
Bacillus subtilis Provides Long-Term Protection in a Murine Model of Allergic Lung Disease by Influencing Bacterial Composition

 and
and 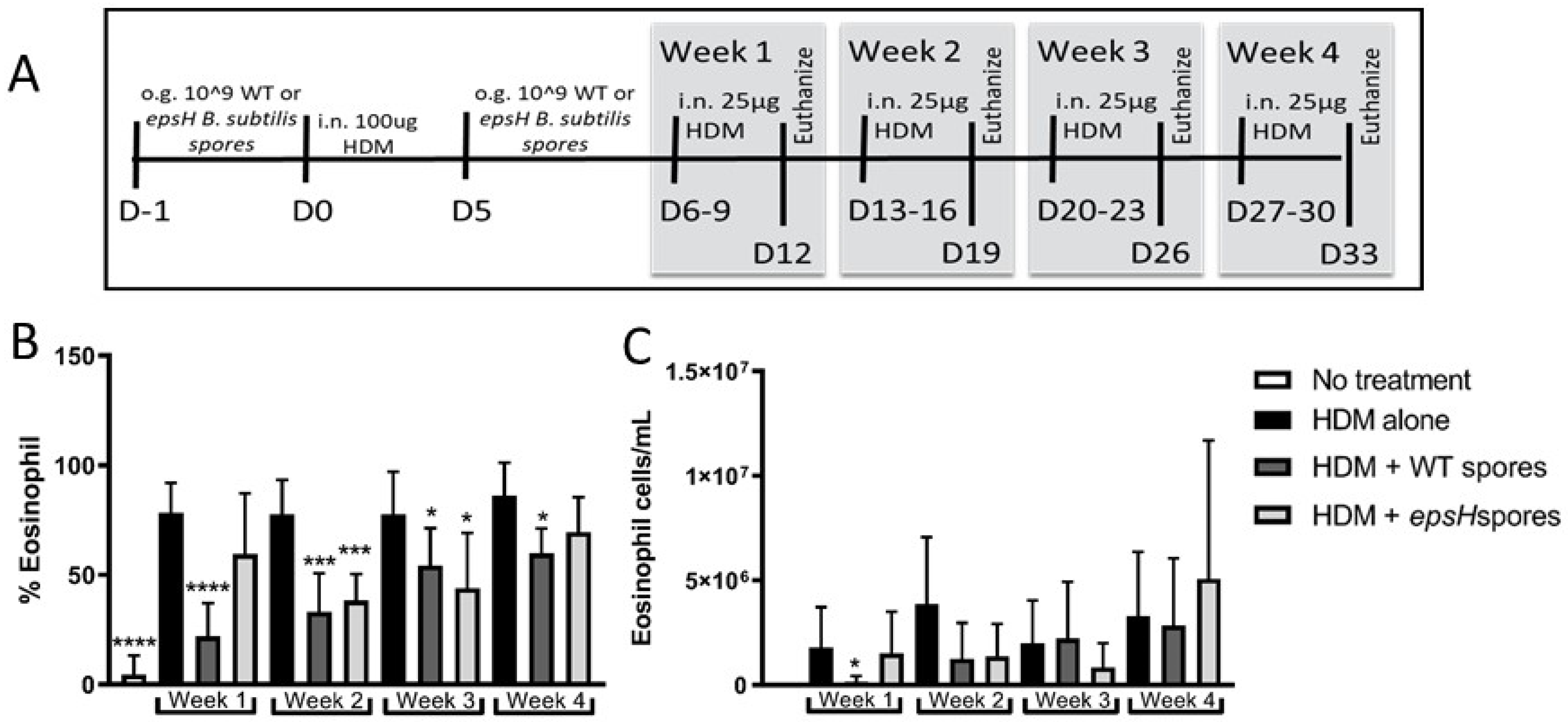{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Allergic Lung Disease Model
2.3. ELISA
2.4. DNA Extraction and 16S rRNA Amplicon Sequencing
2.5. Statistics
3. Results
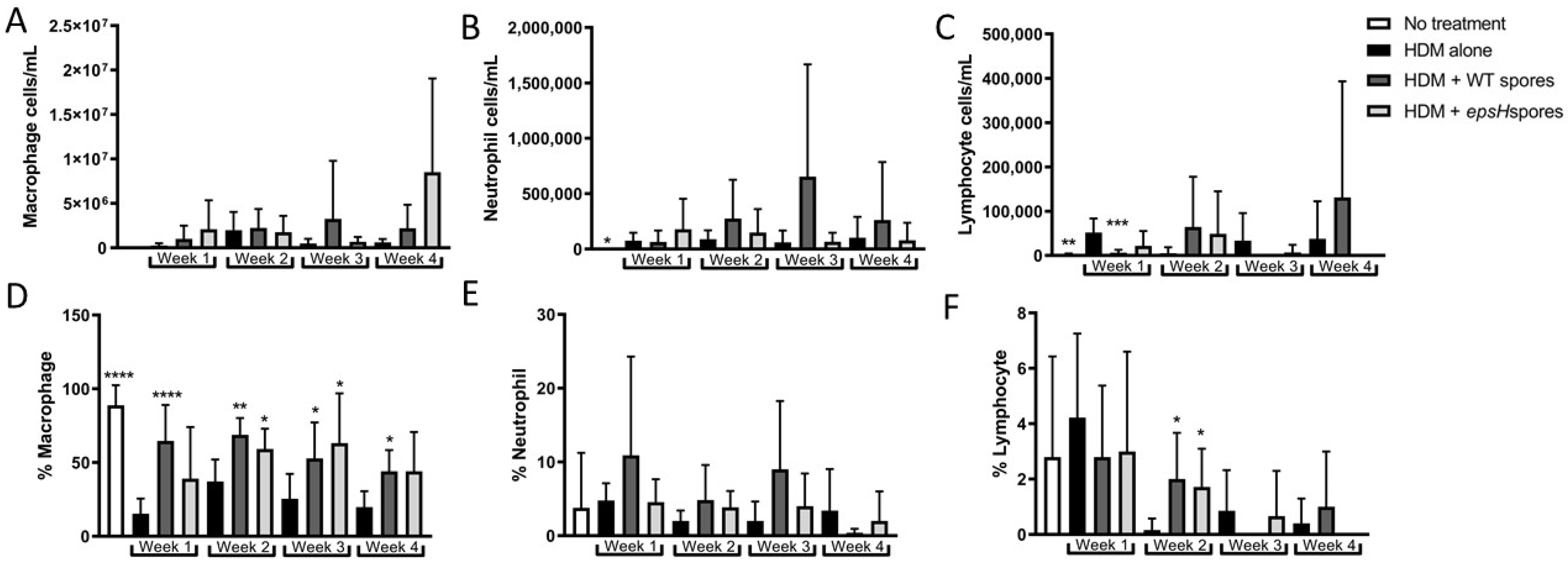
3.1. Bacillus Subtilis Protection from House-Dust Mite-Induced Eosinophilia Wanes after 3 Weeks
3.2. CCL24 Production Is Suppressed by Bacillus subtilis Treatment for 3 Weeks
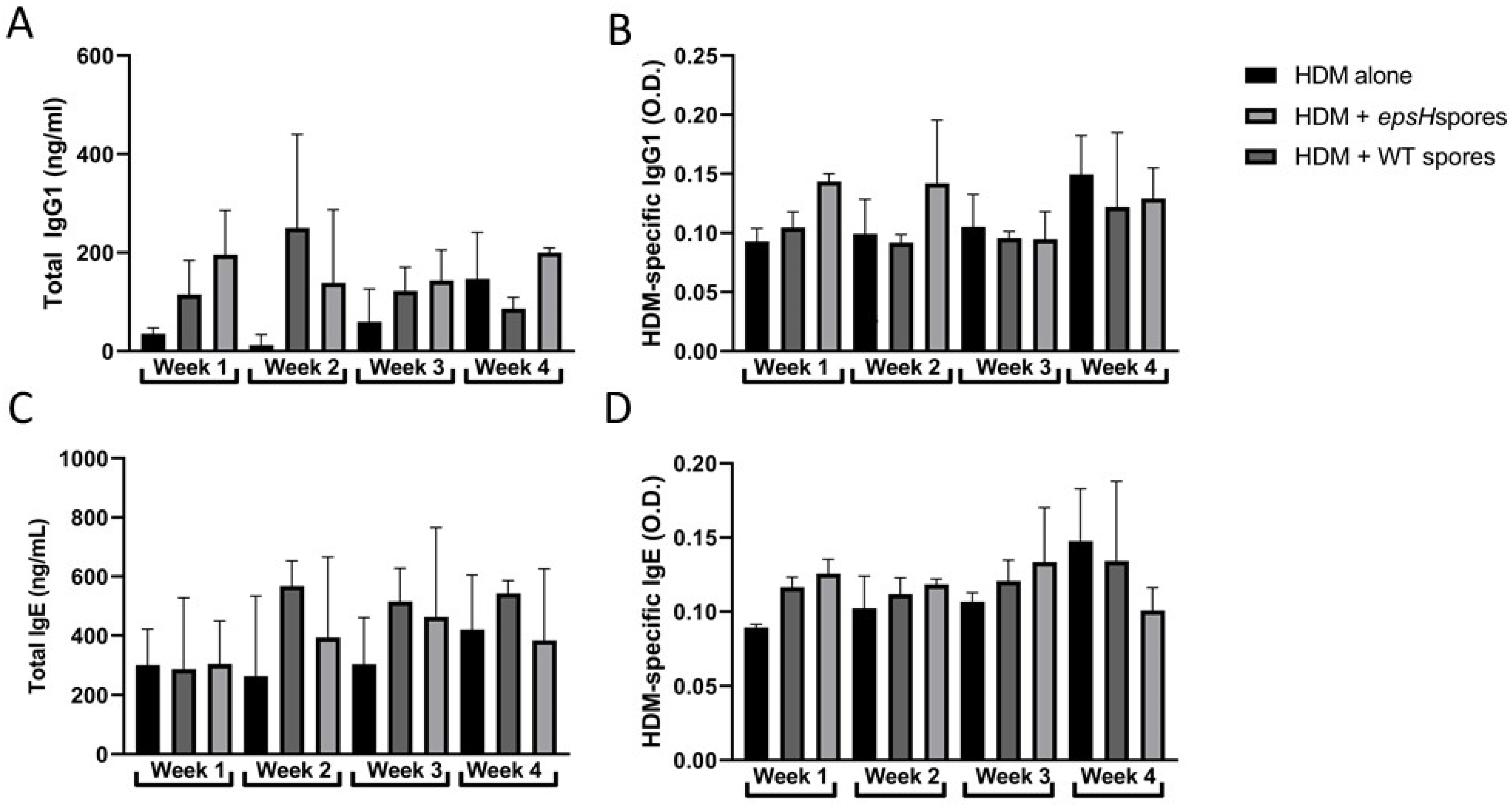
3.3. B. subtilis Treatment Does Not Influence Immunoglobulin Production
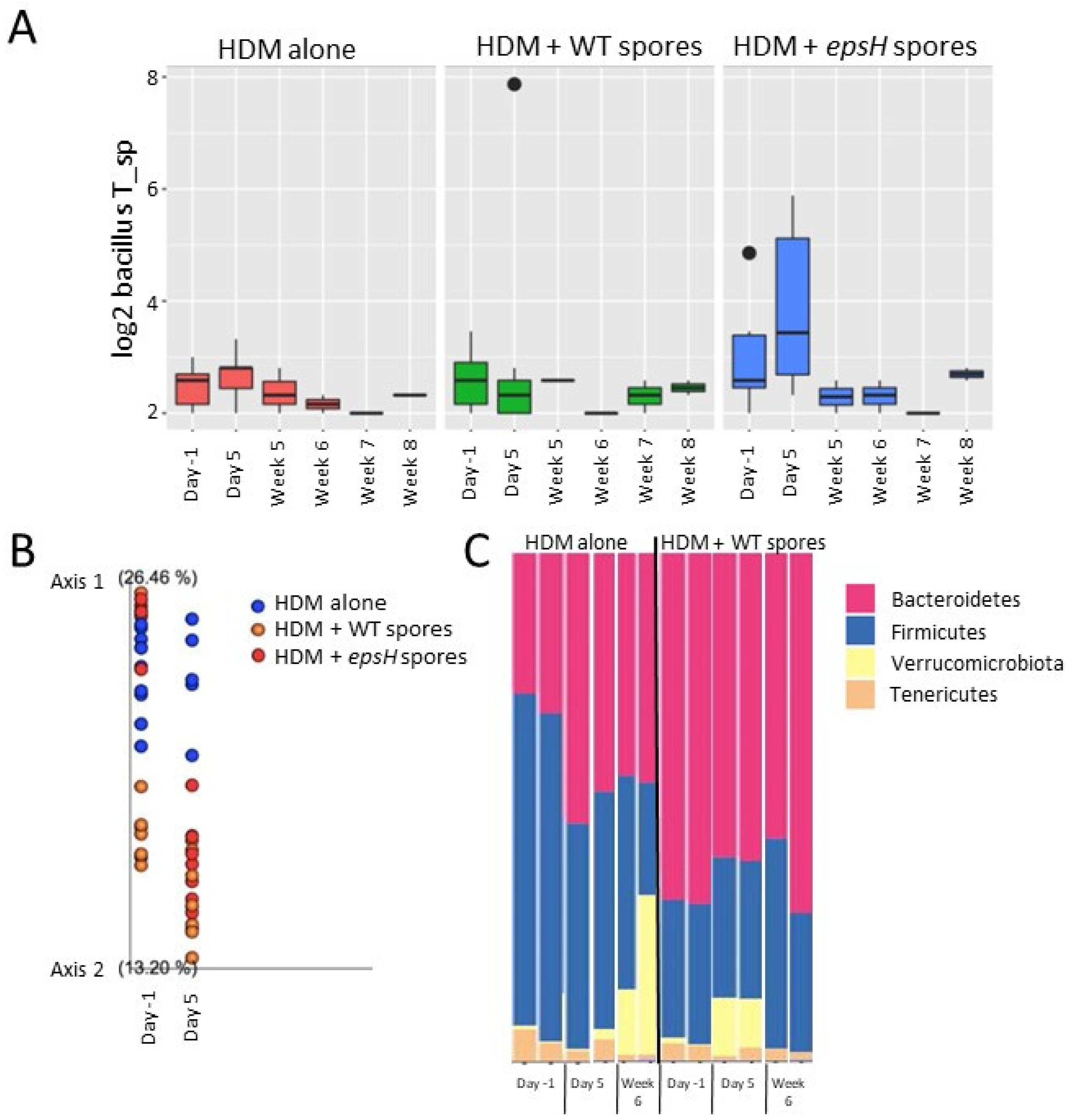
3.4. Bacillus Fecal Composition Is Similar across Mice Regardless of Treatment with B. subtilis Spores
3.5. Bacterial Composition Clustering Correlates with Protection from Eosinophilia
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Platts-Mills, T. The allergy epidemics: 1870–2010. J. Allergy Clin. Immunol. 2015, 136, 3–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akdis, M. Healthy immune response to allergens: T regulatory cells and more. Curr. Opin. Immunol. 2006, 18, 738–744. [Google Scholar] [CrossRef] [PubMed]
