
A New and Profitable Protocol to DNA Extraction in Limnospira maxima
 , ,
, , 
Abstract
:1. Introduction
2. Experimental Design
3. Procedure
3.1. Cyanobacteria Strain, Growth, and Production Medium
3.2. Molecular Identification of the Strain Sampling
3.3. Genomic DNA Extraction
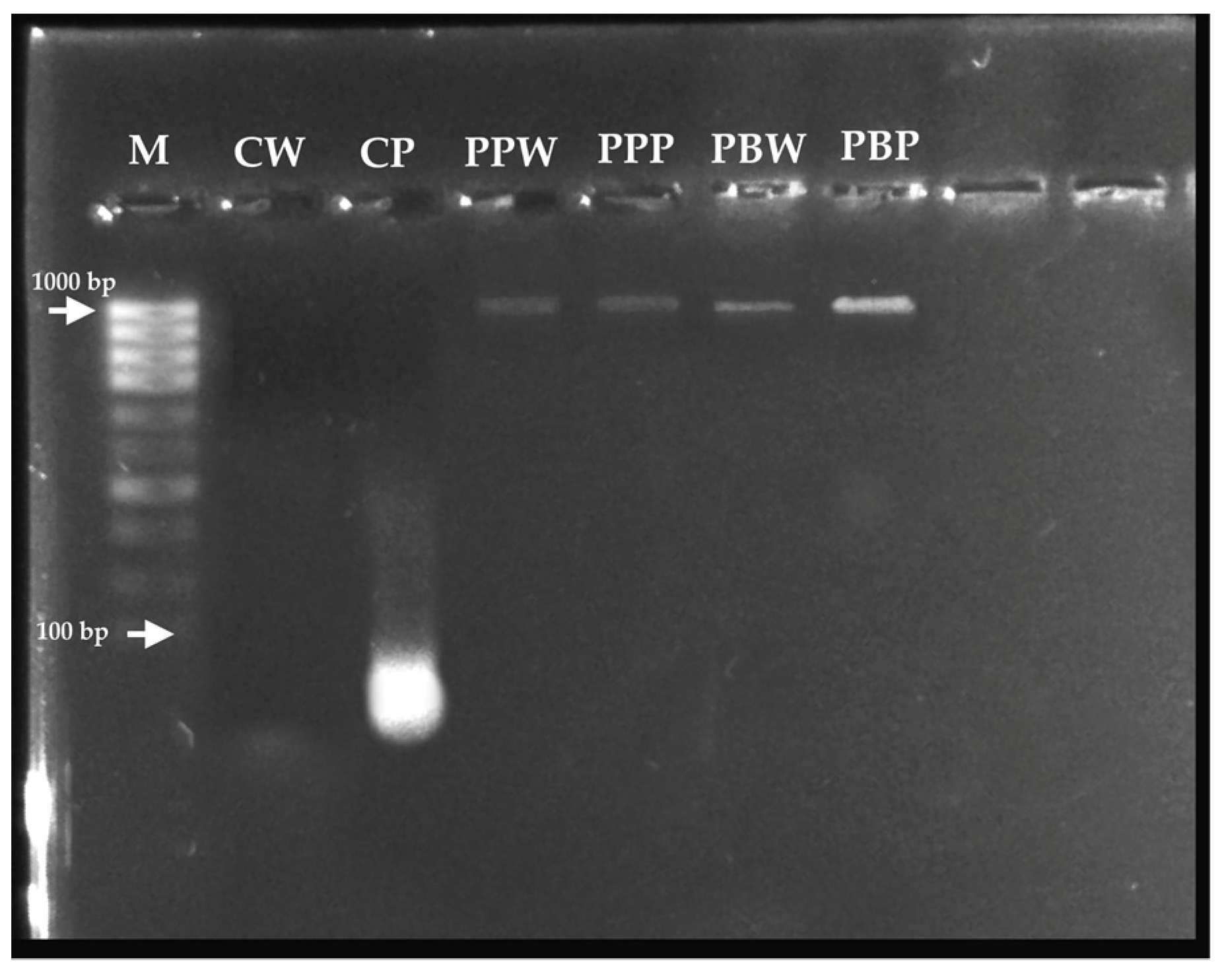
3.4. DNA Amplification by Polymerase Chain Reaction (PCR)
3.5. Sequencing of PCR Products and Sequence Analysis
3.6. Statistical Analysis
4. Results
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gardner, N.L. The Marine Algae of the Pacific Coast of North America; University of California Publications in Botany: Berkeley, CA, USA, 1937; Volume 8, pp. 1–3. [Google Scholar] [CrossRef] [Green Version]
- Nowicka-Krawczyk, P.; Mühlsteinová, R.; Hauer, T. Detailed characterization of the Arthrospira type species separating commercially grown taxa into the new genus Limnospira (Cyanobacteria). Sci. Rep. 2019, 9, 694. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciferri, O. Spirulina, the edible microorganism. Microbiol. Rev. 1983, 47, 551–578. [Google Scholar] [CrossRef] [PubMed]
- Pineda-Rodríguez, Y.Y.; Vegliante, D.; Ariza, A.; Pico, A.; Herazo, D.; Castro, G.R.; Vallejo, A.; Jarma-Orozco, A.; Jaraba-Navas, J.D. Effect of two culture media (Jourdan and Zarrouk) on the production of chlorophyll and phycobiliproteins of Arthrospira maxima. In Proceedings of the Asociacion Colombiana de Ciencias Biologicas, Virtual, 15 September 2021; Red Colombiana de Biología: Riohacha, Colombia, 2021; pp. 136–139. [Google Scholar]
- Milia, M.; Corrias, F.; Addis, P.; Chini Zitelli, G.; Cicchi, B.; Torzillo, G.; Andreotti, V.; Angioni, A. Influence of different light sources on the biochemical composition of Arthrospira spp. grown in model systems. Foods 2022, 11, 399. [Google Scholar] [CrossRef] [PubMed]
- AlFadhly, N.K.Z.; Alhelfi, N.; Altemimi, A.B.; Verma, D.K.; Cacciola, F. Tendencies affecting the growth and cultivation of genus Spirulina: An investigative review on current trends. Plants 2022, 11, 3063. [Google Scholar] [CrossRef] [PubMed]
