [CrossRef]
- He, Y.; Wu, X.; Zhu, Y.; Yang, D.; Zhan, H. Allometric growth pattern of Percocypris pingi pingi larvae. Chin. J. Zool. 2013, 48, 8–15. [Google Scholar]
- National Forestry and Grassland Administration. List of Key Protected Wild Animals in China. Available online: http://www.gov.cn/xinwen/2021-02/09/5586227/files/e007df5cdb364bcdbcb89d169047d6c5.pdf (accessed on 1 March 2021).
- Zhan, H.; Yang, D.; Li, Z.; Wang, Y.; Zhou, L.; Wu, X.; Yang, L. Artificial propagation of Percocypris pingi. J. Hydroecol. 2016, 37, 84–88. [Google Scholar] [CrossRef]
- Jin, F.-P.; Li, G.-H.; Li, L.; Qin, X.; Li, K.; Wu, J.-J.; Gao, H.-T.; Xia, Y.-D.; Liu, F. Effects of temperature, pH and salinity on the survival of juvenile Percocypris pingi retrodorslis. Acta Hydrobiol. Sinica 2018, 42, 578–583. [Google Scholar]
- Crawshaw, L.I. Physiological and behavioral reactions of fishes to temperature change. J. Fish. Res. Board. Can. 1977, 34, 730–734. [Google Scholar] [CrossRef]
- Volkoff, H.; Rønnestad, I. Effects of temperature on feeding and digestive processes in fish. Temperature 2020, 7, 307–320. [Google Scholar] [CrossRef]
- Neubauer, P.; Andersen, K.H. Thermal performance of fish is explained by an interplay between physiology, behaviour and ecology. Conserv. Physiol. 2019, 7, coz025. [Google Scholar] [CrossRef] [Green Version]
- Christensen, E.A.F.; Norin, T.; Tabak, I.; van Deurs, M.; Behrens, J.W. Effects of temperature on physiological performance and behavioral thermoregulation in an invasive fish, the round goby. J. Exp. Biol. 2021, 224, jeb237669. [Google Scholar] [CrossRef]
- Alfonso, S.; Gesto, M.; Sadoul, B. Temperature increase and its effects on fish stress physiology in the context of global warming. J. Fish. Biol. 2021, 98, 1496–1508. [Google Scholar] [CrossRef]
- Chen, X.; Deng, S.; Li, X.; Tan, Z.; Wan, Y.; Yao, L. Biological characteristics and culture techniques of Percocypris pingi. Hebei Fish. 2015, 254, 32–34. [Google Scholar] [CrossRef]
- Jandhyala, S.M.; Talukdar, R.; Subramanyam, C.; Vuyyuru, H.; Sasikala, M.; Reddy, D.N. Role of the normal gut microbiota. World J. Gastroenterol. 2015, 21, 8787–8803. [Google Scholar] [CrossRef]
- Jones, R.M. The influence of the gut microbiota on host physiology: In pursuit of mechanisms. Yale J. Biol. Med. 2016, 89, 285–297. [Google Scholar]
- Pickard, J.M.; Zeng, M.Y.; Caruso, R.; Núñez, G. Gut microbiota: Role in pathogen colonization, immune responses and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2018, 57, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Yang, Y.; Ishii, M.; Nagata, M.; Aw, W.; Obana, N.; Tomita, M.; Nomura, N.; Fukuda, S. Does the gut microbiota modulate host physiology through polymicrobial biofilms? Microbes Environ. 2020, 35, ME20037. [Google Scholar] [CrossRef]
- Ghosh, S.K.; Wong, M.K.-S.; Hyodo, S.; Goto, S.; Hamasaki, K. Temperature modulation alters the gut and skin microbial profiles of chum salmon (Oncorhynchus keta). Front. Marine Sci. 2022, 9, 1027621. [Google Scholar] [CrossRef]
- Bestion, E.; Jacob, S.; Zinger, L.; Gesu, L.D.; Richard, M.; White, J.; Cote, J. Climate warming reduces gut microbiota diversity in a vertebrate ectotherm. Nature Ecol. Evol. 2017, 1, 0161. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Zhang, B.; Liu, H.; Wang, J.; Wang, W.; Zhu, C.; Mu, Z.; Wang, J.; Zhou, J.; Wang, J. Effects of dietary protein level on growth, feed utilization, morphology parameters and muscle nutritional components of Schizopygopsis younghusbandi Regan. Chin. J. Anim. Nutr. 2019, 31, 1231–1239. [Google Scholar] [CrossRef]
- Gabryelak, T.; Zalesna, G.; Roche, H.; Pérès, G. Peroxide metabolism enzymes in erythrocytes of freshwater and marine fish species. Comp. Biochem. Physiol. 1989, 92, 5–8. [Google Scholar] [CrossRef]
