
Climate-Driven Synchrony in Anchovy Fluctuations: A Pacific-Wide Comparison


Abstract
:1. Introduction
2. Materials and Methods
2.1. SDR Data
2.2. Catch Data
2.3. Large-Scale Climate Indices
2.4. Local-Scale Environmental Variables
2.5. Data Analyses
3. Results
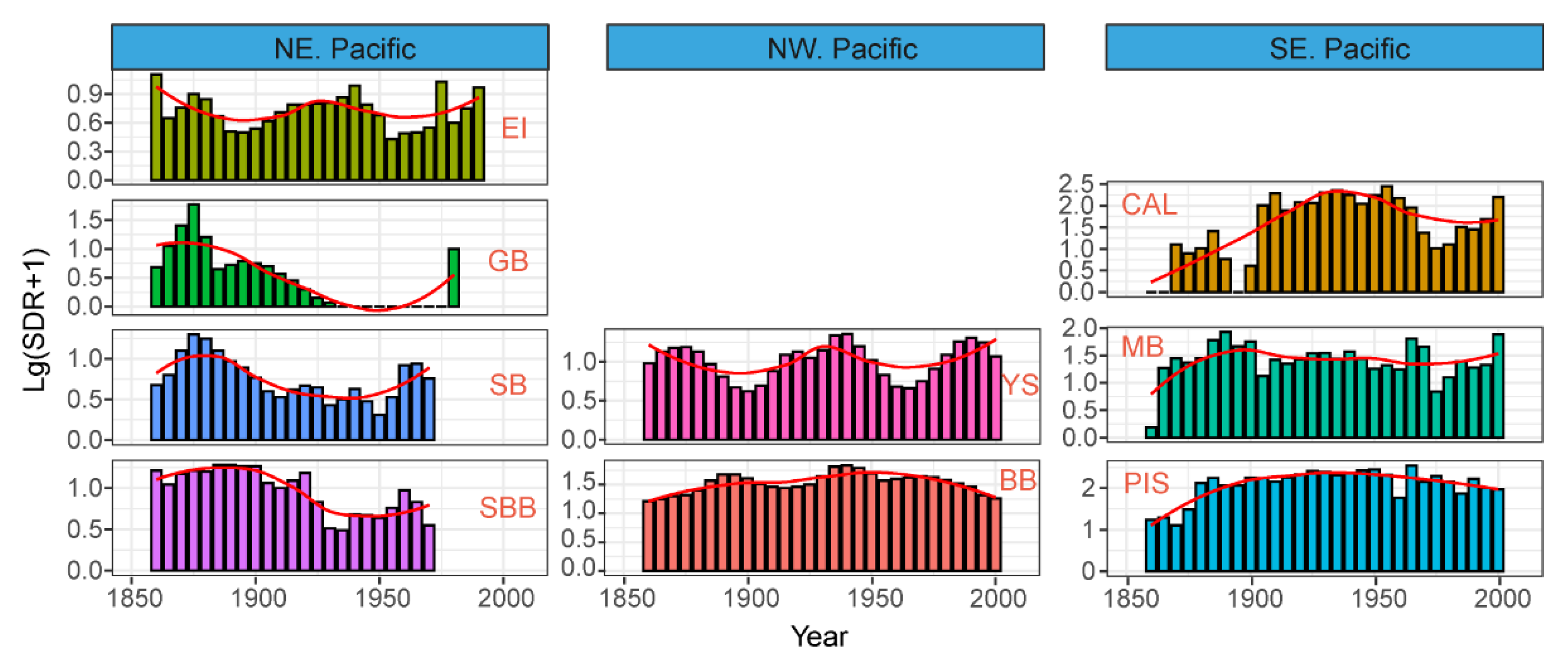
3.1. Major SDR Patterns in the Pan-Pacific
3.2. Long-Term Changes in Anchovy Catch
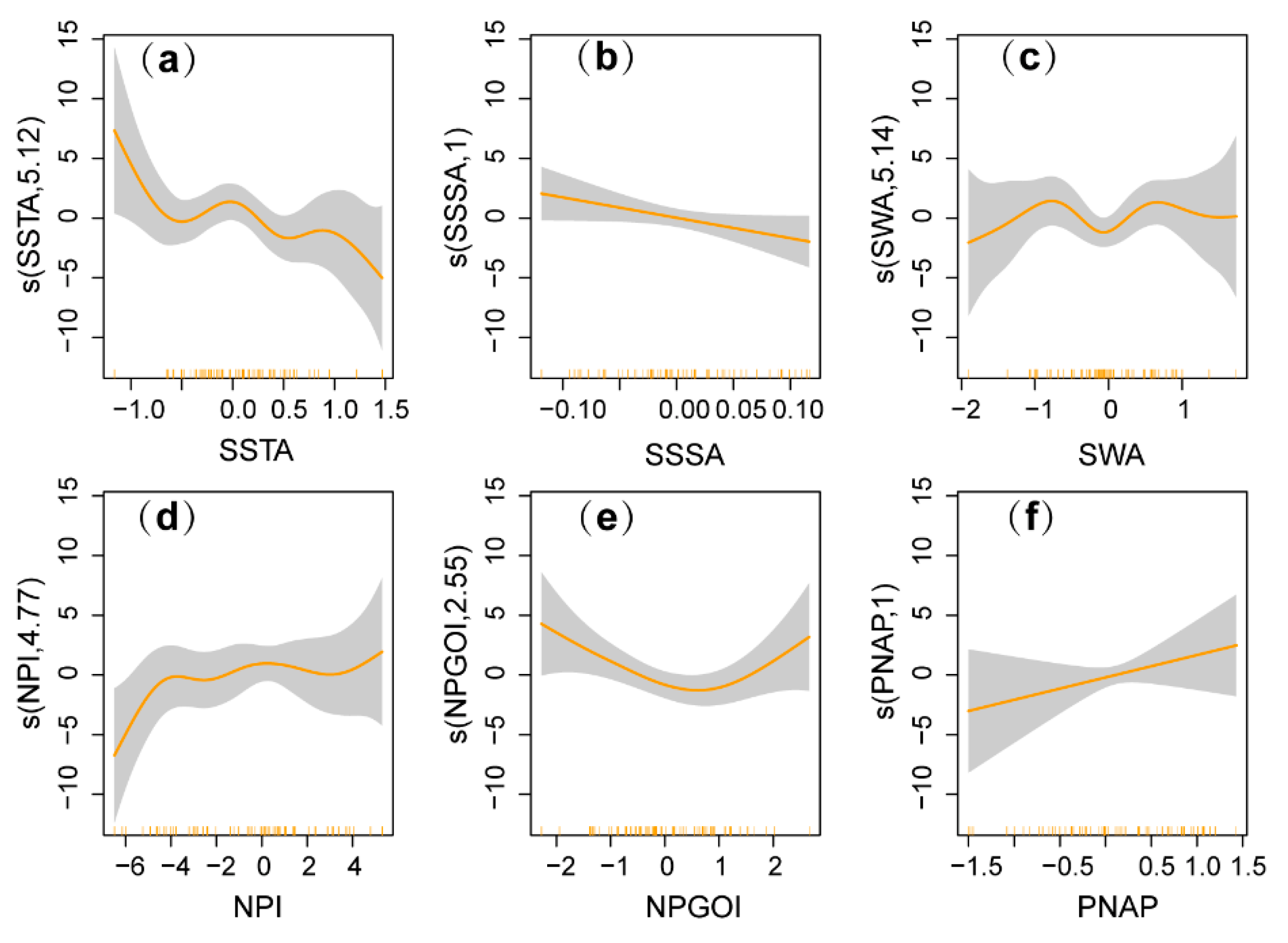
3.3. Results of the GAMs
4. Discussion
4.1. Consideration of the Method Adopted to Assess the Differences in SDR
4.2. Multiple Modes of Variability in Anchovy Fluctuations
4.3. Key Process Affecting Anchovy Fluctuations
4.4. Broader Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alheit, J.; Pohlmann, T.; Casini, M.; Greve, W.; Hinrichs, R.; Mathis, M.; O’Driscoll, K.; Vorberg, R.; Wagner, C. Climate variability drives anchovies and sardines into the North and Baltic Seas. Prog. Oceanogr. 2012, 96, 128–139. [Google Scholar] [CrossRef]
- Martín, P.; Sabatés, A.; Lloret, J.; Martin-Vide, J. Climate modulation of fish populations: The role of the Western Mediterranean Oscillation (WeMO) in sardine (Sardina pilchardus) and anchovy (Engraulis encrasicolus) production in the north-western Mediterranean. Clim. Chang. 2012, 110, 925–939. [Google Scholar] [CrossRef]
- Baumgartner, T.R.; Soutar, A.; Ferreira-Bartrina, V. Reconstruction of the history of Pacific sardine and northern anchovy populations over the past two millenia from sediments of the Santa Barbara Basin, California. Calif. Coop. Ocean. Fish. 1992, 33, 24–40. [Google Scholar]
- Guiñez, M.; Valdés, J.; Sifeddine, A.; Boussafir, M.; Dávila, P.M. Anchovy population and ocean-climatic fluctuations in the Humboldt Current System during the last 700 years and their implications. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2014, 415, 210–224. [Google Scholar] [CrossRef]
- Kuwae, M.; Yamamoto, M.; Sagawa, T.; Ikehara, K.; Irino, T.; Takemura, K.; Takeoka, H.; Sugimoto, T. Multidecadal, centennial, and millennial variability in sardine and anchovy abundances in the western North Pacific and climate–fish linkages during the late Holocene. Prog. Oceanogr. 2017, 159, 86–98. [Google Scholar] [CrossRef]