- Morin, N.; Vallaeys, T.; Hendrickx, L.; Natalie, L.; Wilmotte, A. An efficient DNA isolation protocol for filamentous cyanobacteria of the genus Arthrospira. J. Microbiol. Methods 2010, 80, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Furmaniak, M.A.; Misztak, A.E.; Franczuk, M.D.; Wilmotte, A.; Waleron, M.; Waleron, K.F. Edible cyanobacterial genus Arthrospira: Actual state of the art in cultivation methods, genetics, and application in medicine. Front. Microbiol. 2017, 8, 2541. [Google Scholar] [CrossRef] [PubMed]
- Pompelli, M.F.; Mendes, K.R.; Ramos, M.V.; Santos, J.N.B.; Youssef, D.T.A.; Pereira, J.D.; Endres, L.; Jarma-Orozco, A.; Solano-Gomes, R.; Jarma-Arroyo, B.; et al. Mesophyll thickness and sclerophylly among Calotropis procera morphotypes reveal water-saved adaptation to environments. J. Arid Land 2019, 11, 795–810. [Google Scholar] [CrossRef] [Green Version]
- Rajasekaran, C.; Ajeesh, C.P.M.; Balaji, S.; Shalini, M.; Siva, R.; Das, R.; Fulzele, D.P.; Kalaivani, T. Effect of modified Zarrouk’s medium on growth of different Spirulina strains. Agric. Technol. Biol. Sci. 2016, 13, 67–75. [Google Scholar]
- Abu Almakarem, A.S.; Heilman, K.L.; Conger, H.L.; Shtarkman, Y.M.; Rogers, S.O. Extraction of DNA from plant and fungus tissues in situ. BMC Res. Notes 2012, 5, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilfinger, W.W.; Mackey, K.; Chomczynski, P. Effect of pH and ionic strength on the spectrophotometric assessment of nucleic acid purity. Biotechniques 1997, 22, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Sano, H.; Wakui, A.; Kawachi, M.; Washio, J.; Abiko, Y.; Mayanagi, G.; Yamaki, K.; Tanaka, K.; Takahashi, N.; Sato, T. Profiling system of oral microbiota utilizing polymerase chain reaction-restriction fragment length polymorphism analysis. J. Oral. Biosci. 2021, 63, 292–297. [Google Scholar] [CrossRef] [PubMed]
- Sanger, F.; Coulson, A.R. A rapid method for determining sequences in DNA by primed synthesis with DNA polymerase. J. Mol. Biol. 1975, 94, 441–448. [Google Scholar] [CrossRef] [PubMed]
- Pineda-Rodriguez, Y.Y.; Pompelli, M.F.; Jarma-Orozco, A.; Jaraba-Navas, J.D.D.; Herazo-Cardenas, D.S.; Vallejo-Isaza, A.; Rodriguez-Paez, L.A. Limnospira maxima Strain SISCA 16S Ribosomal RNA Gene, Partial Sequence; National Center for Biotechnology Information: Bethesda, MD, USA, 2023. Available online: https://www.ncbi.nlm.nih.gov/nuccore/OR195505 (accessed on 2 June 2023).
