
Effect of α-Amylase on the Structure of Chia Seed Mucilage
 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Chia Mucilage Preparation
2.3. α-Amylase/Chia Mucilage Mixture Preparation
2.4. Qualitative Test
2.5. Rheological Measurements, Time Sweep Experiments
3. Results
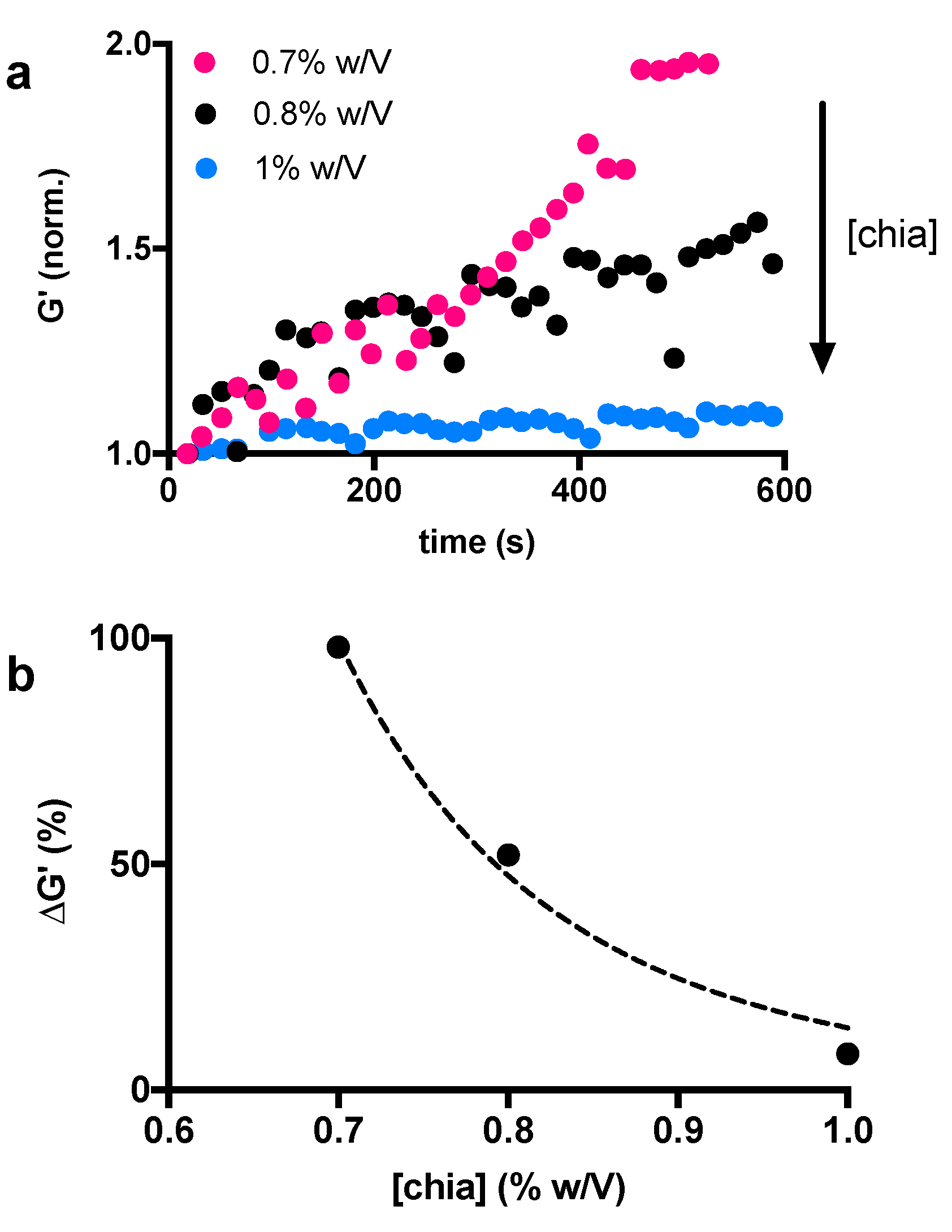
3.1. Role of α-Amylase in Polymer Degradation
3.2. Impact of Amylase Activity on Chia-Based Networks
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cok, M.; Sacco, P.; Porrelli, D.; Travan, A.; Borgogna, M.; Marsich, E.; Paoletti, S.; Donati, I. Mimicking mechanical response of natural tissues. Strain hardening induced by transient reticulation in lactose-modified chitosan (chitlac). Int. J. Biol. Macromol. 2018, 106, 656–660. [Google Scholar] [CrossRef] [PubMed]
- Sacco, P.; Sechi, A.; Trevisan, A.; Picotti, F.; Gianni, R.; Stucchi, L.; Fabbian, M.; Bosco, M.; Paoletti, S.; Marsich, E. A silver complex of hyaluronan–lipoate (SHLS12): Synthesis, characterization and biological properties. Carbohydr. Polym. 2016, 136, 418–426. [Google Scholar] [CrossRef]
- Klemm, D.; Kramer, F.; Moritz, S.; Lindström, T.; Ankerfors, M.; Gray, D.; Dorris, A. Nanocelluloses: A New Family of Nature-Based Materials. Angew. Chem. Int. Ed. 2011, 50, 5438–5466. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef]
- Lopez, F.; Venditti, F.; Cinelli, G.; Ceglie, A. The novel hexadecyltrimethylammonium bromide (CTAB) based organogel as reactor for ester synthesis by entrapped Candida rugosa lipase. Process Biochem. 2006, 41, 114–119. [Google Scholar] [CrossRef]
- Vecchies, F.; Sacco, P.; Marsich, E.; Cinelli, G.; Lopez, F.; Donati, I. Binary Solutions of Hyaluronan and Lactose-Modified Chitosan: The Influence of Experimental Variables in Assembling Complex Coacervates. Polymers 2020, 12, 897. [Google Scholar] [CrossRef]
- Hoffman, A.S. Hydrogels for biomedical applications. Adv. Drug Deliv. Rev. 2012, 64, 18–23. [Google Scholar] [CrossRef]
