
Wnt/β-Catenin Signaling Inhibits Osteogenic Differentiation in Human Periodontal Ligament Fibroblasts

Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture
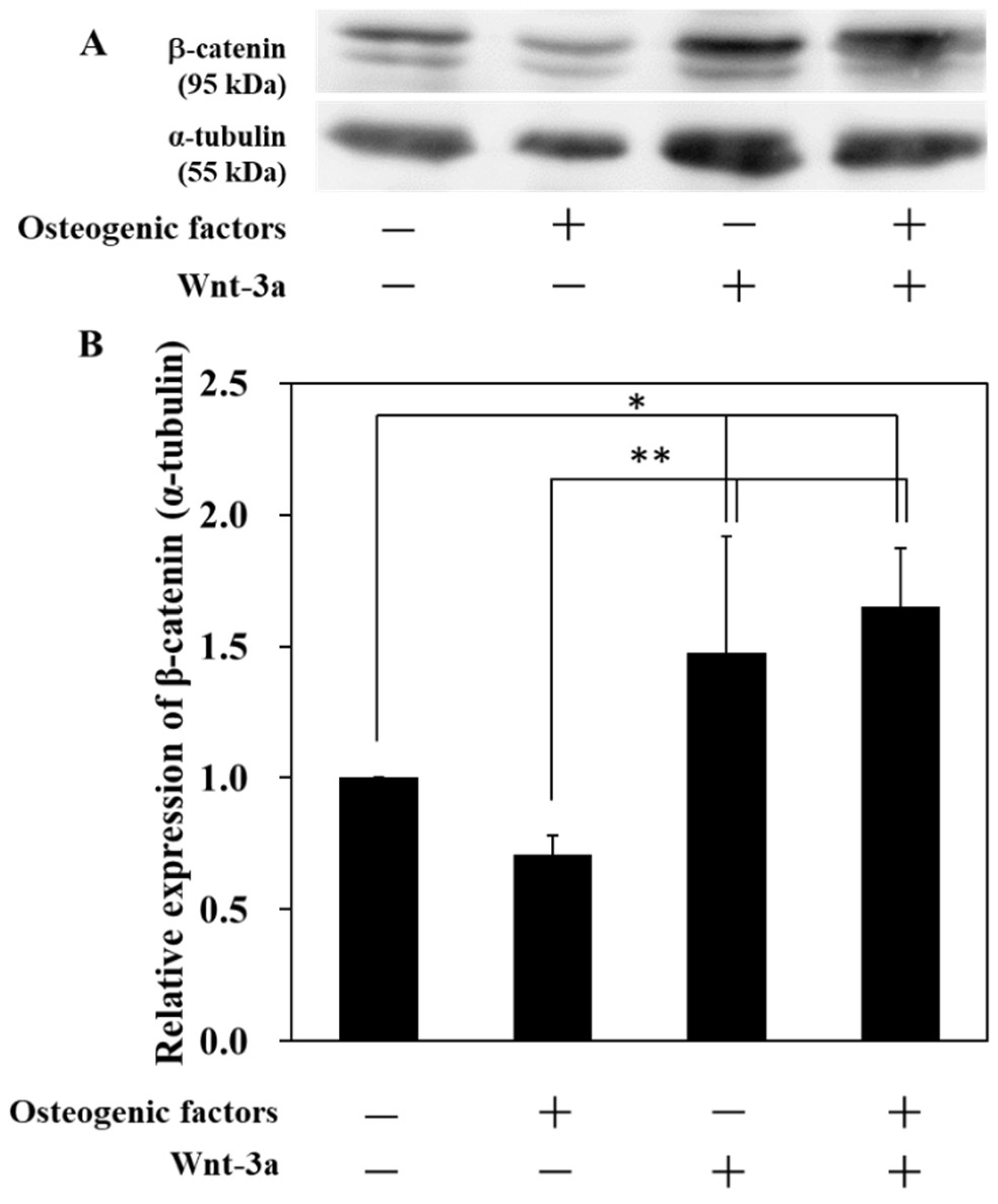
2.3. Western Blotting
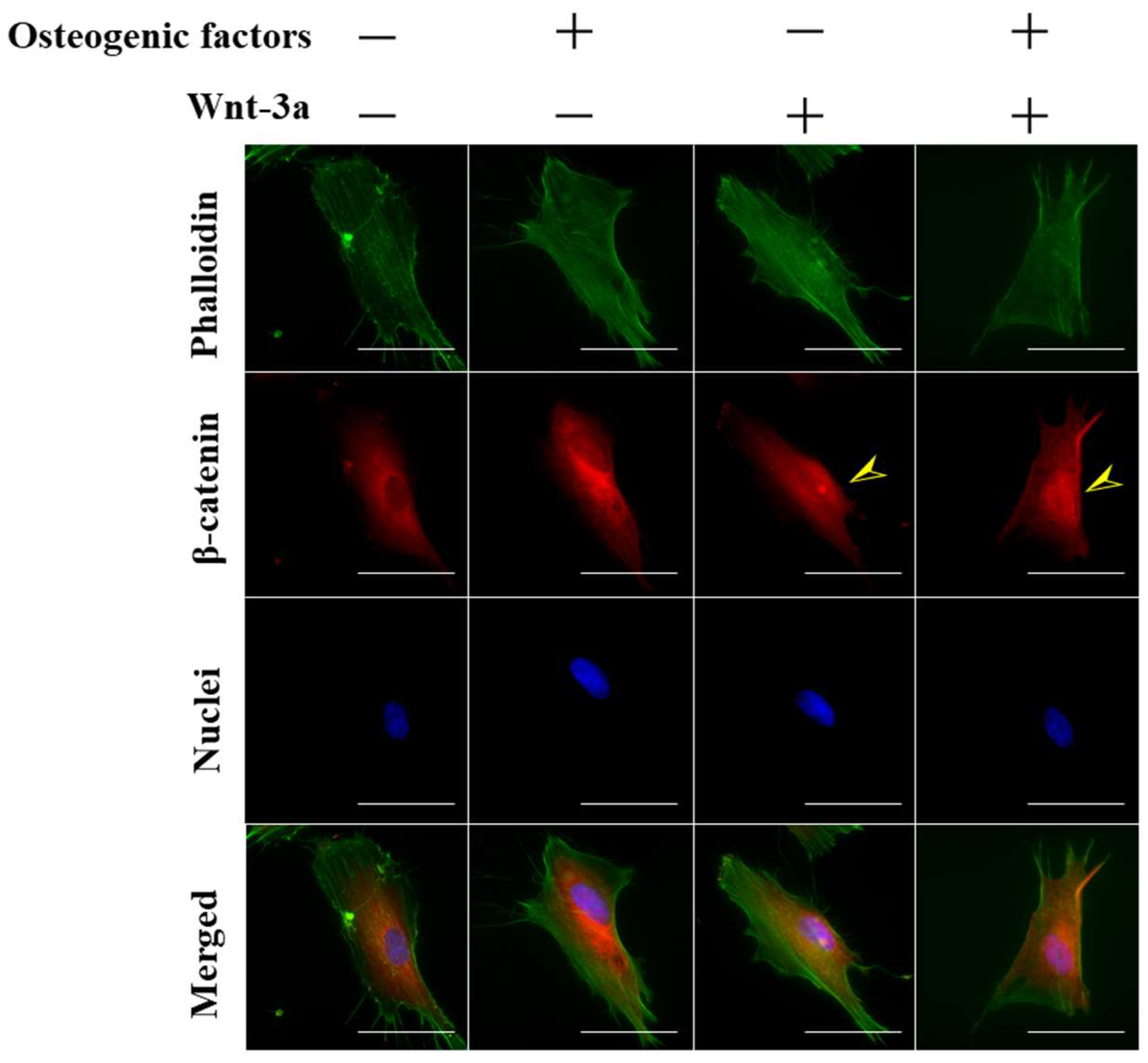
2.4. Immunofluorescence Microscopy
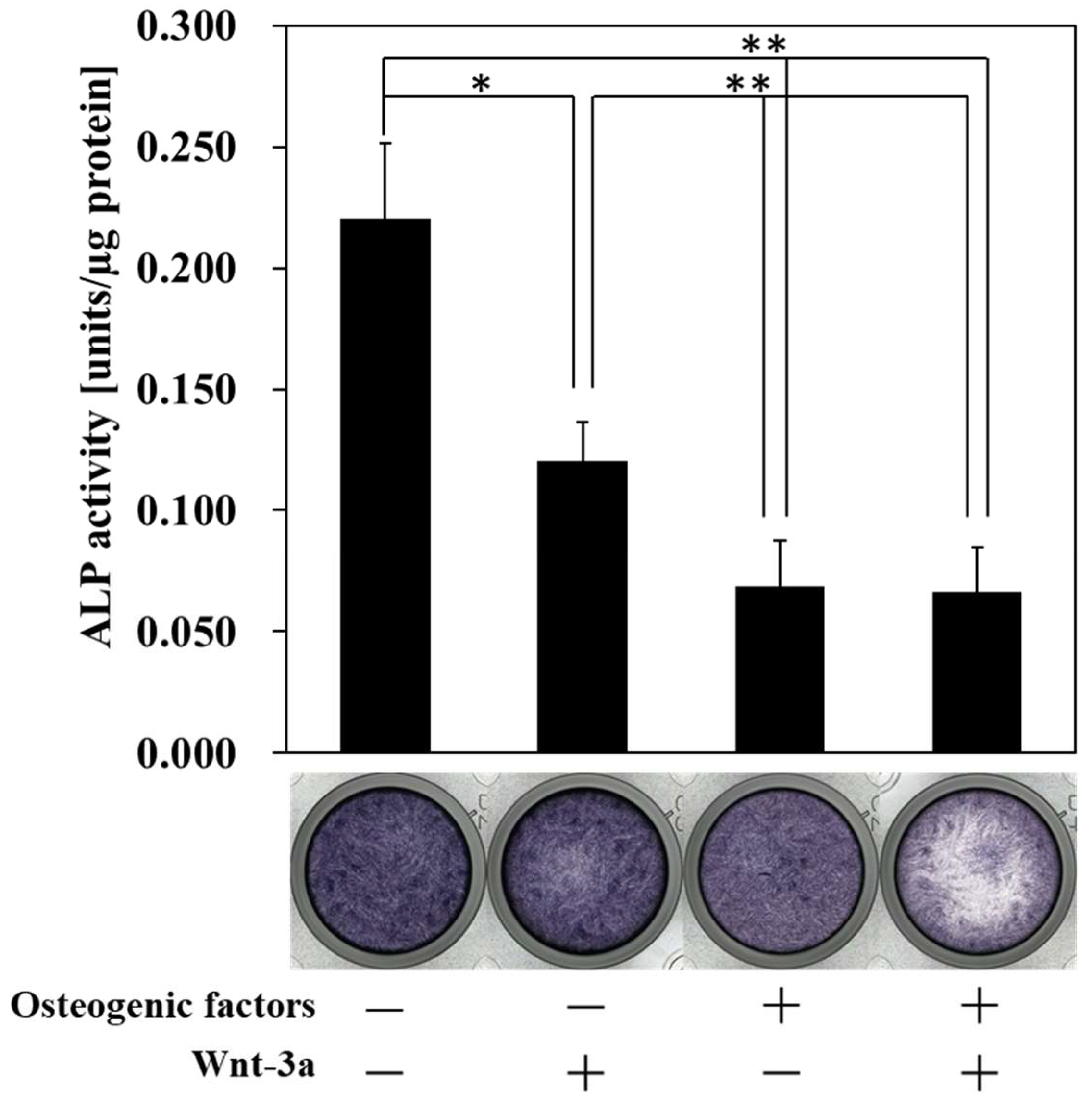
2.5. Alkaline Phosphatase Staining
2.6. Alkaline Phosphatase Activity
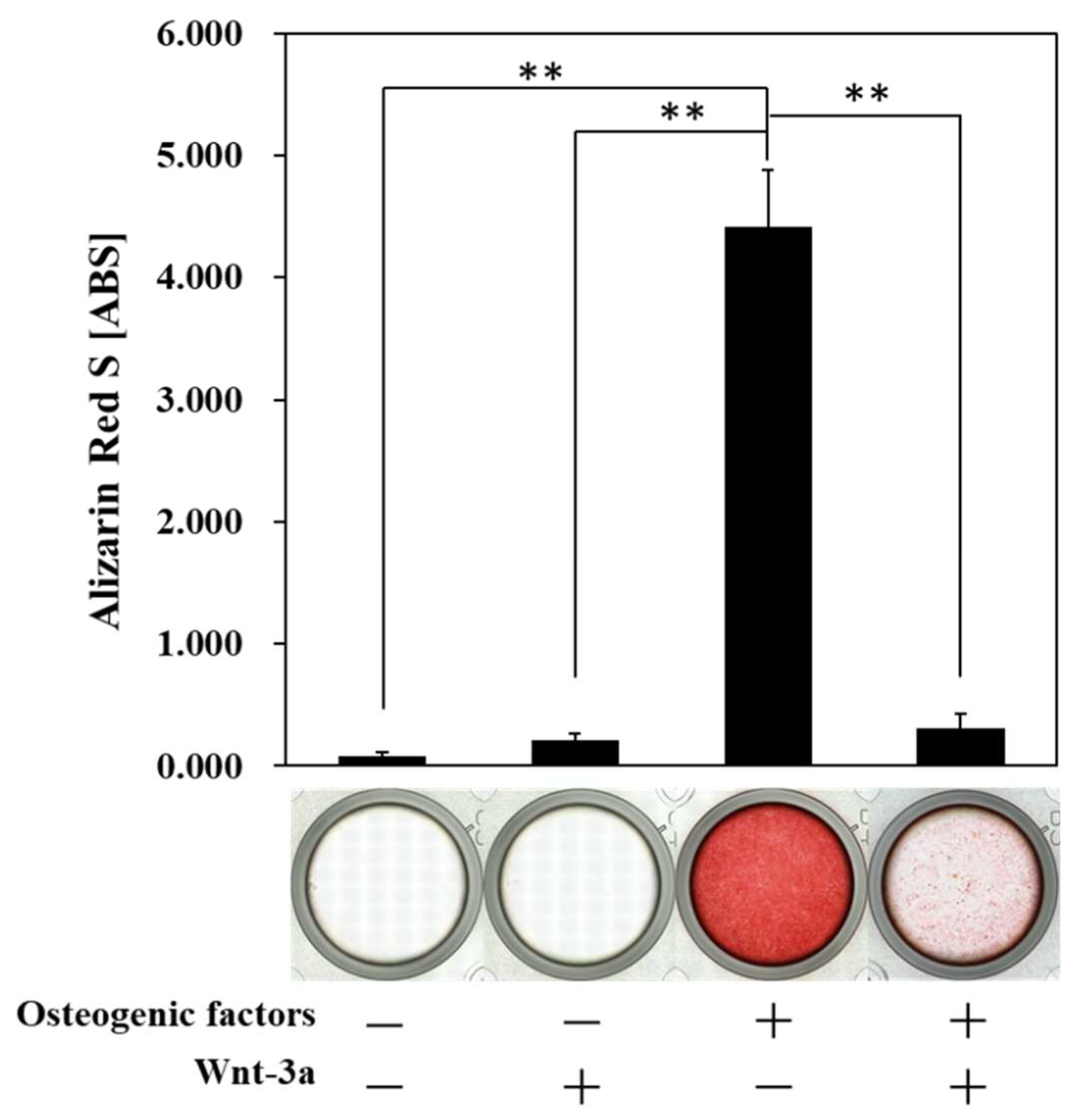
2.7. Alizarin Red S Staining
2.8. Quantitative Real-Time PCR
2.9. Statistical Analysis
3. Results
3.1. Activation of Wnt/β-Catenin Signaling by Wnt-3a Stimulation in HPLFs
3.2. Effect of Wnt/β-Catenin Signaling on the Production of ALP, a Marker of Early Osteogenic Differentiation
3.3. Effect of Wnt/β-Catenin Signaling on the Formation of Calcified Nodules
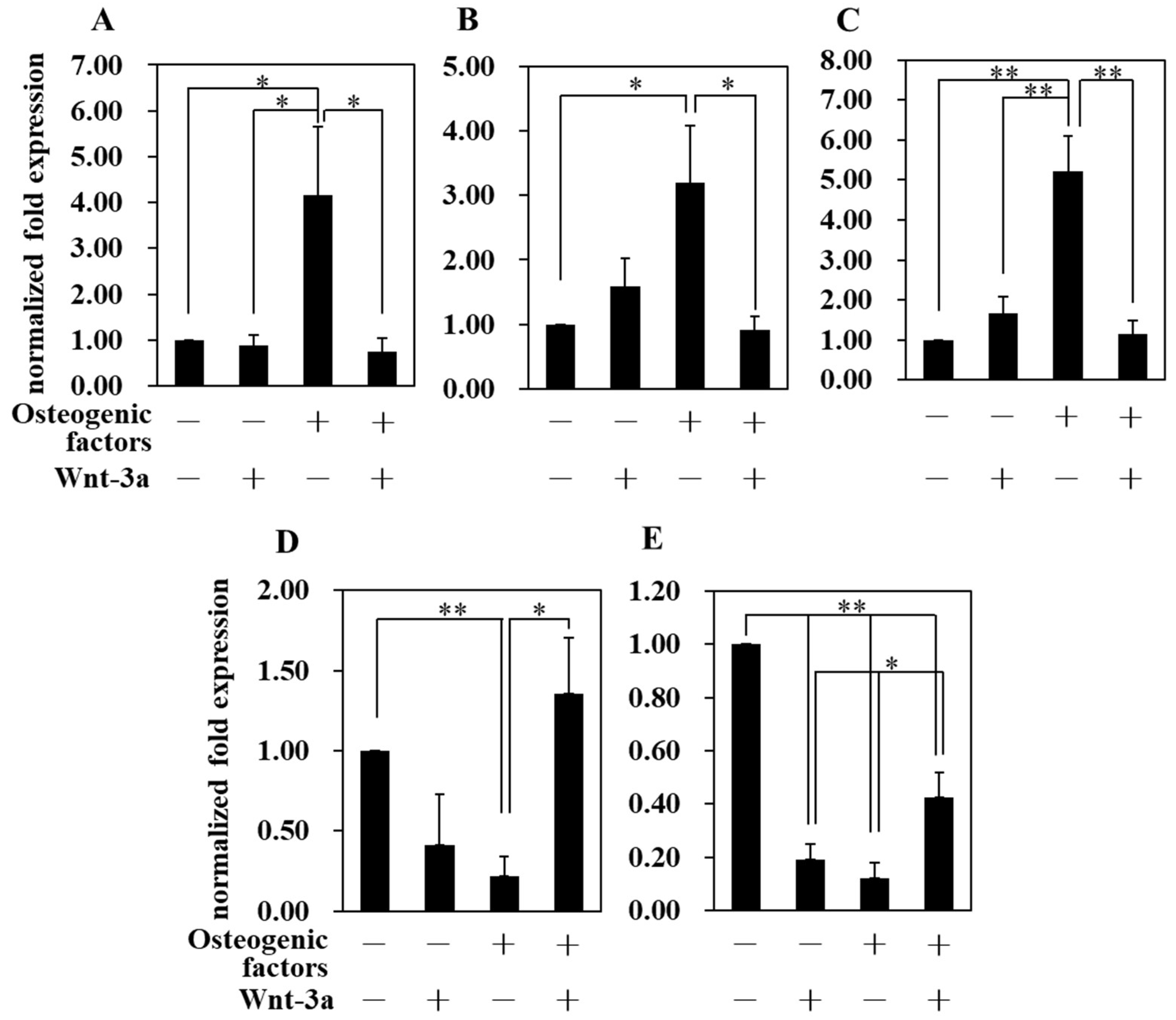
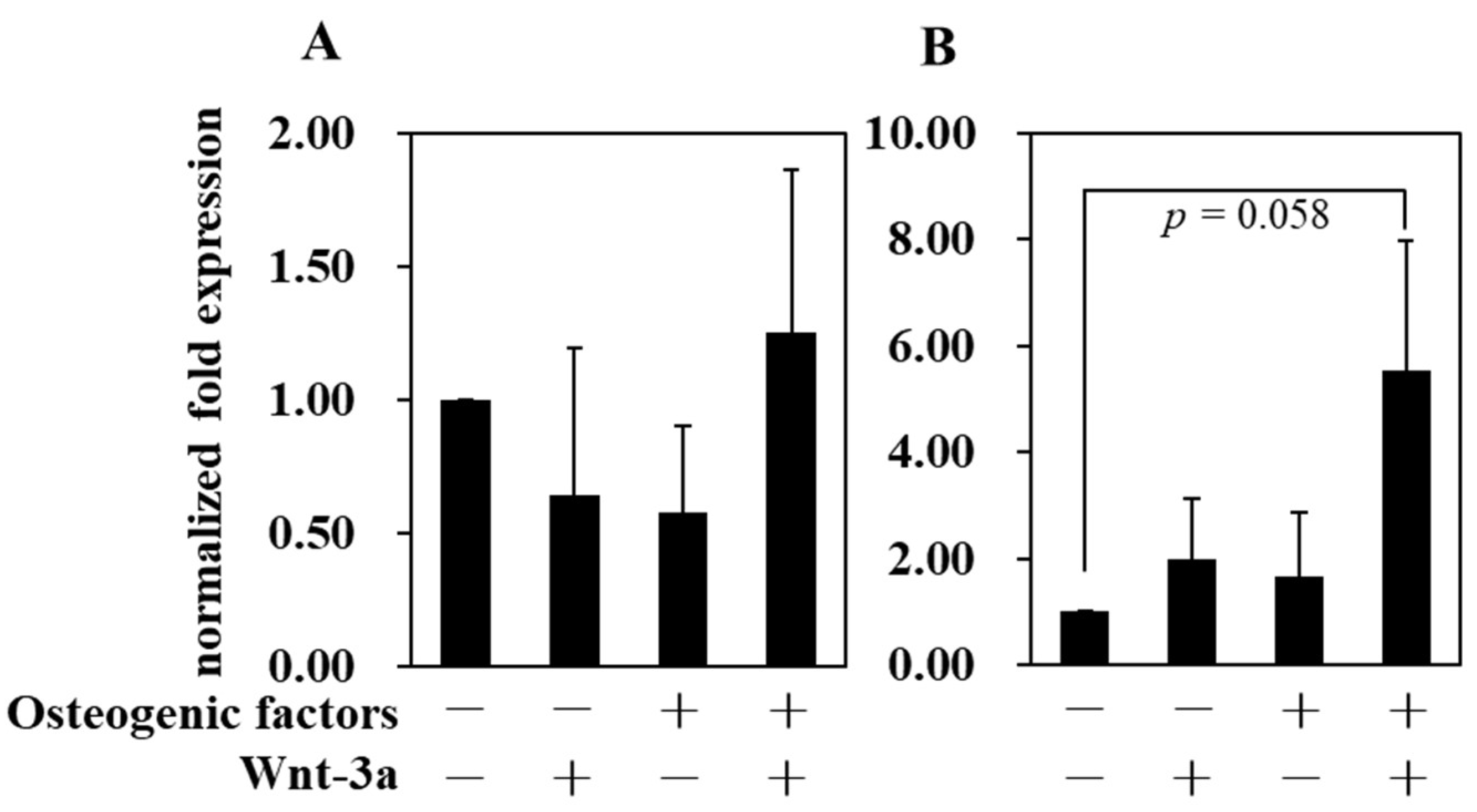
3.4. Effect of Wnt/β-Catenin Signaling on Gene Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Maeda, T.; Hayashi, S. Morphological basis of periodontal nerve endings. Kaibogaku Zasshi J. Anat. 2000, 75, 377–380. [Google Scholar]
- Li, M.; Zhang, C.; Yang, Y. Effects of mechanical forces on osteogenesis and osteoclastogenesis in human periodontal ligament fibroblasts: A systematic review of in vitro studies. Bone Jt. Res. 2019, 8, 19–31. [Google Scholar] [CrossRef]
- Raju, R.; Oshima, M.; Inoue, M.; Morita, T.; Huijiao, Y.; Waskitho, A.; Baba, O.; Inoue, M.; Matsuka, Y. Three-dimensional periodontal tissue regeneration using a bone-ligament complex cell sheet. Sci. Rep. 2020, 10, 1656. [Google Scholar] [CrossRef] [Green Version]
- Bottino, M.C.; Thomas, V.; Schmidt, G.; Vohra, Y.K.; Chu, T.-M.G.; Kowolik, M.J.; Janowski, G.M. Recent advances in the development of GTR/GBR membranes for periodontal regeneration—A materials perspective. Dent. Mater. 2012, 28, 703–721. [Google Scholar] [CrossRef]
- Seo, B.-M.; Miura, M.; Gronthos, S.; Bartold, P.M.; Batouli, S.; Brahim, J.; Young, M.; Robey, P.G.; Wang, C.Y.; Shi, S. Investigation of multipotent postnatal stem cells from human periodontal ligament. Lancet 2004, 364, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Lozano, F.J.; Bueno, C.R.; Insausti, C.L.; Meseguer-Olmo, L.; Ramírez, M.C.; Blanquer, M.B.; Marín, N.; Martinez, S.; Moraleda, J.M. Mesenchymal stem cells derived from dental tissues. Int. Endod. J. 2011, 44, 800–806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Ruan, J.; Weir, M.D.; Ren, K.; Schneider, A.; Wang, P.; Oates, T.W.; Chang, X.; Xu, H.H.K. Periodontal Bone-Ligament-Cementum Regeneration via Scaffolds and Stem Cells. Cells 2019, 8, 537. [Google Scholar] [CrossRef] [Green Version]
