
EEG-Based Mapping of Resting-State Functional Brain Networks in Patients with Parkinson’s Disease
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Protocol
2.3. EEG Signal Pre-Processing
Global and Local Noises
2.4. Source Localization Analysis
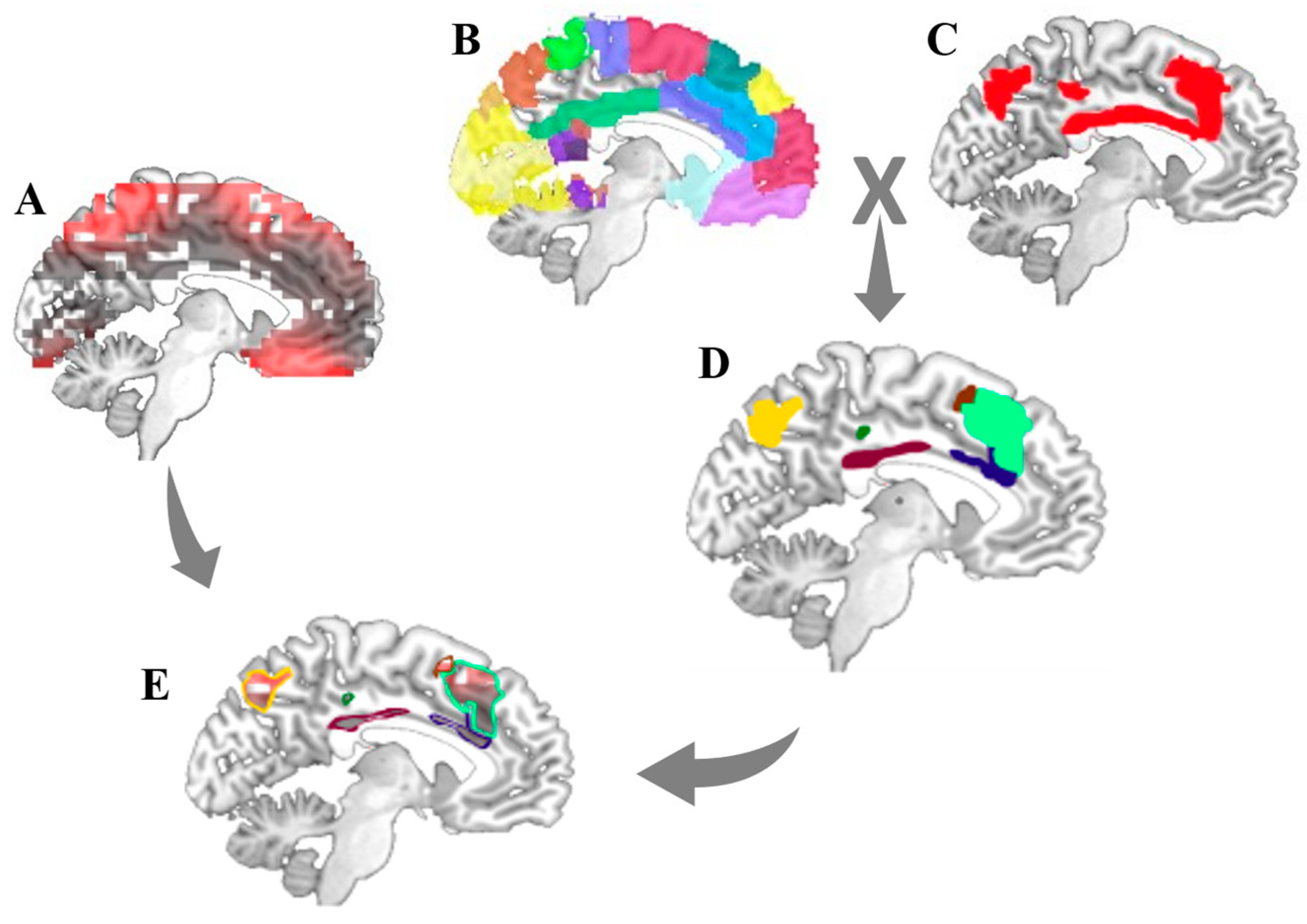
2.5. Resting-State Functional Brain Networks Masks
2.6. Temporal Partial Directed Coherence (tPDC)
2.7. Statistical Analysis
3. Results
3.1. Clinical and Demographic Characteristics of Study Participants
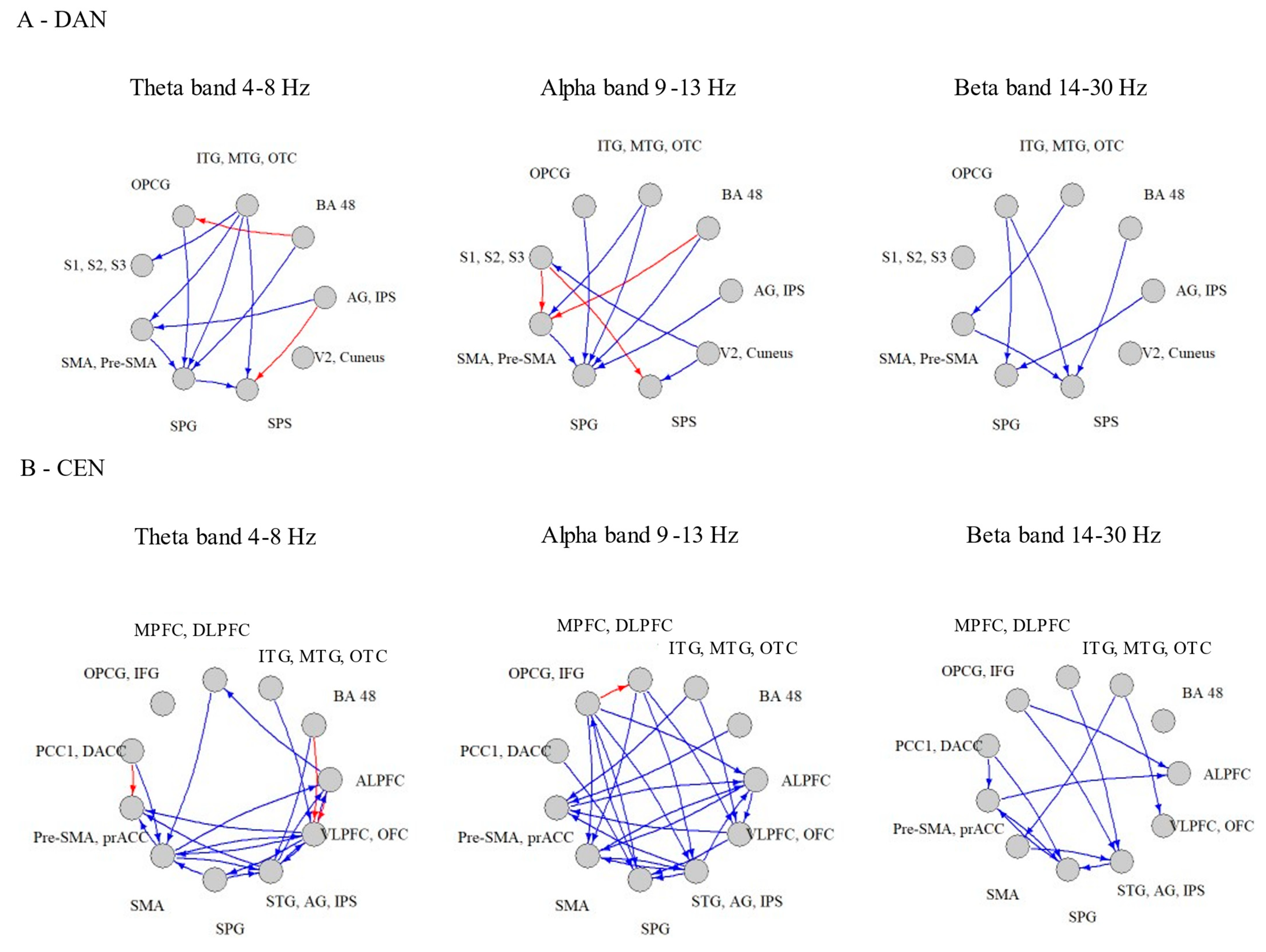
3.2. Within-Network Significant Connections
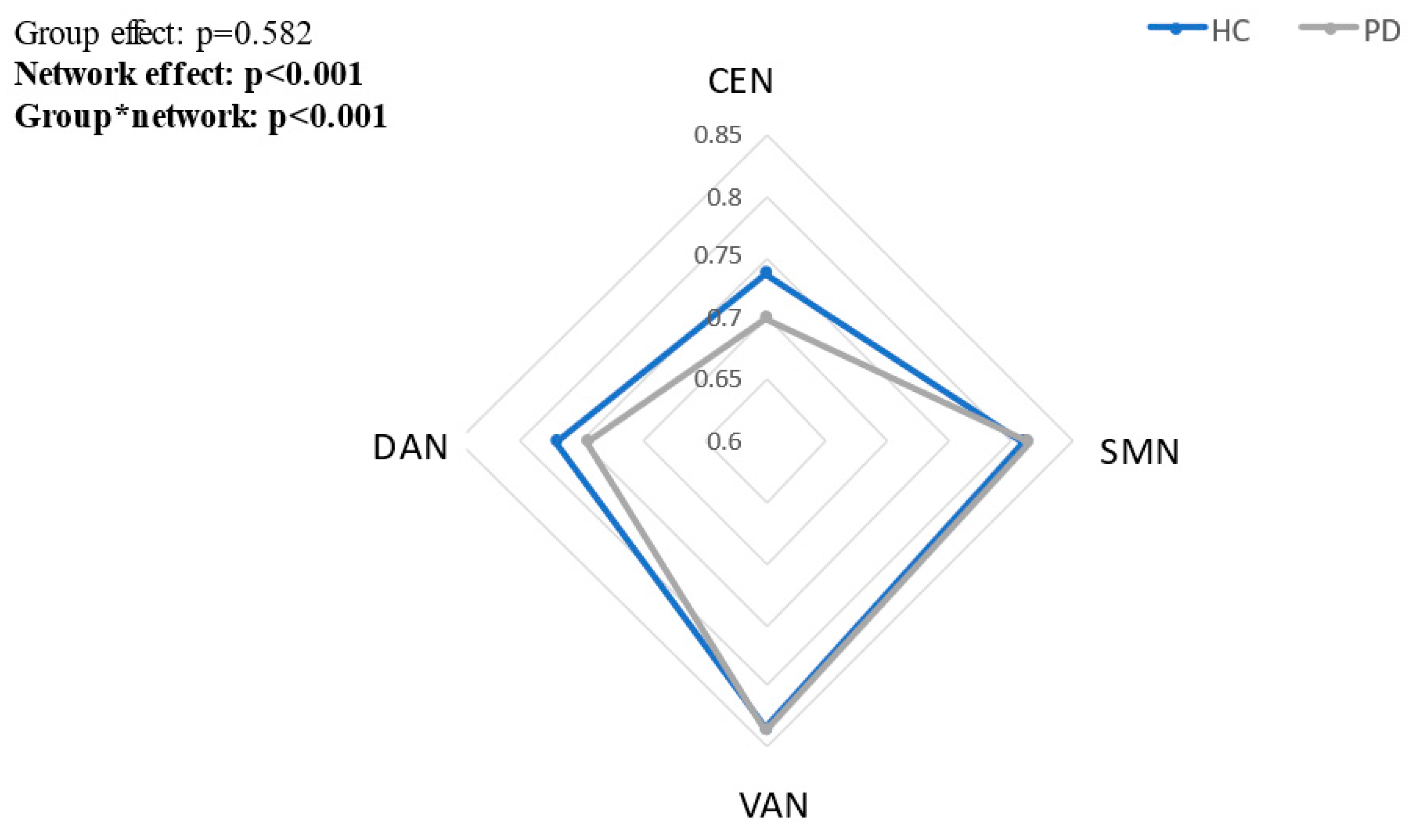
3.3. Between Group Differences in tPDCs within Each Network
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Pagano, G.; Ferrara, N.; Brooks, D.J.; Pavese, N. Age at onset and Parkinson disease phenotype. Neurology 2016, 86, 1400–1407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simon, D.K.; Tanner, C.M.; Brundin, P. Parkinson Disease Epidemiology, Pathology, Genetics, and Pathophysiology. Clin. Geriatr. Med. 2020, 36, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Svenningsson, P.; Westman, E.; Ballard, C.; Aarsland, D. Cognitive impairment in patients with Parkinson’s disease: Diagnosis, biomarkers, and treatment. Lancet Neurol. 2012, 11, 697–707. [Google Scholar] [CrossRef]
- Von Coelln, R.; Gruber-Baldini, A.L.; Reich, S.G.; Armstrong, M.J.; Savitt, J.M.; Shulman, L.M. The inconsistency and instability of Parkinson’s disease motor subtypes. Parkinsonism Relat. Disord. 2021, 88, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Kelly, V.E.; Eusterbrock, A.J.; Shumway-Cook, A. A review of dual-task walking deficits in people with Parkinson’s disease: Motor and cognitive contributions, mechanisms, and clinical implications. Parkinson’s Dis. 2012, 2012, 918719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tessitore, A.; Cirillo, M.; de Micco, R. Functional Connectivity Signatures of Parkinson’s Disease. J. Parkinson’s Dis. 2019, 9, 637–652. [Google Scholar] [CrossRef] [Green Version]
