


Proteomic Analysis Associated with the Immune Response in Hemocytes of Portunus trituberculatus Challenged with Vibrio parahaemolyticus
Abstract
:1. Introduction
2. Material and Methods
2.1. Statement of Ethics
2.2. Experimental Animals and Sample Collection
2.3. Protein Preparation
2.4. iTRAQ Labeling and Reversed-Phase (RP) Fractionation
2.5. Liquid Chromatography–Electrospray Ionization–Tandem Mass Spectrometry (LC-ESI-MS/MS)
2.6. Data Analyses
2.7. Quantitative Real-Time PCR (qRT-PCR)
3. Results
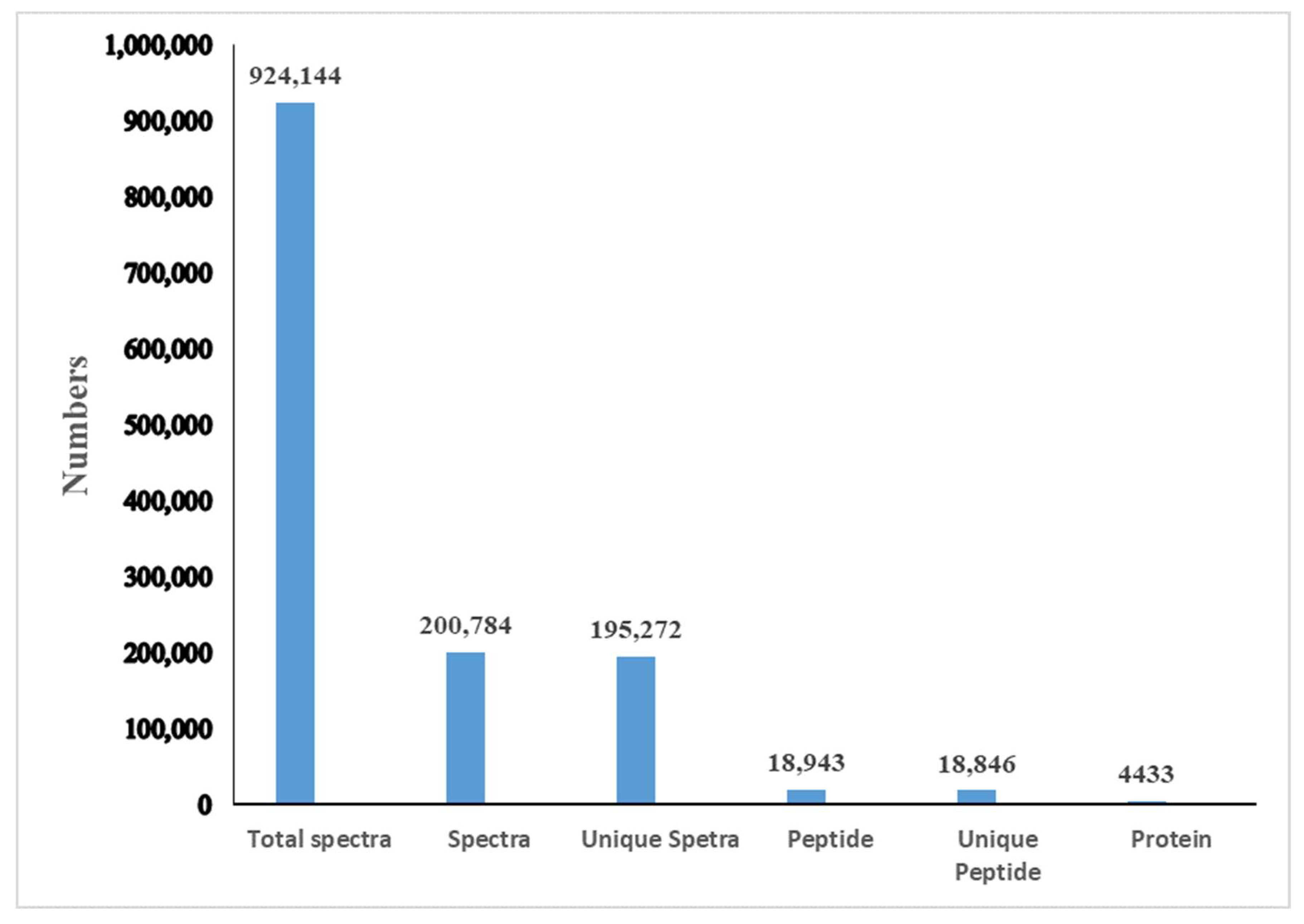
3.1. Protein Profiling
3.2. iTRAQ Quantification
3.3. GO Enrichment Analysis
3.4. Immune-Related KEGG Pathways
3.5. Validation of Proteomic Data by qRT-PCR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fishery Bureau, Ministry of Agriculture, China. China Fisheries Yearbook 2022; Chinese Agriculture Express: Being, China, 2022; Volume 22. (In Chinese)
- Wang, G.L.; Shan, J.; Chen, Y.; Li, Z. Study on pathogens and pathogenesis of emulsification disease of Portunus trituberculatus. Adv. Mar. Sci. 2006, 24, 527–531. (In Chinese) [Google Scholar]
- Zhou, J.; Li, X.; Wang, J.; Wang, Y.; Wan, X.; Fang, W. Analysis of major infectious diseases and their causative agents of Chinese sea crabs. Mar. Sci. 2014, 38, 102–106. (In Chinese) [Google Scholar]
- Yan, B.L.; Qin, G.M.; Bao, Z.H.; Zhang, X.J.; Bi, K.R.; Qin, L. Isolation and identification of Vibrio parahaemolyticus from diseased Portunus trituberculatus. Mar. Sci. Bull. 2010, 29, 560–566. (In Chinese) [Google Scholar]
- Xiao, G.H.; Zhao, C.L.; Cui, Z.J. Main Reasons and Control Methods of Artificial Seedling Raising and Disease of Portunus trituberculatus. Shandong Fish. 2006, 23, 36–37. (In Chinese) [Google Scholar]
- Amparyup, P.; Charoensapsri, W.A. Tassanakajon, Prophenoloxidase system and its role in shrimp immune responses against major pathogens. Fish Shellfish. Immunol. 2013, 34, 990–1001. [Google Scholar] [CrossRef] [PubMed]
- Cerenius, L.; Lee, B.L.; Soderhall, K. The proPO-system: Pros and cons for its role in invertebrate immunity. Trends Immunol. 2008, 29, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lü, J.J.; Liu, P.; Li, J.; Wang, Z.Q.; Zhang, X.H. Cloning of Toll4 in Portunus trituberculatus and its expression in responding to pathogenic infection and low salinity stress. Prog. Fish. Sci. 2018, 39, 146–155. (In Chinese) [Google Scholar]
- Robalino, J.; Carnegie, R.B.; O‘Leary, N.; Ouvry-Patat, S.A.; de la Vega, E.; Prior, S.; Gross, P.S.; Browdy, C.L.; Chapman, R.W.; Schey, K.L.; et al. Contributions of functional genomics and proteomics to the study of immune responses in the Pacific white leg shrimp Litopenaeus vannamei. Vet. Immunol. Immunopathol. 2009, 128, 110–118. [Google Scholar] [CrossRef]