- Checkley, D.M.; Asch, R.G.; Rykaczewski, R.R. Climate, Anchovy, and Sardine. Ann. Rev. Mar. Sci. 2017, 9, 469–493. [Google Scholar] [CrossRef] [Green Version]
- MacCall, A.D. Mechanisms of low-frequency fluctuations in sardine and anchovy populations. In Climate Change and Small Pelagic Fish; Checkley, D., Alheit, J., Oozeki, Y., Roy, C., Eds.; Cambridge University Press: Cambridge, UK, 2009; pp. 285–299. [Google Scholar]
- Schwartzlose, R.A.; Alheit, J.; Bakun, A.; Baumgartner, T.R.; Cloete, R.; Crawford, R.J.M.; Fletcher, W.J.; Green-Ruiz, Y.; Hagen, E.; Kawasaki, T.; et al. Worldwide large-scale fluctuations of sardine and anchovy populations. S. Afr. J. Mar. Sci. 1999, 21, 289–347. [Google Scholar] [CrossRef]
- McClatchie, S.; Hendy, I.L.; Thompson, A.R.; Watson, W. Collapse and recovery of forage fish populations prior to commercial exploitation. Geophys. Res. Lett. 2017, 44, 1877–1885. [Google Scholar] [CrossRef]
- Salvatteci, R.; Field, D.; Gutierrez, D.; Baumgartner, T.; Ferreira, V.; Ortlieb, L.; Sifeddine, A.; Grados, D.; Bertrand, A. Multifarious anchovy and sardine regimes in the Humboldt Current System during the last 150 years. Glob. Chang. Biol. 2018, 24, 1055–1068. [Google Scholar] [CrossRef]
- Soutar, A.; Isaacs, J.D. History of fish populations inferred from fish scales in anaerobic sediments off California. Calif. Coop. Ocean. Fish. 1969, 13, 63–70. [Google Scholar]
- Wright, C.A.; Dallimore, A.; Thomson, R.E.; Patterson, R.T.; Ware, D.M. Late Holocene paleofish populations in Effingham Inlet, British Columbia, Canada. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2005, 224, 367–384. [Google Scholar] [CrossRef]
- Valdés, J.; Ortlieb, L.; Gutierrez, D.; Marinovic, L.; Vargas, G.; Sifeddine, A. 250 years of sardine and anchovy scale deposition record in Mejillones Bay, northern Chile. Prog. Oceanogr. 2008, 79, 198–207. [Google Scholar] [CrossRef]
- Zhou, X.; Sun, Y.; Huang, W.; Smol, J.P.; Tang, Q.; Sun, L. The Pacific decadal oscillation and changes in anchovy populations in the Northwest Pacific. J. Asian. Earth. Sci. 2015, 114, 504–511. [Google Scholar] [CrossRef]
- Jia, H.; Sun, Y.; Zhao, M.; Yang, Z.; Tang, Q. Fish-scale-deposition information and spatial distribution in typical fishery area of the Yellow Sea and East China Sea. J. Fish. China 2008, 32, 584–591. [Google Scholar]