- Yu, B.; Li, Q.; Zhou, M. LPS-induced upregulation of the TLR4 signaling pathway inhibits osteogenic differentiation of human periodontal ligament stem cells under inflammatory conditions. Int. J. Mol. Med. 2019, 43, 2341–2351. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhang, F.; Zhang, N.; Geng, X.; Meng, C.; Wang, X.; Yang, Y. Osteogenic capacity and cytotherapeutic potential of periodontal ligament cells for periodontal regeneration in vitro and in vivo. PeerJ 2019, 7, e6589. [Google Scholar] [CrossRef] [Green Version]
- Koda, N.; Sato, T.; Shinohara, M.; Ichinose, S.; Ito, Y.; Nakamichi, R.; Kayama, T.; Kataoka, K.; Suzuki, H.; Moriyama, K.; et al. The transcription factor mohawk homeobox regulates homeostasis of the periodontal ligament. Development 2017, 144, 313–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takimoto, A.; Kawatsu, M.; Yoshimoto, Y.; Kawamoto, T.; Seiryu, M.; Takano-Yamamoto, T.; Hiraki, Y.; Shukunami, C. Scleraxis and osterix antagonistically regulate tensile force-responsive remodeling of the periodontal ligament and alveolar bone. Development 2015, 142, 787–796. [Google Scholar] [CrossRef]
- Hasegawa, D.; Wada, N.; Maeda, H.; Yoshida, S.; Mitarai, H.; Tomokiyo, A.; Monnouchi, S.; Hamano, S.; Yuda, A.; Akamine, A. Wnt5a Induces Collagen Production by Human Periodontal Ligament Cells Through TGFβ1-Mediated Upregulation of Periostin Expression. J. Cell. Physiol. 2015, 230, 2647–2660. [Google Scholar] [CrossRef]
- Norris, R.A.; Damon, B.; Mironov, V.; Kasyanov, V.; Ramamurthi, A.; Moreno-Rodriguez, R.; Trusk, T.; Potts, J.D.; Goodwin, R.L.; Davis, J.; et al. Periostin regulates collagen fibrillogenesis and the biomechanical properties of connective tissues. J. Cell. Biochem. 2007, 101, 695–711. [Google Scholar] [CrossRef] [Green Version]
- Kishimoto, Y.; Ohkawara, B.; Sakai, T.; Ito, M.; Masuda, A.; Ishiguro, N.; Shukunami, C.; Docheva, D.; Ohno, K. Wnt/β-catenin signaling suppresses expressions of Scx, Mkx, and Tnmd in tendon-derived cells. PLoS ONE 2017, 12, e0182051. [Google Scholar] [CrossRef] [Green Version]