- Leu, J.-H.; Chang, C.-C.; Wu, J.-L.; Hsu, C.-W.; Hirono, I.; Aoki, T.; Juan, H.-F.; Lo, C.-F.; Kou, G.-H.; Huang, H.-C. Comparative analysis of differentially expressed genes in normal and white spot syndrome virus infected Penaeus monodon. BMC Genom. 2007, 8, 120. [Google Scholar] [CrossRef] [PubMed]
- Rojtinnakorn, J.; Hirono, I.; Itami, T.; Takahashi, Y.; Aoki, T. Gene expression in haemocytes of kuruma prawn, Penaeus japonicus, in response to infection with WSSV by EST approach. Fish. Shellfish Immunol. 2002, 13, 69–83. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.B.; Shi, J.Y.; Hao, W.J.; Xiang, T.; Zhou, H.F.; Wang, W.; Meng, Q.; Ding, Z. iTRAQ-based quantitative proteomic analysis of Procambarus clakii hemocytes during Spiroplasma eriocheiris infection. Fish. Shellfish Immunol. 2017, 7, 438–444. [Google Scholar] [CrossRef] [PubMed]
- Anderson, N.L.; Anderson, N.G. Proteome and proteomics: New technologies, new concepts, and new words. Electrophoresis 1998, 19, 1853–1861. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.B.; Xiu, Y.J.; Wang, J.; Liu, X.Q.; Liu, Y.H.; Gua, W.; Wang, W.; Meng, Q.G. iTRAQ-based quantitative proteomic analysis of Macrobrachium rosenbergii hemocytes during Spiroplasma eriocheiris infection. J. Proteom. 2016, 136, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Sun, B.; Wang, Z.; Wang, Z.; Ma, X.; Zhu, F. A proteomic study of hemocyte Proteins from Mud crab (Scylla paramamosain) infected with White spot syndrome Virus or Vibrio alginolyticus. Front. Immunol. 2017, 8, 468. [Google Scholar] [CrossRef] [PubMed]
- Pierce, A.; Unwin, R.D.; Evans, C.A.; Griffiths, S.; Carney, L.; Zhang, L. Eightchannel iTRAQ enables comparison of the activity of six leukemogenic tyrosine kinases. Mol. Cell. Proteom. 2008, 7, 853–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, Q.; Hou, L.; Zhao, Y.; Huang, X.; Huang, Y.; Xia, S.; Gu, W.; Wang, W. iTRAQ based proteomic study of the effects of Spiroplasma eriocheiris on Chinese mitten crab Eriocheir sinensis hemocytes. Fish. Shellfish Immunol. 2014, 40, 182–189. [Google Scholar] [CrossRef]
- Jeswin, J.; Xie, X.L.; Ji, Q.L.; Wang, K.J.; Liu, H.P. Proteomic analysis by iTRAQ in red claw crayfish, Cherax quadricarinatus, hematopoietic tissue cells post white spot syndrome virus infection. Fish. Shellfish Immunol. 2016, 50, 288–296. [Google Scholar] [CrossRef] [PubMed]