- Sacco, P.; Decleva, E.; Tentor, F.; Menegazzi, R.; Borgogna, M.; Paoletti, S.; Kristiansen, K.A.; Vårum, K.M.; Marsich, E. Butyrate-Loaded Chitosan/Hyaluronan Nanoparticles: A Suitable Tool for Sustained Inhibition of ROS Release by Activated Neutrophils. Macromol. Biosci. 2017, 17, 1700214. [Google Scholar] [CrossRef]
- Saha, D.; Bhattacharya, S. Hydrocolloids as thickening and gelling agents in food: A critical review. J. Food Sci. Technol. 2010, 47, 587–597. [Google Scholar] [CrossRef]
- Stanley, N.L.; Taylor, L.J. Rheological basis of oral characteristics of fluid and semi-solid foods: A review. Acta Psychol. 1993, 84, 79–92. [Google Scholar] [CrossRef]
- Cuomo, F.; Iacovino, S.; Sacco, P.; de Leonardis, A.; Ceglie, A.; Lopez, F. Progress in Colloid Delivery Systems for Protection and Delivery of Phenolic Bioactive Compounds: Two Study Cases—Hydroxytyrosol and Curcumin. Molecules 2022, 27, 921. [Google Scholar] [CrossRef] [PubMed]
- Prakash, S. From Rheology to Tribology: Applications of Tribology in Studying Food Oral Processing and Texture Perception. In Advances in Food Rheology and Its Applications; Elsevier: Amsterdam, The Netherlands, 2017; pp. 65–86. [Google Scholar] [CrossRef]
- Sacco, P.; Lipari, S.; Cok, M.; Colella, M.; Marsich, E.; Lopez, F.; Donati, I. Insights into Mechanical Behavior and Biological Properties of Chia Seed Mucilage Hydrogels. Gels 2021, 7, 47. [Google Scholar] [CrossRef]
- Li, J.M.; Nie, S.P. The functional and nutritional aspects of hydrocolloids in foods. Food Hydrocoll 2016, 53, 46–61. [Google Scholar] [CrossRef]
- Soukoulis, C.; Cambier, S.; Serchi, T.; Tsevdou, M.; Gaiani, C.; Ferrer, P.; Taoukis, P.S.; Hoffmann, L. Rheological and structural characterisation of whey protein acid gels co-structured with chia (Salvia hispanica L.) or flax seed (Linum usitatissimum L.) mucilage. Food Hydrocoll. 2018, 89, 542–553. [Google Scholar] [CrossRef]
- Muñoz, L. Mucilage from chia seeds (Salvia hispanica): Microestructure, physico-chemical characterization and applications in food industry. Pontificia Universidad Católica de Chile. 2012. Available online: https://repositorio.uc.cl/bitstream/handle/11534/1889/593967.pdf (accessed on 18 September 2022).