- Deco, G.; Jirsa, V.K.; McIntosh, A.R. Emerging concepts for the dynamical organization of resting-state activity in the brain. Nat. Rev. Neurosci. 2011, 12, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Logothetis, N.K.; Wandell, B.A. Interpreting the BOLD signal. Annu. Rev. Physiol. 2004, 66, 735–769. [Google Scholar] [CrossRef]
- Cole, M.W.; Ito, T.; Cocuzza, C.; Sanchez-Romero, R. The functional relevance of task-state functional connectivity. J. Neurosci. 2021, 41, 2684–2702. [Google Scholar] [CrossRef]
- Ito, T.; Hearne, L.; Mill, R.; Cocuzza, C.; Cole, M.W. Discovering the Computational Relevance of Brain Network Organization. Trends Cogn. Sci. 2020, 24, 25–38. [Google Scholar] [CrossRef]
- Toll, R.T.; Wu, W.; Naparstek, S.; Zhang, Y.; Narayan, M.; Patenaude, B.; Angeles, C.D.L.; Sarhadi, K.; Anicetti, N.; Longwell, P.; et al. An Electroencephalography Connectomic Profile of Posttraumatic Stress Disorder. Am. J. Psychiatry 2020, 177, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Baggio, H.-C.; Segura, B.; Sala-Llonch, R.; Marti, M.-J.; Valldeoriola, F.; Compta, Y.; Tolosa, E.; Junqué, C. Cognitive impairment and resting-state network connectivity in Parkinson’s disease. Hum. Brain Mapp. 2015, 36, 199–212. [Google Scholar] [CrossRef] [PubMed]
- Maidan, I.; Jacob, Y.; Giladi, N.; Hausdorff, J.M.; Mirelman, A. Altered organization of the dorsal attention network is associated with freezing of gait in Parkinson’s disease. Parkinsonism Relat. Disord. 2019, 63, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Nachev, P.; Kennard, C.; Husain, M. Functional role of the supplementary and pre-supplementary motor areas. Nat. Rev. Neurosci. 2008, 9, 856–869. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.H.; Smyser, C.D.; Shimony, J.S. Resting-state fMRI: A review of methods and clinical applications. Am. J. Neuroradiol. 