- Saray, P.; Roytrakul, S.; Pangeson, T.; Phetrungnapha, A. Comparative proteomic analysis of hepatopancreas in Macrobrachium rosenbergii responded to Poly (I:C). Fish. Shellfish Immunol. 2018, 75, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Lü, J.J.; Liu, P.; Li, J.; Wang, Z.Q.; Zhang, X.H. Cloning of HMGBa in Portunus trituberculatus and its expression in responding to pathogenic infection. Acta Hydrobiol. Sin. 2017, 41, 1193–1199. (In Chinese) [Google Scholar]
- Song, L.; Lv, J.J.; Wang, L.; Sun, D.F.; Gao, B.Q.; Liu, P. Characterization of a chitinase-1 gene (PtCht-1) from a marine crab Portunus trituberculatus and its response to immune stress. Gene 2020, 741, 144523. [Google Scholar] [CrossRef]
- Gao, B.Q.; Sun, D.F.; Lv, J.J.; Ren, X.Y.; Liu, P.; Li, J. Transcriptomic analysis provides insight into the mechanism of salinity adjustment in swimming crab Portunus trituberculatus. Genes Genom. 2019, 41, 961–971. [Google Scholar] [CrossRef] [PubMed]
- Li, S.K.; Sun, L.B.; Wu, H.J.; Hu, Z.; Liu, W.H.; Li, Y.Y.; Wen, X. The intestinal microbial diversity in mud crab (Scylla paramamosain) as determined by PCR-DGGE and clone library analysis. J. Appl. Microbiol. 2012, 113, 1341–1351. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.Y.; Xia, X.A.; Wu, Q.Y.; Liu, W.H.; Lin, Y.S. Infection with Hematodinium sp. in mud crabs Scylla serrata cultured in low salinity water in southern China. Dis. Aquat. Org. 2008, 82, 145–150. [Google Scholar] [CrossRef]
- Yang, Y.T.; Chen, I.T.; Lee, C.T.; Chen, C.Y.; Lin, S.S.; Hor, L.I. Draft genome sequences of four strains of Vibrio parahaemolyticus, three of which cause early mortality syndrome/acute hepatopancreatic necrosis disease in shrimp in China and Thailand. Genome Announc. 2014, 2, e00816-14. [Google Scholar] [CrossRef] [PubMed]
- Song, L. Functional identification of chitinase gene family and its immune mechanism in response to salinity changes in Portunus trituberculatus. Master’s Thesis, Shanghai Ocean University, Shanghai, China, 2019; p. 50. (In Chinese). [Google Scholar]
- Zhang, Y.L.; Wang, S.Y.; Xu, A.L.; Chen, J.; Lin, B.K.; Peng, X.X. Affinity proteomic approach for identification of an IgA-like protein in Litopenaeus vannamei and study on its agglutination characterization. J. Proteome Res. 2006, 5, 815–821. [Google Scholar] [CrossRef]
- Bachea, E.; Mialhe, E.; Noel, D.; Bouloa, V.; Morvana, A.; Rodriguezc, J. Knowledge and research prospects in marine mollusc and crustacean immunology. Aquaculture 1995, 132, 17–32. [Google Scholar] [CrossRef]