- Eusebio, N.; Rego, A.; Glasser, N.R.; Castelo-Branco, R.; Balskus, E.P.; Leão, P.N. Distribution and diversity of dimetal-carboxylate halogenases in cyanobacteria. BMC Genom. 2021, 22, 633. [Google Scholar] [CrossRef] [PubMed]
- Martins, J.; Leão, P.; Ramos, V.; Vasconcelos, V. N-Terminal protease gene phylogeny reveals the potential for novel cyanobactin diversity in cyanobacteria. Mar. Drugs 2013, 11, 4902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos, V.M.C.; Castelo-Branco, R.; Leão, P.N.; Martins, J.; Carvalhal-Gomes, S.; Sobrinho da Silva, F.; Mendonça Filho, J.G.; Vasconcelos, V.M. Cyanobacterial diversity in microbial mats from the hypersaline lagoon system of Araruama, Brazil: An in-depth polyphasic study. Front. Microbiol. 2017, 8, 1233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vonshak, A. Spirulina platensis (Arthrospira): Physiology, Cell-Biology and Biotechnology, 1st ed.; CRC Press: London, UK, 1997. [Google Scholar]
- Baurain, D.; Renquin, L.; Grubisic, S.; Scheldeman, P.; Belay, A.; Wilmotte, A. Remarkable conservation of internally transcribed spacer sequences of Arthrospira (“Spirulina”) (Cyanophyceae, Cyanobacteria) strains from four continents and of recent and 30-year-old dried samples from Africa. J. Phycol. 2002, 38, 384–393. [Google Scholar] [CrossRef] [Green Version]
- Faldu, N.; Patel, S.; Vishwakarma, N.P.; Singh, A.K.; Patel, K.; Pandhi, N. Genetic diversity of marine and fresh water cyanobacteria from the Gujarat State of India. Adv. Biosci. Biotechnol. 2014, 5, 1061–1066. [Google Scholar] [CrossRef] [Green Version]
- Zhao, F.; Zhang, X.; Liang, C.; Wu, J.; Bao, Q.; Qin, S. Genome-wide analysis of restriction-modification system in unicellular and filamentous cyanobacteria. Physiol. Genom. 2006, 24, 181–190. [Google Scholar] [CrossRef] [PubMed]
- Mak, Y.M.; Ho, K.K. An improved method for the isolation of chromosomal DNA from various bacteria and cyanobacteria. Nucleic Acids Res. 1992, 20, 4101–4102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yeh, W.-J.; Chen, G.-Y.; Chen, Y.-C. Identification of a blue-green alga Arthrospira maxima using internal transcribed spacer gene sequence. AOT Acta Oceanogr. Taiwanica 2004, 42, 25–37. [Google Scholar]