- Galazzo, G.; van Best, N.; Bervoets, L.; Dapaah, I.O.; Savelkoul, P.H.; Hornef, M.W.; Consortium, G.-M.; Lau, S.; Hamelmann, E.; Penders, J. Development of the Microbiota and Associations With Birth Mode, Diet, and Atopic Disorders in a Longitudinal Analysis of Stool Samples, Collected From Infancy Through Early Childhood. Gastroenterology 2020, 158, 1584–1596. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.A.; Sjogren, Y.M.; Persson, J.O.; Nilsson, C.; Sverremark-Ekstrom, E. Early colonization with a group of Lactobacilli decreases the risk for allergy at five years of age despite allergic heredity. PLoS ONE 2011, 6, e23031. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.O.; Kim, H.J.; Kim, Y.J.; Kang, M.J.; Kwon, J.W.; Seo, J.H.; Kim, H.Y.; Kim, B.J.; Yu, J.; Hong, S.J. Asthma Prevention by Lactobacillus Rhamnosus in a Mouse Model is Associated With CD4+CD25+Foxp3+ T Cells. Allergy Asthma Immunol. Res. 2012, 4, 150–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raftis, E.J.; Delday, M.I.; Cowie, P.; McCluskey, S.M.; Singh, M.D.; Ettorre, A.; Mulder, I.E. Bifidobacterium breve MRx0004 protects against airway inflammation in a severe asthma model by suppressing both neutrophil and eosinophil lung infiltration. Sci. Rep. 2018, 8, 12024. [Google Scholar] [CrossRef]
- Aoki-Yoshida, A.; Yamada, K.; Hachimura, S.; Sashihara, T.; Ikegami, S.; Shimizu, M.; Totsuka, M. Enhancement of Oral Tolerance Induction in DO11.10 Mice by Lactobacillus gasseri OLL2809 via Increase of Effector Regulatory T Cells. PLoS ONE 2016, 11, e0158643. [Google Scholar] [CrossRef] [Green Version]