- Zhao, X.; Guo, L.; Lu, X.; Lu, H.; Wang, F.; Zhong, M.; Chen, J.; Zhang, Y. Evidences of abundant hemocyanin variants in shrimp Litopenaeus vannamei. Mol. Immunol. 2016, 77, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Huang, C.; Qin, Q. Antiviral properties of hemocyanin isolated from shrimp Penaeus monodon. Antivir. Res. 2004, 61, 93–99. [Google Scholar] [CrossRef]
- Dhar, A.K.; Dettori, A.; Roux, M.M.; Klimpel, K.R.; Read, B. Identification of differentially expressed genes in shrimp (Penaeus stylirostris) infected with white spot syndrome virus by cDNA microarrays. Arch. Virol. 2003, 148, 2381–2396. [Google Scholar] [CrossRef]
- Gross, P.S.; Bartlett, T.C.; Browdy, C.L.; Chapman, R.W.; Warr, G.W. Immune gene discovery by expressed sequence tag analysis of hemocytes and hepatopancreas in the Pacific white shrimp, Litopenaeus vannamei, and the Atlantic white shrimp, L-setiferus. Dev. Comp. Immunol. 2001, 25, 565–577. [Google Scholar] [CrossRef]
- Wu, J.; Lin, Q.; Lim, T.K.; Liu, T.; Hew, C. White spot syndrome virus proteins and differentially expressed host proteins identified in shrimp epithelium by shotgun proteomics and cleavable isotope-coded affinity tag. J. Virol. 2007, 81, 11681–11689. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Luan, S.; Dai, P.; Meng, X.H.; Cao, B.X.; Luo, K.; Kong, J. iTRAQ-based comparative proteome analysis for molecular mechanism of defense against acute ammonia toxicity in Pacific White shrimp Litopenaeus vannamei. Fish. Shellfish Immunol. 2018, 74, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Jiang, S.; Huang, J.; Zhou, F.; Yang, Q.; Jiang, S.; Yang, L. C-type lectin response to bacterial infection and ammonia nitrogen stress in tiger shrimp (Penaeus monodon). Fish. Shellfish Immunol. 2019, 90, 188–198. [Google Scholar] [CrossRef]
- Cerenius, L.; Kawabata, S.I.; Lee, B.L.; Nonaka, M.; Söderhäll, K. Proteolytic cascades and their involvement in invertebrate immunity. Trends Biochemical. Sci. 2010, 35, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.M.; Wang, B.; Zhang, J.Q.; Li, F.H.; Xiang, J.H. Multiple forms of alpha-2 macroglobulin in shrimp Fenneropenaeus chinesis and their transcriptional response to WSSV or Vibrio pathogen infection. Dev. Comp. Immunol. 2010, 34, 677–684. [Google Scholar] [CrossRef]
- Li, M.; Wang, J.; Huang, Q.; Li, C. Proteomic analysis highlights the immune responses of the hepatopancreas against Hematodinium infection in Portunus trituberculatus. J. Proteom. 2019, 197, 92–105. [Google Scholar] [CrossRef]
- Li, F.; Xiang, J. Recent advances in researches on the innate immunity of shrimp in China. Dev. Comp. Immunol. 2013, 39, 11–26. [Google Scholar] [CrossRef]