2013, 34, 1866–1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.; Wang, L.; Chen, Y.; Zhao, C.; Li, K.; Chan, P. Changes of functional connectivity of the motor network in the resting state in Parkinson’s disease. Neurosci. Lett. 2009, 460, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Long, X.; Wang, L.; Hallett, M.; Zang, Y.; Li, K.; Chan, P. Functional connectivity of cortical motor areas in the resting state in Parkinson’s disease. Hum. Brain Mapp. 2010, 32, 1443–1457. [Google Scholar] [CrossRef]
- Putcha, D.; Ross, R.; Cronin-Golomb, A.; Janes, A.C.; Stern, C.E. Altered intrinsic functional coupling between core neurocognitive networks in Parkinson’s disease. NeuroImage Clin. 2015, 7, 449–455. [Google Scholar] [CrossRef] [Green Version]
- Bonanni, L.; Perfetti, B.; Bifolchetti, S.; Taylor, J.-P.; Franciotti, R.; Parnetti, L.; Thomas, A.; Onofrj, M. Quantitative electroencephalogram utility in predicting conversion of mild cognitive impairment to dementia with Lewy bodies. Neurobiol. Aging 2014, 36, 434–445. [Google Scholar] [CrossRef] [Green Version]
- Klassen, B.T.; Hentz, J.G.; Shill, H.A.; Driver-Dunckley, E.; Evidente, V.G.H.; Sabbagh, M.N.; Adler, C.H.; Caviness, J.N. Quantitative EEG as a predictive biomarker for Parkinson disease dementia. Neurology 2011, 77, 118–124. [Google Scholar] [CrossRef]
- Caviness, J.N.; Hentz, J.G.; Belden, C.M.; Shill, H.A.; Driver-Dunckley, E.D.; Sabbagh, M.N.; Powell, J.J.; Adler, C.H. Longitudinal EEG Changes Correlate with Cognitive Measure Deterioration in Parkinson’s Disease. J. Park. Dis. 2015, 5, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Caviness, J.N.; Utianski, R.L.; Hentz, J.G.; Beach, T.G.; Dugger, B.N.; Shill, H.A.; Driver-Dunckley, E.D.; Sabbagh, M.N.; Mehta, S.; Adler, C.H. Differential spectral quantitative electroencephalography patterns between control and Parkinson’s disease cohorts. Eur. J. Neurol. 2016, 23, 387–392. [Google Scholar] [CrossRef] [PubMed]