- Smelt, M.J.; de Haan, B.J.; Bron, P.A.; van Swam, I.; Meijerink, M.; Wells, J.M.; Faas, M.M.; de Vos, P. L. Plantarum, L. salivarius, and L. lactis attenuate Th2 responses and increase Treg frequencies in healthy mice in a strain dependent manner. PLoS ONE 2012, 7, e47244. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.; Kamiya, T.; Liu, Y.; Kadoki, M.; Kakuta, S.; Oshima, K.; Hattori, M.; Takeshita, K.; Kanai, T.; Saijo, S.; et al. Inhibition of Dectin-1 Signaling Ameliorates Colitis by Inducing Lactobacillus-Mediated Regulatory T Cell Expansion in the Intestine. Cell Host Microbe 2015, 18, 183–197. [Google Scholar] [CrossRef] [Green Version]
- Sherid, M.; Samo, S.; Sulaiman, S.; Husein, H.; Sifuentes, H.; Sridhar, S. Liver abscess and bacteremia caused by lactobacillus: Role of probiotics? Case report and review of the literature. BMC Gastroenterol. 2016, 16, 138. [Google Scholar] [CrossRef]
- Swartzendruber, J.A.; Incrocci, R.W.; Wolf, S.A.; Jung, A.; Knight, K.L. Bacillus subtilis exopolysaccharide prevents allergic eosinophilia. Allergy 2019, 74, 819–821. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Yi, H. An exopolysaccharide from Bacillus subtilis alleviates airway inflammatory responses via the NF-kappaB and STAT6 pathways in asthmatic mice. Biosci. Rep. 2022, 42, BSR20212461. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.E.; Paynich, M.L.; Kearns, D.B.; Knight, K.L. Protection from Intestinal Inflammation by Bacterial Exopolysaccharides. J. Immunol. 2014, 192, 4813–4820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paik, W.; Alonzo, F., 3rd; Knight, K.L. Probiotic Exopolysaccharide Protects against Systemic Staphylococcus aureus Infection, Inducing Dual-Functioning Macrophages That Restrict Bacterial Growth and Limit Inflammation. Infect. Immun. 2019, 87, e00791-18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piewngam, P.; Zheng, Y.; Nguyen, T.H.; Dickey, S.W.; Joo, H.S.; Villaruz, A.E.; Glose, K.A.; Fisher, E.L.; Hunt, R.L.; Li, B.; et al. Pathogen elimination by probiotic Bacillus via signalling interference. Nature 2018, 562, 532–537. [Google Scholar] [CrossRef]
- Paynich, M.L.; Jones-Burrage, S.E.; Knight, K.L. Exopolysaccharide from Bacillus subtilis Induces Anti-Inflammatory M2 Macrophages That Prevent T Cell-Mediated Disease. J. Immunol. 2017, 198, 2689–2698. [Google Scholar] [CrossRef] [Green Version]
- Hammad, H.; Plantinga, M.; Deswarte, K.; Pouliot, P.; Willart, M.A.; Kool, M.; Muskens, F.; Lambrecht, B.N. Inflammatory dendritic cells—Not basophils—Are necessary and sufficient for induction of Th2 immunity to inhaled house dust mite allergen. J. Exp. Med. 2010, 207, 2097–2111. [Google Scholar] [CrossRef] [Green Version]