- Brütsch, L.; Stringer, F.J.; Kuster, S.; Windhab, E.J.; Fischer, P. Chia seed mucilage—A vegan thickener: Isolation, tailoring viscoelasticity and rehydration. Food Funct. 2019, 10, 4854–4860. [Google Scholar] [CrossRef] [PubMed]
- Timilsena, Y.P.; Adhikari, R.; Kasapis, S.; Adhikari, B. Rheological and microstructural properties of the chia seed polysaccharide. Int. J. Biol. Macromol. 2015, 81, 991–999. [Google Scholar] [CrossRef]
- Cuomo, F.; Iacovino, S.; Cinelli, G.; Messia, M.C.; Marconi, E.; Lopez, F. Effect of additives on chia mucilage suspensions: A rheological approach. Food Hydrocoll 2020, 109, 106118. [Google Scholar] [CrossRef]
- Ferraro, G.; Fratini, E.; Sacco, P.; Asaro, F.; Cuomo, F.; Donati, I.; Lopez, F. Structural characterization and physical ageing of mucilage from chia for food processing applications. Food Hydrocoll. 2020, 129, 107614. [Google Scholar] [CrossRef]
- Coorey, R.; Tjoe, A.; Jayasena, V. Gelling Properties of Chia Seed and Flour. J. Food Sci. 2014, 79, E859–E866. [Google Scholar] [CrossRef] [PubMed]
- Cuomo, F.; Iacovino, S.; Messia, M.C.; Sacco, P.; Lopez, F. Protective action of lemongrass essential oil on mucilage from chia (Salvia hispanica) seeds. Food Hydrocoll. 2020, 105, 105860. [Google Scholar] [CrossRef]
- Timilsena, Y.P.; Adhikari, R.; Kasapis, S.; Adhikari, B. Physicochemical, Thermal and Rheological Characteristics of a Novel Mucilage from Chia Seed (Salvia Hispanica). In Gums and Stabilisers for the Food Industry 18–Hydrocolloid Functionality for Affordable and Sustainable Global Food Solutions; Royal Society of Chemistry: London, UK, 2016; pp. 65–75. [Google Scholar] [CrossRef]
- Fernandes, S.S.; Salas-Mellado, M.D. Addition of chia seed mucilage for reduction of fat content in bread and cakes. Food Chem. 2017, 227, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Gachons, C.P.D.; Breslin, P.A.S. Salivary Amylase: Digestion and Metabolic Syndrome. Curr. Diabetes Rep. 2016, 16, 1–7. [Google Scholar] [CrossRef]
- Dhital, S.; Warren, F.J.; Butterworth, P.J.; Ellis, P.R.; Gidley, M.J. Mechanisms of starch digestion by α-amylase—Structural basis for kinetic properties. Crit. Rev. Food Sci. Nutr. 2017, 57, 875–892. [Google Scholar] [CrossRef] [PubMed]
- Date, K.; Yamazaki, T.; Toyoda, Y.; Hoshi, K.; Ogawa, H. α-Amylase expressed in human small intestinal epithelial cells is essential for cell proliferation and differentiation. J. Cell Biochem. 2020, 121, 1238–1249. [Google Scholar] [CrossRef]
- Mandel, A.L.; Gachons, C.P.D.; Plank, K.L.; Alarcon, S.; Breslin, P.A.S. Individual differences in AMY1 gene copy number, salivary α-amylase levels, and the perception of oral starch. PLoS ONE 2010, 5, e13352. [Google Scholar] [CrossRef]
- Yu, T.-S.; Zeeman, S.C.; Thorneycroft, D.; Fulton, D.C.; Dunstan, H.; Lue, W.-L.; Hegemann, B.; Tung, S.-Y.; Umemoto, T.; Chapple, A.; et al. α-Amylase Is Not Required for Breakdown of Transitory Starch in Arabidopsis Leaves. J. Biol. Chem. 2005, 280, 9773–9779. [Google Scholar] [CrossRef] [PubMed]