- Popovic, N.T.; Strunjak-Perovic, I.; Coz-Rakovac, R.; Barisic, J.; Jadan, M.; Berakovic, A.P.; Klobucar, R.S. Tricaine methane-sulfonate (MS-222) application in fish anaesthesia. J. Appl. Ichthyol. 2012, 28, 553–564. [Google Scholar] [CrossRef] [Green Version]
- Xiang, J.; He, T.; Wang, P.; Xie, M.; Xiang, J.; Ni, J. Opportunistic pathogens are abundant in the gut of cultured giant spiny frog (Paa spinosa). Aquacul. Res. 2018, 49, 2033–2041. [Google Scholar] [CrossRef]
- Ni, J.; Huang, R.; Zhou, H.; Xu, X.; Li, Y.; Cao, P.; Zhong, K.; Ge, M.; Chen, X.; Hou, B.; et al. Analysis of the relationship between the degree of dysbiosis in gut microbiota and prognosis at different stages of primary hepatocellular carcinoma. Front. Microbiol. 2019, 10, 1458. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, J.; Fu, C.; Huang, R.; Li, Z.; Li, S.; Cao, P.; Zhong, K.; Ge, M.; Gao, Y. Metabolic syndrome cannot mask the changes of faecal microbiota compositions caused by primary hepatocellular carcinoma. Lett. Appl. Microbiol. 2021, 73, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. UPARSE: Highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Dixon, P. VEGAN, a package of R functions for community ecology. J. Veg. Sci. 2003, 14, 927–930. [Google Scholar] [CrossRef]
- Meyer, J.L.; Sale, M.J.; Mulholland, P.J.; Poff, N.L. Impacts of climate change on aquatic ecosystem functioning and health. J. Am. Water Resour. Assoc. 1999, 35, 1373–1386. [Google Scholar] [CrossRef]
- Daufresne, M.; Lengfellner, K.; Sommer, U. Global warming benefits the small in aquatic ecosystems. Proc. Natl. Acad. Sci. USA 2009, 106, 12788–12793. [Google Scholar] [CrossRef] [Green Version]
- Eissa, A.E.; Zaki, M.M. The impact of global climatic changes on the aquatic environment. Procedia Environ. Sci. 2011, 4, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Häder, D.-P.; Barnes, P.W. Comparing the impacts of climate change on the responses and linkages between terrestrial and aquatic ecosystems. Sci. Total. Environ. 2019, 682, 239–246. [Google Scholar] [CrossRef] [PubMed]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.-M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological responses to recent climate change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Root, T.L.; Price, J.T.; Hall, K.R.; Schneider, S.H.; Rosenzweig, C.; Pounds, J.A. Fingerprints of global warming on wild animals and plants. Nature 2003, 421, 57–60. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, C. About the relationships between heat conservation and body size of animals. Goett. Stud. 1847, 1, 595–708. [Google Scholar]
- Atkinson, D. Temperature and organism size: A biological law for ectotherms? Adv. Ecol. Res. 1994, 25, 1–58. [Google Scholar]
- Angilletta, M.J.; Dunham, A.E. The temperature-size rule in ectotherms: Simple evolutionary explanations may not be general. Am. Nat. 2003, 162, 332–342. [Google Scholar] [CrossRef] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The human microbiome project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.J.; Zhu, Q.G.; Yang, Z.; Zhao, N.; Ni, J.J. Cross seasonal inheritance and impact of ambient water microbiota on the gut microbiota of Rhinogobio cylindricus Günther. Appl. Ecol. Environ. Res. 2022, 20, 4539–4551. [Google Scholar] [CrossRef]
- Pörtner, H.O. Climate variations and the physiological basis of temperature dependent biogeography: Systemic to molecular hierarchy of thermal tolerance in animals. Comp. Biochem. Physiol. 2002, 132, 739–761. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Álvarez, R.M.; Morales, A.E.; Sanz, A. Antioxidant defenses in fish: Biotic and abiotic factors. Rev. Fish. Biol. Fish. 2005, 15, 75–88. [Google Scholar] [CrossRef]