- Qiu, W.; Chen, L.; Kassem, M. Activation of non-canonical Wnt/JNK pathway by Wnt3a is associated with differentiation fate determination of human bone marrow stromal (mesenchymal) stem cells. Biochem. Biophys. Res. Commun. 2011, 413, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Miyabara, S.; Yuda, Y.; Kasashima, Y.; Kuwano, A.; Arai, K. Regulation of Tenomodulin Expression Via Wnt/β-catenin Signaling in Equine Bone Marrow-derived Mesenchymal Stem Cells. J. Equine Sci. 2014, 25, 7–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Ge, Y.; Chen, G.; Yan, Z.; Yu, M.; Feng, L.; Jiang, Z.; Guo, W.; Tian, W. Hertwig’s epithelial root sheath cells regulate osteogenic differentiation of dental follicle cells through the Wnt pathway. Bone 2014, 63, 158–165. [Google Scholar] [CrossRef] [PubMed]
- Nemoto, E.; Koshikawa, Y.; Kanaya, S.; Tsuchiya, M.; Tamura, M.; Somerman, M.J.; Shimauchi, H. Wnt signaling inhibits cementoblast differentiation and promotes proliferation. Bone 2009, 44, 805–812. [Google Scholar] [CrossRef]
- Krieghoff, E.; Behrens, J.; Mayr, B. Nucleo-cytoplasmic distribution of β-catenin is regulated by retention. J. Cell Sci. 2006, 119, 1453–1463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, W.; Cao, Y.; Yang, H.; Han, N.; Zhu, X.; Fan, Z.; Du, J.; Zhang, F. CB1 enhanced the osteo/dentinogenic differentiation ability of periodontal ligament stem cells via p38 MAPK and JNK in an inflammatory environment. Cell Prolif. 2019, 52, e12691. [Google Scholar] [CrossRef]
- Ibarra, B.; Machen, C.; Atit, R. Wnt-Dependent Activation of ERK Mediates Repression of Chondrocyte Fate during Calvarial Development. J. Dev. Biol. 2021, 9, 23. [Google Scholar] [CrossRef]
- Brunt, K.R.; Zhang, Y.; Mihic, A.; Li, M.; Li, S.-H.; Xue, P.; Zhang, W.; Basmaji, S.; Tsang, K.; Weisel, R.D.; et al. Role of WNT/β-Catenin Signaling in Rejuvenating Myogenic Differentiation of Aged Mesenchymal Stem Cells from Cardiac Patients. Am. J. Pathol. 2012, 181, 2067–2078. [Google Scholar] [CrossRef] [PubMed]
- Hessle, L.; Johnson, K.A.; Anderson, H.C.; Narisawa, S.; Sali, A.; Goding, J.W.; Terkeltaub, R.; Millán, J.L. Tissue-nonspecific alkaline phosphatase and plasma cell membrane glycoprotein-1 are central antagonistic regulators of bone mineralization. Proc. Natl. Acad. Sci. USA 2002, 99, 9445–9449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhang, Y.; Choukroun, J.; Ghanaati, S.; Miron, R.J. Effects of an injectable platelet-rich fibrin on osteoblast behavior and bone tissue formation in comparison to platelet-rich plasma. Platelets 2018, 29, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Nam, S.W.; Kim, S.H.; Han, Y. Discovery and development of berberine derivatives as stimulants of osteoblast differentiation. Biochem. Biophys. Res. Commun. 