- Olde Dubbelink, K.T.E.; Stoffers, D.; Deijen, J.B.; Twisk, J.W.R.; Stam, C.J.; Berendse, H.W. Cognitive decline in Parkinson’s disease is associated with slowing of resting-state brain activity: A longitudinal study. Neurobiol. Aging 2013, 34, 408–418. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Heldmann, M.; Herrmann, L.; Brüggemann, N.; Münte, T.F. Altered alpha and theta oscillations correlate with sequential working memory in Parkinson’s disease. Brain Commun. 2022, 4, fcac096. [Google Scholar] [CrossRef] [PubMed]
- Başar, E.; Başar-Eroglu, C.; Karakaş, S.; Schürmann, M. Are cognitive processes manifested in event-related gamma, alpha, theta and delta oscillations in the EEG? Neurosci. Lett. 1999, 259, 165–168. [Google Scholar] [CrossRef]
- Gola, M.; Kamiński, J.; Brzezicka, A.; Wróbel, A. Beta band oscillations as a correlate of alertness—Changes in aging. Int. J. Psychophysiol. 2012, 85, 62–67. [Google Scholar] [CrossRef]
- Peláez Suárez, A.A.; Berrillo Batista, S.; Pedroso Ibáñez, I.; Casabona Fernández, E.; Fuentes Campos, M.; Chacón, L.M. Eeg-derived functional connectivity patterns associated with mild cognitive impairment in Parkinson’s disease. Behav. Sci. 2021, 11, 40. [Google Scholar] [CrossRef]
- Goetz, C.G.; Tilley, B.C.; Shaftman, S.R.; Stebbins, G.T.; Fahn, S.; Martinez-Martin, P.; Poewe, W.; Sampaio, C.; Stern, M.B.; Dodel, R.; et al. Movement Disorder Society-Sponsored Revision of the Unified Parkinson’s Disease Rating Scale (MDS-UPDRS): Scale presentation and clinimetric testing results. Mov. Disord. 2008, 23, 2129–2170. [Google Scholar] [CrossRef]
- Nasreddine, Z.S.; Phillips, N.A.; Bédirian, V.; Charbonneau, S.; Whitehead, V.; Collin, I.; Cummings, J.L.; Chertkow, H. The Montreal Cognitive Assessment, MoCA: A Brief Screening Tool For Mild Cognitive Impairment. J Am Geriatr Soc. 2005, 53, 695–699. [Google Scholar] [CrossRef]
- Sánchez-Cubillo, I.; Periáñez, J.; Adrover-Roig, D.; Rodríguez-Sánchez, J.; Ríos-Lago, M.; Tirapu, J.; Barceló, F. Construct validity of the Trail Making Test: Role of task-switching, working memory, inhibition/interference control, and visuomotor abilities. J. Int. Neuropsychol. Soc. 2009, 15, 438–450. [Google Scholar] [CrossRef]