- Huang, J.; Sun, Y.; Jia, H.; Tang, Q. Last 150-Year Variability in Japanese Anchovy (Engraulis japonicus) Abundance Based on the Anaerobic Sediments of the Yellow Sea Basin in the Western North Pacific. J. Ocean. Univ. China 2016, 15, 131–136. [Google Scholar] [CrossRef]
- Li, H.; Tang, Q.; Ito, S.-I.; Sun, Y. Evidence of bottom-up effects of climate on Japanese anchovy (Engraulis japonicus) in the western North Pacific. J. Oceanogr. 2021, 77, 589–605. [Google Scholar] [CrossRef]
- Li, H.; Tang, Q.; Sun, Y. Response of Japanese anchovy (Engraulis japonicus) to the Pacific Decadal Oscillation in the Yellow Sea over the past 400 a. Acta Oceanol. Sin. 2022; in press. [Google Scholar] [CrossRef]
- Holmgren, D. Decadel-Centennial Variability in Marine Ecosystem of the Northeast Pacific Ocean: The Use of Fish Scales Deposition in Sediments; University of Washington: Washington, DC, USA, 2001. [Google Scholar]
- Soutar, A.; Isaacs, J.D. Abundance of pelagic fish during the 19th and 20th centuries as recorded in anaerobic sediment off the Californias. Fish. Bull. 1974, 72, 257–273. [Google Scholar]
- Holmgren-Urba, D.; Baumgartner, T.R. A 250-year history of pelagic fish abundances from the anaerobic sediments of the central Gulf of California. Calif. Coop. Ocean. Fish. 1993, 34, 60–68. [Google Scholar]
- Gutiérrez, D.; Sifeddine, A.; Field, D.B.; Ortlieb, L.; Vargas, G.; Chávez, F.P.; Velazco, F.; Ferreira, V.; Tapia, P.; Salvatteci, R.; et al. Rapid reorganization in ocean biogeochemistry off Peru towards the end of the Little Ice Age. Biogeosciences 2009, 6, 835–848. [Google Scholar] [CrossRef] [Green Version]
- FAO. Fishery and Aquaculture Statistics. In Global Capture Production 1950–2015 (FishstatJ); FAO: Rome, Italy, 2017. [Google Scholar]
- Chavez, F.P.; Ryan, J.; Lluch-Cota, S.E.; Ñiquen, C.M. From Anchovies to Sardines and Back: Multidecadal Change in the Pacific Ocean. Science 2003, 299, 217–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Lorenzo, E.; Schneider, N.; Cobb, K.M.; Franks, P.J.S.; Chhak, K.; Miller, A.J.; McWilliams, J.C.; Bograd, S.J.; Arango, H.; Curchitser, E.; et al. North Pacific Gyre Oscillation links ocean climate and ecosystem change. Geophys. Res. Lett. 2008, 35, L08607. [Google Scholar] [CrossRef] [Green Version]