2020, 527, 110–116. [Google Scholar] [CrossRef] [PubMed]
- Sudo, H.; Kodama, H.A.; Amagai, Y.; Yamamoto, S.; Kasai, S. In vitro differentiation and calcification in a new clonal osteogenic cell line derived from newborn mouse calvaria. J. Cell Biol. 1983, 96, 191–198. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroiwa, M.; Chihara, K.; Higashi, S. Electron microscopic studies on Sharpey’s fibers in the alveolar bone of rat molars. Kaibogaku Zasshi J. Anat. 1994, 69, 776–782. [Google Scholar]
- Choi, J.W.; Arai, C.; Ishikawa, M.; Shimoda, S.; Nakamura, Y. Fiber system degradation, and periostin and connective tissue growth factor level reduction, in the periodontal ligament of teeth in the absence of masticatory load. J. Periodontal Res. 2011, 46, 513–521. [Google Scholar] [CrossRef]
- Lee, S.-J.; Lee, E.-H.; Park, S.-Y.; Kim, J.-E. Induction of fibrillin-2 and periostin expression in Osterix-knockdown MC3T3-E1 cells. Gene 2017, 596, 123–129. [Google Scholar] [CrossRef]
- Wilde, J.; Yokozeki, M.; Terai, K.; Kudo, A.; Moriyama, K. The divergent expression of periostin mRNA in the periodontal ligament during experimental tooth movement. Cell Tissue Res. 2003, 312, 345–351. [Google Scholar] [CrossRef]
- Yokoya, F.; Imamoto, N.; Tachibana, T.; Yoneda, Y. β-Catenin Can Be Transported into the Nucleus in a Ran-unassisted Manner. Mol. Biol. Cell 1999, 10, 1119–1131. [Google Scholar] [CrossRef] [PubMed]
- Koike, M.; Kose, S.; Furuta, M.; Taniguchi, N.; Yokoya, F.; Yoneda, Y.; Imamoto, N. β-Catenin Shows an Overlapping Sequence Requirement but Distinct Molecular Interactions for Its Bidirectional Passage through Nuclear Pores. J. Biol. Chem. 2004, 279, 34038–34047. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.H.; Liu, B.; Cheng, D.; Williams, B.O.; Mah, S.J.; Helms, J.A. Wnt signaling regulates homeostasis of the periodontal ligament. J. Periodontal Res. 2014, 49, 751–759. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, L.; Zhang, Y.; Li, D.; Zhang, W.; Zhang, D.; Xu, X. Analyses of key mRNAs and lncRNAs for different osteo-differentiation potentials of periodontal ligament stem cell and gingival mesenchymal stem cell. J. Cell. Mol. Med. 2021, 25, 6217–6231. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Hutmacher, D.W.; Sae-Lim, V.; Zhou, Z.; Woodruff, M.; Lim, T.M. Osteogenic and Adipogenic Induction Potential of Human Periodontal Cells. J. Periodontol. 2008, 79, 525–534. [Google Scholar] [CrossRef]
- Zhang, J.; Zhang, W.; Dai, J.; Wang, X.; Shen, S.G. Overexpression of Dlx2 enhances osteogenic differentiation of BMSCs and MC3T3-E1 cells via direct upregulation of Osteocalcin and Alp. Int. J. Oral Sci. 2019, 11, 12. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Huang, Y.; Tan, L. Downregulation of lncRNA DANCR promotes osteogenic differentiation of periodontal ligament stem cells. BMC Dev. Biol. 2020, 20, 2. [Google Scholar] [CrossRef] [Green Version]
- Soda, M.; Saitoh, I.; Murakami, T.; Inada, E.; Iwase, Y.; Noguchi, H.; Shibasaki, S.; Kurosawa, M.; Sawami, T.; Terunuma, M.; et al. Repeated human deciduous tooth-derived dental pulp cell reprogramming factor transfection yields multipotent intermediate cells with enhanced iPS cell formation capability. Sci. Rep. 2019, 9, 1490. [Google Scholar] [CrossRef] [Green Version]
- Zhang, P.; Chang, W.-H.; Fong, B.; Gao, F.; Liu, C.; Al Alam, D.; Bellusci, S.; Lu, W. Regulation of Induced Pluripotent Stem (iPS) Cell Induction by Wnt/β-Catenin Signaling. J. Biol. Chem. 2014, 289, 9221–9232. [Google Scholar] [CrossRef] [Green Version]
- Hong, H.; Pischon, N.; Santana, R.; Palamakumbura, A.H.; Chase, H.B.; Gantz, D.; Guo, Y.; Uzel, M.I.; Ma, D.; Trackman, P.C. A role for lysyl oxidase regulation in the control of normal collagen deposition in differentiating osteoblast cultures. J. Cell. Physiol. 2004, 200, 53–62. [Google Scholar] [CrossRef]
- Aaron, J.E. Periosteal Sharpey’s fibers: A novel bone matrix regulatory system? Front. Endocrinol. 2012, 3, 98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuura, T.; Tokutomi, K.; Sasaki, M.; Katafuchi, M.; Mizumachi, E.; Sato, H. Distinct Characteristics of Mandibular Bone Collagen Relative to Long Bone Collagen: Relevance to Clinical Dentistry. BioMed Res. Int. 2014, 2014, 769414. [Google Scholar] [CrossRef] [PubMed]