- Li, M.; Li, C.; Wang, J.; Song, S. Molecular characterization and expression of a novel Toll gene from the swimming crab Portunus trituberculatus. Mol. Immunol. 2015, 67, 388–397. [Google Scholar] [CrossRef]
- Yin, F.; Gao, Q.; Tang, B.; Sun, P.; Han, K.; Huang, W. Transcriptome and analysis on the complement and coagulation cascades pathway of large yellow croaker (Larimichthys crocea) to ciliate ectoparasite Cryptocaryon irritans infection. Fish. Shellfish Immunol. 2016, 50, 127–141. [Google Scholar] [CrossRef]
- Larvie, M.; Takahashi, K. Fluid phase barrier immunity and serine protease cascades–the essential roles of the coagulation system. J. Hematol. Thromboembolic Dis. 2014, 2, 1000158. [Google Scholar]
- Ponprateep, S.; Vatanavicharn, T.; Lo, C.F.; Tassanakajon, A.; Rimphanitchayakit, V. Alpha-2-macroglobulin is a modulator of prophenoloxidase system in pacific white shrimp Litopenaeus vannamai. Fish. Shellfish Immunol. 2017, 62, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Prota, A.E.; Campbell, J.A.; Schelling, P.; Forrest, J.C.; Watson, M.J.; Peters, T.R.; Aurrand-Lions, M.; Imhof, B.A.; Dermody, T.S.; Stehle, T. Crystal structure of human junctional adhesion molecule 1: Implications for reovirus binding. Proc. Natl. Acad. Sci. USA 2003, 100, 5366–5371. [Google Scholar] [CrossRef] [PubMed]
- Kostrewa, D.; Brockhaus, M.; D’Arcy, A.; Dale, G.E.; Nelboeck, P.; Schmid, G.; Mueller, F.; Bazzoni, G.; Dejana, E.; Bartfai, T.; et al. X-ray structure of junctional adhesion molecule: Structural basis for hemophilic adhesion via a novel dimerization motif. EMBO J. 2001, 20, 4391–4398. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward (F)/Reverse (R) Primers | Sequences from 5′ to 3′ |
|---|---|
| Ribosomal protein L10-qRT-F | GGTGAAGAACTGCGGCAAGGA |
| Ribosomal protein L10-qRT-R | GCATCCCCGTTTGGAGCCTAT |
| Scavenger receptor class B-qRT-F | AATGGAGCCAAGCCCGACCT |
| Scavenger receptor class B-qRT-R | TCGTCCCAAACCCACCACCT |
| Hemocyanin subunit 3-qRT-F | GCCAAACAGGTTCCTCATTCCC |
| Hemocyanin subunit 3-qRT-R | GCAACCCTTCAACGGCAGCA |
| C type lectin containing domain protein-qRT-F | AGAGGCGAACAATGGCGAGTG |
| C type lectin containing domain protein-qRT-R | TTAGACAGCGGTGACGCAGAGG |
| Clip domain serine proteinase 2-qRT-F | GCTGGACCACACTGAAGGGGAT |
| Clip domain serine proteinase 2-qRT-R | ATGCCACATCTGGGAGGCTGCT |
| Serine protease-qRT-F | TAACCTGCTGCTGAGTGCCTACC |
| Serine protease-qRT-R | TCCACCCTCGTCACCATTACAAG |
| β-actin-F | CGAAACCTTCAACACTCCCG |
| β-actin-R | GGGACAGTGTGTGAAACGCC |
| KEGG_ID | Protein_ID | Description | Ratio (36 h vs. Control) | Ratio (72 h vs. Control) |