- Delorme, A.; Makeig, S. EEGLAB: An open source toolbox for analysis of single-trial EEG dynamics including independent component analysis. J. Neurosci. Methods 2004, 134, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, M.; Kastner, J.; Wagner, M.; Hawes, S.; Ebersole, J.S. A standardized boundary element method volume conductor model. Clin. Neurophysiol. 2002, 113, 702–712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pascual-Marqui, R.D.; Esslen, M.; Kochi, K.; Lehmann, D. Functional imaging with low-resolution brain electromagnetic tomography (LORETA): A review. Methods Find. Exp. Clin. Pharmacol. 2002, 24 (Suppl. SC), 91–95. [Google Scholar] [PubMed]
- Pascual-Marqui, R.D. Standardized low-resolution brain electromagnetic tomography (sLORETA): Technical details. Methods Find. Exp. Clin. Pharmacol. 2002, 24 (Suppl. SD), 5–12. [Google Scholar]
- Laird, A.R.; Fox, P.M.; Eickhoff, S.B.; Turner, J.A.; Ray, K.L.; McKay, D.R.; Glahn, D.C.; Beckmann, C.F.; Smith, S.M. Behavioral Interpretations of Intrinsic Connectivity Networks. J. Cogn. Neurosci. 2011, 23, 4022–4037. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.M.; Fox, P.T.; Miller, K.L.; Glahn, D.C.; Mackay, C.E.; Filippini, N.; Watkins, K.; Toro, R.; Laird, A.; Beckmann, C.F. Correspondence of the brain’s functional architecture during activation and rest. Proc. Natl. Acad. Sci. USA 2009, 106, 13040–13045. [Google Scholar] [CrossRef] [Green Version]
- Vergotte, G.; Torre, K.; Chirumamilla, V.C.; Anwar, A.R.; Groppa, S.; Perrey, S.; Muthuraman, M. Dynamics of the human brain network revealed by time-frequency effective connectivity in fNIRS. Biomed. Opt. Express 2017, 8, 5326–5341. [Google Scholar] [CrossRef] [Green Version]