- Alheit, J.; Bakun, A. Population synchronies within and between ocean basins: Apparent teleconnections and implications as to physical–biological linkage mechanisms. J. Mar. Syst. 2010, 79, 267–285. [Google Scholar] [CrossRef]
- Litzow, M.A.; Mueter, F.J.; Hobday, A.J. Reassessing regime shifts in the North Pacific: Incremental climate change and commercial fishing are necessary for explaining decadal-scale biological variability. Glob. Chang. Biol. 2014, 20, 38–50. [Google Scholar] [CrossRef]
- Litzow, M.A.; Ciannelli, L.; Puerta, P.; Wettstein, J.J.; Rykaczewski, R.R.; Opiekun, M. Non-stationary climate–salmon relationships in the Gulf of Alaska. Proc. Royal Soc. B. 2018, 285, 20181855. [Google Scholar] [CrossRef] [Green Version]
- Litzow, M.A.; Malick, M.J.; Bond, N.A.; Cunningham, C.J.; Gosselin, J.L.; Ward, E.J. Quantifying a Novel Climate Through Changes in PDO-Climate and PDO-Salmon Relationships. Geophys. Res. Lett. 2020, 47, e2020GL087972. [Google Scholar] [CrossRef]
- Ma, S.; Liu, Y.; Li, J.; Fu, C.; Ye, Z.; Sun, P.; Yu, H.; Cheng, J.; Tian, Y. Climate-induced long-term variations in ecosystem structure and atmosphere-ocean-ecosystem processes in the Yellow Sea and East China Sea. Prog. Oceanogr. 2019, 175, 183–197. [Google Scholar] [CrossRef]
- Jung, H.K.; Rahman, S.M.M.; Kang, C.-K.; Park, S.-Y.; Heon Lee, S.; Je Park, H.; Kim, H.-W.; Lee, C.I. The influence of climate regime shifts on the marine environment and ecosystems in the East Asian Marginal Seas and their mechanisms. Deep Sea Res. Part II Top. Stud. Oceanogr. 2017, 143, 110–120. [Google Scholar] [CrossRef]
- Overland, J.E.; Alheit, J.; Bakun, A.; Hurrell, J.W.; Mackas, D.L.; Miller, A.J. Climate controls on marine ecosystems and fish populations. J. Mar. Syst. 2010, 79, 305–315. [Google Scholar] [CrossRef]
- Ma, S.; Tian, Y.; Fu, C.; Yu, H.; Li, J.; Liu, Y.; Cheng, J.; Wan, R.; Watanabe, Y. Climate-induced nonlinearity in pelagic communities and non-stationary relationships with physical drivers in the Kuroshio ecosystem. Fish Fish. 2020, 22, 1–17. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix, Version 0.84. 2017.