|---|---|---|---|---|
| Cell adhesion molecules (CAMs) | comp38016_c0 comp42008_c1 comp44017_c0 comp56776_c0 comp54701_c0 comp52938_c0 comp23347_c0 comp62976_c0 comp48402_c0 comp132136_c0 comp43324_c0 comp463272_c0 comp53019_c0 comp39439_c0 comp47792_c0 comp16281_c0 comp41102_c0 comp44493_c0 | Tyrosine-protein phosphatase 10D Nidogen-1 E2C1D1_HARSA Teneurin-3 Type-2 ice-structuring protein Q9GV46_9EUCA oxygenase Laminin subunit beta-1 Techylectin-5B Transforming growth factor–beta-induced protein ig-h3 Integrin alpha-8 Ficolin-1 Plexin-A1 Motile sperm domain-containing protein 2 Laminin subunit gamma-1 MAM and LDL-receptor class A domain-containing protein C10orf112 Ryncolin-2 Basement membrane-specific heparan sulfate proteoglycan core protein von Willebrand factor D and EGF domain-containing fasciclin-2 | 0.68 0.57 1.28 1.68 2.23 0.77 0.60 0.58 0.83 1.78 1.35 0.70 0.74 0.80 0.98 0.91 0.98 0.88 | 0.71 0.49 1.04 1.15 0.45 0.60 0.48 0.53 0.84 1.36 1.22 0.51 0.56 0.82 1.21 0.60 0.83 0.76 |
| Complement and coagulation cascades | comp41102_c0 comp23347_c0 comp48863_c0 comp49374_c1 comp39018_c0 | von Willebrand factor D and EGF domain-containing protein Techylectin-5B Alpha-2-macroglobulin Alpha-2-macroglobulin-like protein 1 Serine proteinase stubble | 0.98 0.60 1.54 1.22 1.27 | 0.83 0.48 1.31 1.25 1.33 |
| Phagosomes | comp53019_c0 comp50028_c0 comp57849_c0 comp42008_c0 comp47792_c0 comp26865_c0 comp51455_c0 comp16281_c0 comp41102_c0 comp50625_c0 comp26085_c0 comp32356_c0 comp52938_c0 comp23347_c0 comp62976_c0 comp50837_c0 comp55957_c0 comp49730_c0 comp44017_c0 comp56776_c0 comp48402_c0 comp30556_c0 | Laminin subunit gamma-1 Early endosome antigen 1 Tubulin alpha chain Nidogen-1 Ryncolin-2 Vesicle-trafficking protein SEC22b-B Cytoplasmic dynein 1 light intermediate chain 2 Basement membrane-specific heparan sulfate proteoglycan core protein von Willebrand factor D and EGF domain-containing protein Cdc42 homolog Tubulin alpha-3 chain Pulmonary surfactant-associated protein D Laminin subunit beta-1 Techylectin-5B Transforming growth factor–beta-induced protein ig-h3 Protein croquemort Tubulin alpha-2 chain Tubulin alpha chain E2C1D1_HARSA teneurin-3 C-type lectin protein Integrin alpha-8 Syntaxin-12 | 0.74 0.88 0.84 0.57 0.98 0.78 0.78 0.91 0.98 0.62 1.11 1.17 0.77 0.60 0.58 0.71 0.83 0.80 1.28 1.68 0.83 0.82 | 0.56 0.84 0.79 0.49 1.21 0.75 0.75 0.60 0.83 0.76 1.22 1.73 0.60 0.48 0.53 0.83 0.95 0.88 1.04 1.15 0.84 0.86 |