- Haufe, S.; Nikulin, V.V.; Müller, K.-R.; Nolte, G. A critical assessment of connectivity measures for EEG data: A simulation study. Neuroimage 2013, 64, 120–133. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Vossel, S.; Geng, J.J.; Fink, G.R. Dorsal and ventral attention systems: Distinct neural circuits but collaborative roles. Neuroscientist 2014, 20, 150–159. [Google Scholar] [CrossRef]
- Corbetta, M.; Shulman, G.L. Control of goal-directed and stimulus-driven attention in the brain. Nat. Rev. Neurosci. 2002, 3, 201–215. [Google Scholar] [CrossRef] [PubMed]
- Brown, C.A.; Schmitt, F.A.; Smith, C.D.; Gold, B.T. Distinct patterns of default mode and executive control network circuitry contribute to present and future executive function in older adults. Neuroimage 2019, 195, 320–332. [Google Scholar] [CrossRef]
- Cai, S.; Peng, Y.; Chong, T.; Zhang, Y.; Von Deneen, K.M.; Huang, L.; Cai, Y.P.S. Differentiated Effective Connectivity Patterns of the Executive Control Network in Progressive MCI: A Potential Biomarker for Predicting AD. Curr. Alzheimer Res. 2017, 14, 937–950. [Google Scholar] [CrossRef] [PubMed]
- Chan, R.C.K.; Shum, D.; Toulopoulou, T.; Chen, E.Y. Assessment of executive functions: Review of instruments and identification of critical issues. Arch. Clin. Neuropsychol. 2008, 23, 201–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, W.; Qiu, C.; Jiang, X.; Shen, B.; Zhang, L.; Liu, W.; Zhang, W.; Chen, J. Can the Executive Control Network be Used to Diagnose Parkinson’s Disease and as an Efficacy Indicator of Deep Brain Stimulation? Parkinson’s Dis. 2020, 2020, 6348102. [Google Scholar] [CrossRef] [Green Version]
- Biswal, B.; Yetkin, F.Z.; Haughton, V.M.; Hyde, J.S. Functional connectivity in the motor cortex of resting human brain using echo-planar mri. Magn. Reson. Med. 1995, 34, 537–541. [Google Scholar] [CrossRef]
- De Luca, M.; Smith, S.; de Stefano, N.; Federico, A.; Matthews, P.M. Blood oxygenation level dependent contrast resting state networks are relevant to functional activity in the neocortical sensorimotor system. Exp. Brain Res. 2005, 167, 587–594. [Google Scholar] [CrossRef]