- Havenith, C.E.; Breedijk, A.J.; Betjes, M.G.; Calame, W.; Beelen, R.H.; Hoefsmit, E.C. T cell priming in situ by intratracheally instilled antigen-pulsed dendritic cells. Am. J. Respir. Cell Mol. Biol. 1993, 8, 319–324. [Google Scholar] [CrossRef]
- Shimoda, K.; van Deursen, J.; Sangster, M.Y.; Sarawar, S.R.; Carson, R.T.; Tripp, R.A.; Chu, C.; Quelle, F.W.; Nosaka, T.; Vignali, D.A.; et al. Lack of IL-4-induced Th2 response and IgE class switching in mice with disrupted Stat6 gene. Nature 1996, 380, 630–633. [Google Scholar] [CrossRef]
- Heiman, A.S.; Abonyo, B.O.; Darling-Reed, S.F.; Alexander, M.S. Cytokine-stimulated human lung alveolar epithelial cells release eotaxin-2 (CCL24) and eotaxin-3 (CCL26). J. Interferon Cytokine Res. 2005, 25, 82–91. [Google Scholar] [CrossRef]
- Pope, S.M.; Zimmermann, N.; Stringer, K.F.; Karow, M.L.; Rothenberg, M.E. The eotaxin chemokines and CCR3 are fundamental regulators of allergen-induced pulmonary eosinophilia. J. Immunol. 2005, 175, 5341–5350. [Google Scholar] [CrossRef] [Green Version]
- D’Arienzo, R.; Maurano, F.; Mazzarella, G.; Luongo, D.; Stefanile, R.; Ricca, E.; Rossi, M. Bacillus subtilis spores reduce susceptibility to Citrobacter rodentium-mediated enteropathy in a mouse model. Res. Microbiol. 2006, 157, 891–897. [Google Scholar] [CrossRef] [PubMed]