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; Chapman and Hall/CRC: New York, NY, USA, 2017; p. 496. [Google Scholar]
- Hastie, T.J.; Tibshirani, R.J. Monographs on Statistics and Applied Probability in Generalized Linear Models; Chapman & Hall: London, UK, 1990; Volume 43, pp. 205–208. [Google Scholar]
- Zuur, A.; Ieno, E.N.; Smith, G.M. Analyzing Ecological Data; Springer: New York, NY, USA, 2007; p. 672. [Google Scholar]
- Vrieze, S.I. Model selection and psychological theory: A discussion of the differences between the Akaike information criterion (AIC) and the Bayesian information criterion (BIC). Psychol. Methods 2012, 17, 228–243. [Google Scholar] [CrossRef] [Green Version]
- McHugh, J.L. Meristic Variations and Populations of Northern Anchovy (Engraulis Mordax Mordax). Scripps Inst. Oceanogr. Bull. 1951, 6, 123–160. [Google Scholar] [CrossRef]
- Oozeki, Y.; Ñiquen Carranza, M.; Takasuka, A.; Ayón Dejo, P.; Kuroda, H.; Tam Malagas, J.; Okunishi, T.; Vásquez Espinoza, L.; Gutiérrez Aguilar, D.; Okamura, H.; et al. Synchronous multi-species alternations between the northern Humboldt and Kuroshio Current systems. Deep Sea Res. Part II Top. Stud. Oceanogr. 2019, 159, 11–21. [Google Scholar] [CrossRef]
- Izquierdo-Peña, V.; Lluch-Cota, S.E.; Hernandez-Rivas, M.E.; Martínez-Rincón, R.O. Revisiting the Regime Problem hypothesis: 25 years later. Deep Sea Res. Part II Top. Stud. Oceanogr. 2019, 159, 4–10. [Google Scholar] [CrossRef]
- Takasuka, A.; Oozeki, Y.; Kubota, H.; Lluch-Cota, S.E. Contrasting spawning temperature optima: Why are anchovy and sardine regime shifts synchronous across the North Pacific? Prog. Oceanogr. 2008, 77, 225–232. [Google Scholar] [CrossRef]
- Hilborn, R.; Quinn, T.P.; Schindler, D.E.; Rogers, D.E. Biocomplexity and fisheries sustainability. Proc. Natl. Acad. Sci. USA 2003, 100, 6564–6568. [Google Scholar] [CrossRef] [Green Version]
- Rogers, L.A.; Schindler, D.E.; Lisi, P.J.; Holtgrieve, G.W.; Leavitt, P.R.; Bunting, L.; Finney, B.P.; Selbie, D.T.; Chen, G.; Gregory-Eaves, I.; et al. Centennial-scale fluctuations and regional complexity characterize Pacific salmon population dynamics over the past five centuries. Proc. Natl. Acad. Sci. USA 2013, 110, 1750–1755. [Google Scholar] [CrossRef] [Green Version]