- Almalki, S.G.; Agrawal, D.K. Key transcription factors in the differentiation of mesenchymal stem cells. Differentiation 2016, 92, 41–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carvalho, M.S.; Cabral, J.M.; da Silva, C.L.; Vashishth, D. Synergistic effect of extracellularly supplemented osteopontin and osteocalcin on stem cell proliferation, osteogenic differentiation, and angiogenic properties. J. Cell. Biochem. 2019, 120, 6555–6569. [Google Scholar] [CrossRef]
- Komori, T.; Yagi, H.; Nomura, S.; Yamaguchi, A.; Sasaki, K.; Deguchi, K.; Shimizu, Y.; Bronson, R.T.; Gao, Y.H.; Inada, M.; et al. Targeted Disruption of Cbfa1 Results in a Complete Lack of Bone Formation owing to Maturational Arrest of Osteoblasts. Cell 1997, 89, 755–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.N.; Hwang, H.-S.; Oh, S.-H.; Roshanzadeh, A.; Kim, J.-W.; Song, J.H.; Kim, E.-S.; Koh, J.-T. Elevated extracellular calcium ions promote proliferation and migration of mesenchymal stem cells via increasing osteopontin expression. Exp. Mol. Med. 2018, 50, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Rittling, S.R.; Matsumoto, H.N.; McKee, M.D.; Nanci, A.; Novick, K.E.; Kowalski, A.J.; Noda, M.; Denhardt, D.T.; An, X.-R. Mice Lacking Osteopontin Show Normal Development and Bone Structure but Display Altered Osteoclast Formation In Vitro. J. Bone Miner. Res. 1998, 13, 1101–1111. [Google Scholar] [CrossRef]
- Bailey, S.; Karsenty, G.; Gundberg, C.; Vashishth, D. Osteocalcin and osteopontin influence bone morphology and mechanical properties. Ann. N. Y. Acad. Sci. 2017, 1409, 79–84. [Google Scholar] [CrossRef]
- Uribe-Etxebarria, V.; García-Gallastegui, P.; Pérez-Garrastachu, M.; Casado-Andrés, M.; Irastorza, I.; Unda, F.; Ibarretxe, G.; Subirán, N. Wnt-3a Induces Epigenetic Remodeling in Human Dental Pulp Stem Cells. Cells 2020, 9, 652. [Google Scholar] [CrossRef] [Green Version]
- Rolph, D.N.; Deb, M.; Kanji, S.; Greene, C.J.; Das, M.; Joseph, M.; Aggarwal, R.; Leblebicioglu, B.; Das, H. Ferutinin directs dental pulp-derived stem cells towards the osteogenic lineage by epigenetically regulating canonical Wnt signaling. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165314. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, J.; Deng, F.; Huang, E.; Yan, Z.; Wang, Z.; Deng, Y.; Zhang, Q.; Zhang, Z.; Ye, J.; et al. Canonical Wnt signaling acts synergistically on BMP9-induced osteo/odontoblastic differentiation of stem cells of dental apical papilla (SCAPs). Biomaterials 2015, 39, 145–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arzate, H.; Zeichner-David, M.; Mercado-Celis, G. Cementum proteins: Role in cementogenesis, biomineralization, periodontium formation and regeneration. Periodontology 2000 2015, 67, 211–233. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-H.; Zhang, X.-J.; Dang, N.-N.; Ma, Z.-F.; Xu, L.; Wu, J.-J.; Sun, Y.-J.; Duan, Y.-Z.; Lin, Z.; Jin, Y. Apical tooth germ cell-conditioned medium enhances the differentiation of periodontal ligament stem cells into cementum/periodontal ligament-like tissues. J. Periodontal Res. 2009, 44, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Gauthier, P.; Yu, Z.; Tran, Q.T.; Bhatti, F.-U.; Zhu, X.; Huang, G.T.-J. Cementogenic genes in human periodontal ligament stem cells are downregulated in response to osteogenic stimulation while upregulated by vitamin C treatment. Cell Tissue Res. 2017, 368, 79–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komaki, M.; Iwasaki, K.; Arzate, H.; Narayanan, A.S.; Izumi, Y.; Morita, I. Cementum protein 1 (CEMP1) induces a cementoblastic phenotype and reduces osteoblastic differentiation in periodontal ligament cells. J. Cell. Physiol. 2012, 227, 649–657. [Google Scholar] [CrossRef] [PubMed]
- Kadokura, H.; Yamazaki, T.; Masuda, Y.; Kato, Y.; Hasegawa, A.; Sakagami, H.; Yokose, S. Establishment of a Primary Culture System of Human Periodontal Ligament Cells that Differentiate into Cementum Protein 1-expressing Cementoblast-like Cells. In Vivo 2019, 33, 349–352. [Google Scholar] [CrossRef] [Green Version]
- Du, J.; Li, M. Functions of Periostin in dental tissues and its role in periodontal tissues’ regeneration. Cell. Mol. Life Sci. 2017, 74, 4279–4286. [Google Scholar] [CrossRef]
- Afanador, E.; Yokozeki, M.; Oba, Y.; Kitase, Y.; Takahashi, T.; Kudo, A.; Moriyama, K. Messenger RNA expression of periostin and Twist transiently decrease by occlusal hypofunction in mouse periodontal ligament. Arch. Oral Biol. 2005, 50, 1023–1031. [Google Scholar] [CrossRef]
- Xu, H.-Y.; Nie, E.-M.; Deng, G.; Lai, L.-Z.; Sun, F.-Y.; Tian, H.; Fang, F.-C.; Zou, Y.-G.; Wu, B.-L.; Ou-Yang, J. Periostin is essential for periodontal ligament remodeling during orthodontic treatment. Mol. Med. Rep. 2017, 15, 1800–1806. [Google Scholar] [CrossRef] [Green Version]
- Yoshimoto, Y.; Takimoto, A.; Watanabe, H.; Hiraki, Y.; Kondoh, G.; Shukunami, C. Scleraxis is required for maturation of tissue domains for proper integration of the musculoskeletal system. Sci. Rep. 2017, 7, 45010. [Google Scholar] [CrossRef] [Green Version]