| Lysosomes | comp30813_c0 comp49562_c0 comp4076_c0 comp31475_c0 comp48159_c0 comp170042_c0 comp40157_c0 comp30098_c0 comp60626_c0 comp109451_c0 | AP-3 complex subunit delta-1 ADP-ribosylation factor-binding protein GGA1 AP-3 complex subunit delta-1 AP-1 complex subunit gamma-1 AP-3 complex subunit beta-2 Chitooligosaccharidolytic beta-N-acetylglucosaminidase AP-1 complex subunit sigma-2 E9GTP3_DAPPU putative uncharacterized protein AP-1 complex subunit beta-1 ADP-ribosylation factor-binding protein GGA3 | 0.86 0.77 0.88 0.83 0.87 1.54 0.76 0.82 0.81 0.73 | 0.79 0.77 0.77 0.87 0.83 1.35 0.88 0.84 0.80 0.79 |
| Tight junctions | comp53821_c0 comp43480_c0 comp21124_c0 comp17686_c0 comp434052_c0 comp36045_c0 comp54789_c0 comp283514_c0 comp37064_c0 comp54333_c0 | Spectrin beta chain Lethal(2) giant larvae protein homolog 2 FACT complex subunit spt16 Exocyst complex component 3 Myosin regulatory light chain 2 MICAL-like protein 2 Myosin heavy chain MAGUK p55 subfamily member 7 Symplekin Myosin heavy chain | 0.86 0.88 0.82 0.87 0.79 0.87 0.93 0.81 0.79 0.80 | 0.81 0.74 0.82 0.74 1.35 0.78 1.39 0.77 0.79 1.12 |
| ECM–receptor interactions | comp53019_c0 comp42008_c1 comp39439_c0 comp47792_c0 comp16281_c0 comp41102_c0 comp47172_c0 comp52938_c0 comp23347_c0 comp62976_c0 comp44017_c0 comp48402_c0 | Laminin subunit gamma-1 Nidogen-1 MAM and LDL-receptor class A domain-containing protein C10orf112 Ryncolin-2 Basement membrane-specific heparan sulfate proteoglycan core protein von Willebrand factor D and EGF domain-containing protein Laminin subunit alpha Laminin subunit beta-1 Techylectin-5B Transforming growth factor–beta-induced protein ig-h3 E2C1D1_HARSA Teneurin-3 Integrin alpha-8 | 0.74 0.57 0.80 0.98 0.91 0.98 0.73 0.77 0.60 0.58 1.28 0.83 | 0.56 0.49 0.82 1.21 0.60 0.83 0.54 0.60 0.48 0.53 1.04 0.84 |
| MAPK signaling pathway | comp50940_c0 comp41423_c0 comp50628_c0 comp44493_c0 comp53980_c0 comp50625_c0 comp51079_c0 | Caspase-3 Mitogen-activated protein kinase ERK-A Serine/threonine-protein kinase 3 Fasciclin-2 JNK-interacting protein 3 Cdc42 homolog Serine/threonine-protein kinase 3 | 0.86 1.20 0.83 0.88 0.82 0.62 0.79 | 0.79 1.21 0.78 0.76 0.81 0.76 0.80 |
| Metabolism of xenobiotics by cytochrome P450 | comp40890_c0 comp25721_c0 | UDP-glucuronosyltransferase 2B13 Glutathione S-transferase Mu 2 | 1.23 0.67 | 1.17 0.60 |