- Pauly, D.; Christensen, V. Primary production required to sustain global fisheries. Nature 1995, 374, 255–257. [Google Scholar] [CrossRef]
- Rykaczewski, R.R.; Checkley, D.M. Influence of ocean winds on the pelagic ecosystem in upwelling regions. Proc. Natl. Acad. Sci. USA 2008, 105, 1965–1970. [Google Scholar] [CrossRef] [Green Version]
- Ayón, P.; Swartzman, G.; Bertrand, A.; Gutiérrez, M.; Bertrand, S. Zooplankton and forage fish species off Peru: Large-scale bottom-up forcing and local-scale depletion. Prog. Oceanogr. 2008, 79, 208–214. [Google Scholar] [CrossRef]
- Roemmich, D.; McGowan, J. Climatic Warming and the Decline of Zooplankton in the California Current. Science 1995, 267, 1324–1326. [Google Scholar] [CrossRef] [PubMed]
- Fiechter, J.; Rose, K.A.; Curchitser, E.N.; Hedstrom, K.S. The role of environmental controls in determining sardine and anchovy population cycles in the California Current: Analysis of an end-to-end model. Prog. Oceanogr. 2015, 138, 381–398. [Google Scholar] [CrossRef] [Green Version]
- Bakun, A. Global climate change and intensification of coastal ocean upwelling. Science 1990, 247, 198–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belkin, I.M. Rapid warming of Large Marine Ecosystems. Prog. Oceanogr. 2009, 81, 207–213. [Google Scholar] [CrossRef]
- Yatsu, A.; Chiba, S.; Yamanaka, Y.; Ito, S.I.; Shimizu, Y.; Kaeriyama, M.; Watanabe, Y. Climate forcing and the Kuroshio/Oyashio ecosystem. ICES J. Mar. Sci. 2013, 70, 922–933. [Google Scholar] [CrossRef] [Green Version]
- Hollowed, A.B.; Barange, M.; Beamish, R.J.; Brander, K.; Cochrane, K.; Drinkwater, K.; Foreman, M.G.G.; Hare, J.A.; Holt, J.; Ito, S.-i.; et al. Projected impacts of climate change on marine fish and fisheries. ICES J. Mar. Sci. 2013, 70, 1023–1037. [Google Scholar] [CrossRef] [Green Version]
- Jang, C.J.; Park, J.; Park, T.; Yoo, S. Response of the ocean mixed layer depth to global warming and its impact on primary production: A case for the North Pacific Ocean. ICES J. Mar. Sci. 2011, 68, 996–1007. [Google Scholar] [CrossRef] [Green Version]