- Huang, A.H.; Watson, S.S.; Wang, L.; Baker, B.M.; Akiyama, H.; Brigande, J.V.; Schweitzer, R. Requirement for Scleraxis in the recruitment of mesenchymal progenitors during embryonic tendon elongation. Development 2019, 146, dev182782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawatsu, M.; Takeshita, N.; Takimoto, A.; Yoshimoto, Y.; Seiryu, M.; Ito, A.; Kimura, S.; Kawamoto, T.; Hiraki, Y.; Shukunami, C.; et al. Scleraxis upregulated by transforming growth factor-β1 signaling inhibits tension-induced osteoblast differentiation of priodontal ligament cells via ephrin A2. Bone 2021, 149, 115969. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Abbreviation | −/− (DMEM only) | −/+ | +/− | +/+ |
| Osteogenic factors | − | − | + | + |
| Wnt−3a (50 ng/mL) | − | + | − | + |
| GENE | ID | Sequence | |
|---|---|---|---|
| 18S rRNA | NR_145820.1 | Forward | GTAACCCGTTGAACCCCATTC |
| Reverse | CCATCCAATCGGTAGTAGCG | ||
| Runx2 | NM_001015051.3 | Forward | TTCGTCAGGATCCTATCAGTTTC |
| Reverse | TTTAATAGCGTGCTGCCATTC | ||
| Osteopontin | NM_000582.2 | Forward | CTGGATGACCAGAGTGCTGA |
| Reverse | TTGCTCTCATCATTGGCTTTC | ||
| Osteocalcin | NM_199173.6 | Forward | CCTCACACTCCTCGCCCTATT |
| Reverse | CCCTCCTGCTTGGACACAAA | ||
| Type-I collagen | NM_000088.4 | Forward | GGGATTCCCTGGACCTAAAG |
| Reverse | TCCCTGAGCTCCAGCCTCTCC | ||
| Periostin | NM_001135934.2 | Forward | AAGCTCAGGATCCTATCAGTTTC |
| Reverse | TGGTTGGCACAAATAATGTCC | ||
| Mkx | NM_001242702.2 | Forward | TTACAAGCACCGTGACAACC |
| Reverse | AAGCCGACGTCTTGCATTAG | ||
| Scx | NM_001080514.3 | Forward | GAGAACACCCAGCCCAAAC |
| Reverse | CTGCGAATCGCTGTCTTTCT | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iizumi, R.; Honda, M. Wnt/β-Catenin Signaling Inhibits Osteogenic Differentiation in Human Periodontal Ligament Fibroblasts. Biomimetics 2022, 7, 224. https://doi.org/10.3390/biomimetics7040224
Iizumi R, Honda M. Wnt/β-Catenin Signaling Inhibits Osteogenic Differentiation in Human Periodontal Ligament Fibroblasts. Biomimetics. 2022; 7(4):224. https://doi.org/10.3390/biomimetics7040224
Chicago/Turabian StyleIizumi, Ryoya, and Michiyo Honda. 2022. "Wnt/β-Catenin Signaling Inhibits Osteogenic Differentiation in Human Periodontal Ligament Fibroblasts" Biomimetics 7, no. 4: 224. https://doi.org/10.3390/biomimetics7040224