| Focal adhesion | comp53019_c0 comp42008_c1 comp53679_c0 comp39439_c0 comp47792_c0 comp434052_c0 comp41423_c0 comp16281_c0 comp41102_c0 comp50625_c0 comp47172_c0 comp51983_c0 comp36045_c0 comp37473_c0 comp52938_c0 comp23347_c0 comp40576_c0 comp62976_c0 comp44017_c0 comp52761_c0 comp48402_c0 | Laminin subunit gamma-1 Nidogen-1 Vascular endothelial growth factor receptor 1 MAM and LDL-receptor class A domain-containing protein C10orf112 Ryncolin-2 Myosin regulatory light chain 2 Mitogen-activated protein kinase ERK-A Basement membrane-specific heparan sulfate proteoglycan core protein von Willebrand factor D and EGF domain-containing protein Cdc42 homolog Laminin subunit alpha Dedicator of cytokinesis protein 2 MICAL-like protein 2 Guanine nucleotide-releasing factor 2 Laminin subunit beta-1 Techylectin-5B Talin-2 Transforming growth factor–beta-induced protein ig-h3 E2C1D1_HARSA Teneurin-3 Platelet-derived growth factor receptor alpha Integrin alpha-8 | 0.74 0.57 0.67 0.80 0.98 0.79 1.20 0.91 0.98 0.62 0.73 0.84 0.87 0.86 0.77 0.60 0.88 0.58 1.28 0.76 0.83 | 0.56 0.49 0.63 0.82 1.21 1.35 1.21 0.60 0.83 0.76 0.54 0.80 0.78 0.83 0.60 0.48 0.82 0.53 1.04 0.83 0.84 |
| Peroxisomes | comp96590_c0 comp52167_c0 comp40945_c0 | ATP-binding cassette subfamily D member 3 Superoxide dismutase [Cu-Zn] Superoxide dismutase [Cu-Zn] | 0.66 1.36 1.23 | 0.60 1.07 1.51 |
| Antigen processing and presentation | comp7092_c0 | Heat shock 70 kDa protein cognate 3 | 0.89 | 0.78 |
| Toll-like receptor signaling pathway | comp41423_c0 comp50625_c0 | Mitogen-activated protein kinase ERK-A Cdc42 homolog | 1.20 0.62 | 1.21 0.76 |
| T cell receptor signaling pathway | comp48402_c0 comp41423_c0 | Integrin alpha-8 Mitogen-activated protein kinase ERK-A | 0.83 1.20 | 0.84 1.21 |
| B cell receptor signaling pathway | comp41423_c0, comp50625_c0 | Mitogen-activated protein kinase ERK-A Cdc42 homolog | 1.20 0.62 | 1.21 0.76 |
| TNF signaling pathway | comp50940_c0 comp41423_c0 comp44944_c0 | Caspase-3 Mitogen-activated protein kinase ERK-A Dynamin-1-like protein | 0.86 1.20 0.85 | 0.79 1.21 0.81 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gao, B.; Ren, X.; Lv, J.; Meng, X.; Liu, P.; Li, J. Proteomic Analysis Associated with the Immune Response in Hemocytes of Portunus trituberculatus Challenged with Vibrio parahaemolyticus. Fishes 2022, 7, 259. https://doi.org/10.3390/fishes7050259
Gao B, Ren X, Lv J, Meng X, Liu P, Li J. Proteomic Analysis Associated with the Immune Response in Hemocytes of Portunus trituberculatus Challenged with Vibrio parahaemolyticus. Fishes. 2022; 7(5):259. https://doi.org/10.3390/fishes7050259
Chicago/Turabian StyleGao, Baoquan, Xianyun Ren, Jianjian Lv, Xianliang Meng, Ping Liu, and Jian Li. 2022. "Proteomic Analysis Associated with the Immune Response in Hemocytes of Portunus trituberculatus Challenged with Vibrio parahaemolyticus" Fishes 7, no. 5: 259. https://doi.org/10.3390/fishes7050259