- Caspers, J.; Rubbert, C.; Eickhoff, S.B.; Hoffstaedter, F.; Südmeyer, M.; Hartmann, C.J.; Sigl, B.; Teichert, N.; Aissa, J.; Turowski, B.; et al. Within- and across-network alterations of the sensorimotor network in Parkinson’s disease. Neuroradiology 2021, 63, 2073–2085. [Google Scholar] [CrossRef]
- Lange, F.; Brückner, C.; Knebel, A.; Seer, C.; Kopp, B. Executive dysfunction in Parkinson’s disease: A meta-analysis on the Wisconsin Card Sorting Test literature. Neurosci. Biobehav. Rev. 2018, 93, 38–56. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HC (n = 28) | PD (n = 29) | p Values | |
|---|---|---|---|
| Age (years) | 60.82 ± 7.47 | 64.36 ± 7.11 | 0.075 |
| Gender (male/female) | 13/15 | 16/13 | 0.518 |
| MoCA | 27.44 ± 1.97 | 26.77 ± 2.1 | 0.104 |
| CTT (part B) | 84.85 ± 29.66 | 101.56 ± 57.03 | 0.186 |
| MDS-UPDRS motor | 1.38 ± 1.74 | 18.89 ± 10.05 | <0.001 |
| Disease duration (years) | na | 2.8 ± 0.28 | na |
| LEDD (mg) | na | 183.90 ± 249.0 | na |
| HY score (HY1/HY2) | na | 10/19 | na |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leviashvili, S.; Ezra, Y.; Droby, A.; Ding, H.; Groppa, S.; Mirelman, A.; Muthuraman, M.; Maidan, I. EEG-Based Mapping of Resting-State Functional Brain Networks in Patients with Parkinson’s Disease. Biomimetics 2022, 7, 231. https://doi.org/10.3390/biomimetics7040231
Leviashvili S, Ezra Y, Droby A, Ding H, Groppa S, Mirelman A, Muthuraman M, Maidan I. EEG-Based Mapping of Resting-State Functional Brain Networks in Patients with Parkinson’s Disease. Biomimetics. 2022; 7(4):231. https://doi.org/10.3390/biomimetics7040231
Chicago/Turabian StyleLeviashvili, Sarah, Yael Ezra, Amgad Droby, Hao Ding, Sergiu Groppa, Anat Mirelman, Muthuraman Muthuraman, and Inbal Maidan. 2022. "EEG-Based Mapping of Resting-State Functional Brain Networks in Patients with Parkinson’s Disease" Biomimetics 7, no. 4: 231. https://doi.org/10.3390/biomimetics7040231