- Lynam, C.P.; Llope, M.; Möllmann, C.; Helaouët, P.; Bayliss-Brown, G.A.; Stenseth, N.C. Interaction between top-down and bottom-up control in marine food webs. Proc. Natl. Acad. Sci. USA 2017, 114, 1952–1957. [Google Scholar] [CrossRef] [Green Version]
- Pikitch, E.K.; Santora, C.; Babcock, E.A.; Bakun, A.; Bonfil, R.; Conover, D.O.; Dayton, P.; Doukakis, P.; Fluharty, D.; Heneman, B.; et al. Ecosystem-Based Fishery Management. Science 2004, 305, 346–347. [Google Scholar] [CrossRef]
- Tian, Y.; Kidokoro, H.; Watanabe, T.; Iguchi, N. The late 1980s regime shift in the ecosystem of Tsushima warm current in the Japan/East Sea: Evidence from historical data and possible mechanisms. Prog. Oceanogr. 2008, 77, 127–145. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
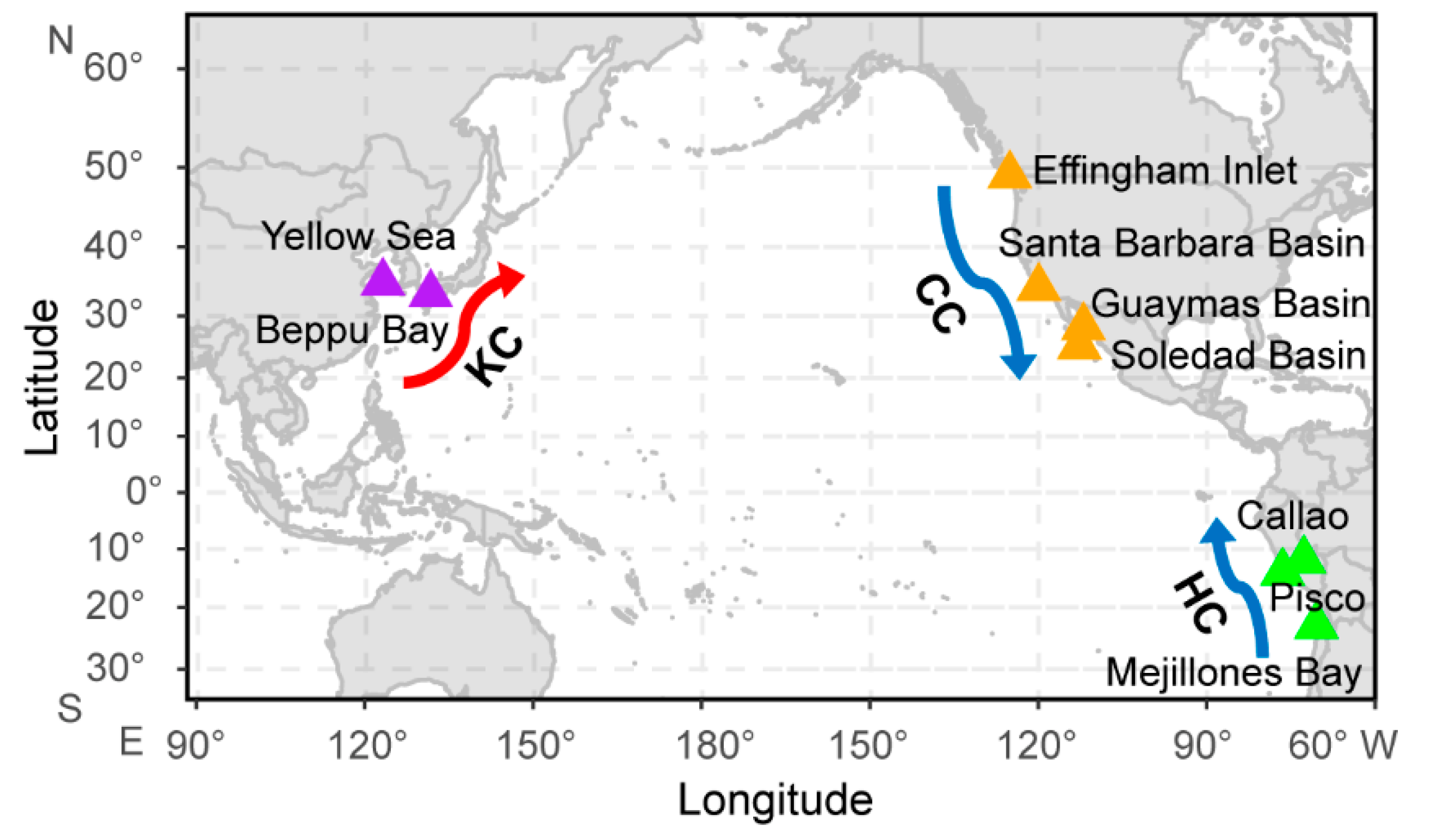
| Site | Location | Period | Mean Sample Interval (Years) | Reference |
|---|---|---|---|---|
| Effingham Inlet | southwest British Columbia, Canada | 1860–1990 | 2–5 | [19] |
| Santa Barbara Basin | southern California, USA | 1860–1970 | 5 | [20] |
| Guaymas Basin | central Gulf of California, Mexico | 1860–1980 | 10 | [21] |
| Soledad Basin | southern Baja California, Mexico | 1860–1970 | 5 | [20] |
| Callao | central Peru | 1860–2000 | 7 | [22] |
| Pisco | central Peru | 1860–2000 | 6 | [22] |
| Mejillones Bay | northern Chile | 1860–2000 | 3 | [13] |
| Yellow Sea | eastern China | 1860–2000 | 5 | [18] |
| Beppu Bay | southwest Japan | 1860–2000 | 7 | [5] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Zhang, X.; Zhang, Y.; Liu, Q.; Liu, F.; Li, D.; Zhang, H. Climate-Driven Synchrony in Anchovy Fluctuations: A Pacific-Wide Comparison. Fishes 2022, 7, 193. https://doi.org/10.3390/fishes7040193
Li H, Zhang X, Zhang Y, Liu Q, Liu F, Li D, Zhang H. Climate-Driven Synchrony in Anchovy Fluctuations: A Pacific-Wide Comparison. Fishes. 2022; 7(4):193. https://doi.org/10.3390/fishes7040193
Chicago/Turabian StyleLi, Haoyu, Xiaonan Zhang, Yang Zhang, Qi Liu, Fengwen Liu, Donglin Li, and Hucai Zhang. 2022. "Climate-Driven Synchrony in Anchovy Fluctuations: A Pacific-Wide Comparison" Fishes 7, no. 4: 193. https://doi.org/10.3390/fishes7040193