- Yang, S.; Yan, T.; Zhao, L.; Wu, H.; Du, Z.; Yan, T.; Xiao, Q. Effects of temperature on activities of antioxidant enzymes and Na+/K+—ATPase, and hormone levels in Schizothorax prenanti. J. Thermal. Biol. 2018, 72, 155–160. [Google Scholar] [CrossRef] [PubMed]
- Luo, H.; Li, Q.; Li, C. The impact of water temperature during the fish reproduction in the upper Yangtze River due to the cascade development in the lower Jinsha River. J. China Inst. Water Resour. Hydropow. Res. 2012, 10, 256–259. [Google Scholar]
- Li, Q.; Li, C.; Luo, H. Ecological target water temperature of rare and endemic fish in Yangtze River upstream. J. China Inst. Water Resour. Hydropow. Res. 2012, 10, 86–91. [Google Scholar]
- Sun, D.; Tian, H.; Zhang, H.; Zhou, Z.; Zhou, H. Monitoring of water temperature and changing relationship between the water temperature and air temperature in the upper Yangtze River. J. Build. Energ. Effic. 2010, 12, 74–77. [Google Scholar] [CrossRef]
- Kalita, A.; Hu, J.; Torres, A.G. Recent advances in adherence and invasion of pathogenic Escherichia coli. Curr. Opin. Infect. Dis. 2014, 27, 459–464. [Google Scholar] [CrossRef]
- Poolman, J.T.; Wacker, M. Extraintestinal pathogenic Escherichia coli, a common human pathogen: Challenges for vaccine development and progress in the field. J. Infect. Dis. 2016, 213, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Dikshit, R.P.; Dikshit, K.L.; Liu, Y.; Webster, D.A. The bacterial hemoglobin from Vitreoscilla can support the aerobic growth of Escherichia coli lacking terminal oxidases. Arch. Biochem. Biophys. 1992, 293, 241–245. [Google Scholar] [CrossRef]
- Deng, H.; Li, Z.; Tan, Y.; Guo, Z.; Liu, Y.; Wang, Y.; Yuan, Y.; Yang, R.; Bi, Y.; Bai, Y.; et al. A novel strain of Bacteroides fragilis enhances phagocytosis and polarises M1 macrophages. Sci. Rep. 2016, 6, 29401. [Google Scholar] [CrossRef] [Green Version]
- Tan, H.; Zhai, Q.; Chen, W. Investigations of Bacteroides spp. towards next-generation probiotics. Food Res. Int. 2019, 116, 637–644. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Item | Culture Time (d) | Culture Temperature (°C) | |||
|---|---|---|---|---|---|
| 14 | 18 | 22 | 26 | ||
| Body weight (g/ind.) | 0 | 33.61 ± 0.77 | 34.65 ± 0.45 | 34.43 ± 0.97 | 33.39 ± 0.93 |
| 20 | 35.95 ± 0.64 a | 40.84 ± 0.97 b | 40.34 ± 0.68 b | 36.11 ± 1.00 a | |
| 40 | 39.94 ± 1.21 a | 48.33 ± 1.17 b | 46.84 ± 1.12 b | 37.42 ± 1.16 a | |
| 60 | 42.58 ± 0.48 a | 53.25 ± 1.41 b | 52.79 ± 1.09 b | 40.02 ± 1.36 a | |
| Weight gain rate (%) | 20 | 6.97 ± 0.55 a | 17.87 ± 3.02 b | 17.22 ± 2.26 b | 8.19 ± 4.05 a |
| 40 | 11.08 ± 1.39 a | 18.34 ± 0.74 b | 16.13 ± 1.40 b | 3.62 ± 0.35 c | |
| 60 | 6.66 ± 2.24 a | 10.17 ± 0.76 ab | 12.7 ± 2.79 b | 6.96 ± 1.96 a | |
| Specific growth rate (%) | 20 | 0.34 ± 0.03 a | 0.82 ± 0.13 b | 0.79 ± 0.10 b | 0.39 ± 0.19 a |
| 40 | 0.53 ± 0.07 a | 0.84 ± 0.03 b | 0.75 ± 0.06 b | 0.18 ± 0.02 c | |
| 60 | 0.32 ± 0.11 a | 0.48 ± 0.03 a,b | 0.60 ± 0.13 b | 0.33 ± 0.09 a | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.; Li, X.; Zhu, Y.; Gong, J.; Zhu, T.; Ni, J.; Yang, D. Effects of Water Temperature on the Growth, Antioxidant Capacity, and Gut Microbiota of Percocypris pingi Juveniles. Fishes 2022, 7, 374. https://doi.org/10.3390/fishes7060374
Wu X, Li X, Zhu Y, Gong J, Zhu T, Ni J, Yang D. Effects of Water Temperature on the Growth, Antioxidant Capacity, and Gut Microbiota of Percocypris pingi Juveniles. Fishes. 2022; 7(6):374. https://doi.org/10.3390/fishes7060374
Chicago/Turabian StyleWu, Xingbing, Xuemei Li, Yongjiu Zhu, Jinling Gong, Tingbing Zhu, Jiajia Ni, and Deguo Yang. 2022. "Effects of Water Temperature on the Growth, Antioxidant Capacity, and Gut Microbiota of Percocypris pingi Juveniles" Fishes 7, no. 6: 374. https://doi.org/10.3390/fishes7060374