{kind=link}
{kind=link}
| Extraction Method | Sample Type | |
|---|---|---|
| Wet Sample (W) | Powdered Sample (P) | |
| Cetyltrimethylammonium bromide (CTAB 2X) (Kit 1) | T1. CTAB-W | T2. CTAB-P |
| PureLink™ Genomic DNA Mini Kit (Pbact) (Kit 2) | T3. Pbact-W | T4. Pbact-P |
| PureLink® Plant Total DNA (Pplant) (Kit 3) | T5. Pplant-W | T6. Pplant-P |
| Solution | Final Concentration | 10 mL |
|---|---|---|
| dH2O (Merck KGaA, part number: 38796) | 5.6 mL | |
| 1 M Tris-HCl- 7.5 (Merck KGaA, part number: T3253) | 100 mM | 1.0 mL |
| 5 M NaCl (Merck KGaA, part number: 1064041000) | 1400 mM | 2.8 mL |
| 0.5 M EDTA-8.0 (Merck KGaA, part number: 324506) | 20 mM | 0.4 mL |
| CTAB (Merck KGaA, part number: H6269-500G) | 2% (w/v) | 0.2 g |
| PVP 10% (w/v) (Merck KGaA, part number: T3253-500G) | 1% (v/v) | 0.1 mL |
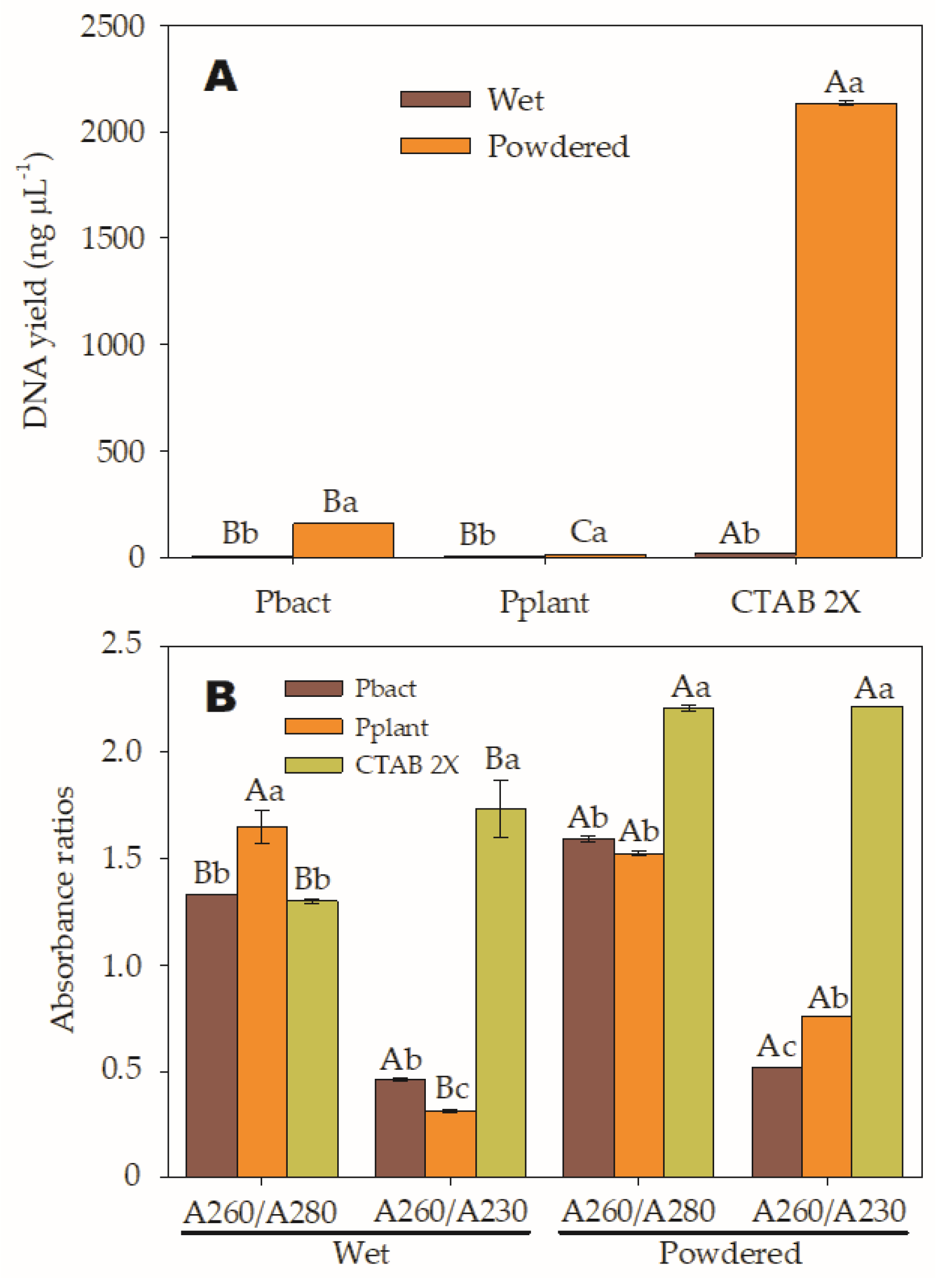
| Treatments | Nucleic Acid Concentration (ng/µL) | Purity Ratios | |
|---|---|---|---|
| A260/A280 | A260/A230 | ||
| T1. CTAB-W | 20.4 | 1.3 | 1.7 |
| T2. CTAB-P | 2134.30 | 2.2 | 2.2 |
| T3. Pbact-W | 6.9 | 1.3 | 0.4 |
| T4. Pbact-P | 157 | 1.6 | 0.5 |
| T5. Pplant-W | 3.9 | 1.6 | 0.3 |
| T6. Pplant-P | 12.5 | 1.5 | 0.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pineda-Rodriguez, Y.Y.; Pompelli, M.F.; Jarma-Orozco, A.; Rodríguez, N.V.; Rodriguez-Paez, L.A. A New and Profitable Protocol to DNA Extraction in Limnospira maxima. Methods Protoc. 2023, 6, 62. https://doi.org/10.3390/mps6040062
Pineda-Rodriguez YY, Pompelli MF, Jarma-Orozco A, Rodríguez NV, Rodriguez-Paez LA. A New and Profitable Protocol to DNA Extraction in Limnospira maxima. Methods and Protocols. 2023; 6(4):62. https://doi.org/10.3390/mps6040062
Chicago/Turabian StylePineda-Rodriguez, Yirlis Yadeth, Marcelo F. Pompelli, Alfredo Jarma-Orozco, Novisel Veitía Rodríguez, and Luis Alfonso Rodriguez-Paez. 2023. "A New and Profitable Protocol to DNA Extraction in Limnospira maxima" Methods and Protocols 6, no. 4: 62. https://doi.org/10.3390/mps6040062