- Karimi, M.; Biria, D. The promiscuous activity of alpha-amylase in biodegradation of low-density polyethylene in a polymer-starch blend. Sci. Rep. 2019, 9, 2612. [Google Scholar] [CrossRef]
- Butterworth, P.J.; Warren, F.J.; Ellis, P.R. Human α-amylase and starch digestion: An interesting marriage. Starch-Stärke 2011, 63, 395–405. [Google Scholar] [CrossRef]
- Zinoviadou, K.G.; Janssen, A.M.; de Jongh, H.H.J. Tribological properties of neutral polysaccharide solutions under simulated oral conditions. J. Food Sci. 2008, 73, E88–E94. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Ahmad, M.; Manzoor, K.; Purwar, R.; Ikram, S. A review on latest innovations in natural gums based hydrogels: Preparations & applications. Int. J. Biol. Macromol. 2019, 136, 870–890. [Google Scholar] [CrossRef]
- Bijttebier, A.; Goesaert, H.; Delcour, J.A. Amylase action pattern on starch polymers. Biologia 2008, 63, 989–999. [Google Scholar] [CrossRef]
- Stanley, D.; Farnden, K.J.F.; Macrae, E.A. Plant α-amylases: Functions and roles in carbohydrate metabolism. Biologia 2005, 60, 65–71. [Google Scholar]
- Muralikrishna, G.; Nirmala, M. Cereal α-amylases—An overview. Carbohydr. Polym. 2005, 60, 163–173. [Google Scholar] [CrossRef]
- Furlani, F.; Marfoglia, A.; Marsich, E.; Donati, I.; Sacco, P. Strain Hardening in Highly Acetylated Chitosan Gels. Biomacromolecules 2021, 22, 2902–2909. [Google Scholar] [CrossRef]
- Bianco, S.; Panja, S.; Adams, D.J. Using Rheology to Understand Transient and Dynamic Gels. Gels 2022, 8, 132. [Google Scholar] [CrossRef] [PubMed]
- Grossmann, L.; Wefers, D.; Bunzel, M.; Weiss, J.; Zeeb, B. Accessibility of transglutaminase to induce protein crosslinking in gelled food matrices—Influence of network structure. LWT 2017, 75, 271–278. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Best-Fit Values | [α-Amylase] (mg/mL) | |||
|---|---|---|---|---|
| 2.6 | 5 | 15 | 20 | |
| a | 1.91 ± 0.09 | 1.73 ± 0.06 | 1.87 ± 0.10 | 2.06 ± 0.12 |
| b | −0.168 ± 0.008 | −0.174 ± 0.007 | −0.199 ± 0.010 | −0.235 ± 0.011 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piazza, F.; Colella, M.; Cinelli, G.; Lopez, F.; Donati, I.; Sacco, P. Effect of α-Amylase on the Structure of Chia Seed Mucilage. Biomimetics 2022, 7, 141. https://doi.org/10.3390/biomimetics7040141
Piazza F, Colella M, Cinelli G, Lopez F, Donati I, Sacco P. Effect of α-Amylase on the Structure of Chia Seed Mucilage. Biomimetics. 2022; 7(4):141. https://doi.org/10.3390/biomimetics7040141
Chicago/Turabian StylePiazza, Francesco, Matilde Colella, Giuseppe Cinelli, Francesco Lopez, Ivan Donati, and Pasquale Sacco. 2022. "Effect of α-Amylase on the Structure of Chia Seed Mucilage" Biomimetics 7, no. 4: 141. https://doi.org/10.3390/biomimetics7040141