- Chung, K.F. Precision medicine in asthma: Linking phenotypes to targeted treatments. Curr. Opin. Pulm. Med. 2018, 24, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Virkud, Y.V.; Burks, A.W.; Steele, P.H.; Edwards, L.J.; Berglund, J.P.; Jones, S.M.; Scurlock, A.M.; Perry, T.T.; Pesek, R.D.; Vickery, B.P. Novel baseline predictors of adverse events during oral immunotherapy in children with peanut allergy. J. Allergy Clin. Immunol. 2017, 139, 882–888.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conroy, D.M.; Williams, T.J. Eotaxin and the attraction of eosinophils to the asthmatic lung. Respir. Res. 2001, 2, 150–156. [Google Scholar] [CrossRef] [Green Version]
- Humbles, A.A.; Conroy, D.M.; Marleau, S.; Rankin, S.M.; Palframan, R.T.; Proudfoot, A.E.; Wells, T.N.; Li, D.; Jeffery, P.K.; Griffiths-Johnson, D.A.; et al. Kinetics of eotaxin generation and its relationship to eosinophil accumulation in allergic airways disease: Analysis in a guinea pig model in vivo. J. Exp. Med. 1997, 186, 601–612. [Google Scholar] [CrossRef] [Green Version]
- Ying, S.; Robinson, D.S.; Meng, Q.; Rottman, J.; Kennedy, R.; Ringler, D.J.; Mackay, C.R.; Daugherty, B.L.; Springer, M.S.; Durham, S.R.; et al. Enhanced expression of eotaxin and CCR3 mRNA and protein in atopic asthma. Association with airway hyperresponsiveness and predominant co-localization of eotaxin mRNA to bronchial epithelial and endothelial cells. Eur. J. Immunol. 1997, 27, 3507–3516. [Google Scholar] [CrossRef]
- Feehley, T.; Plunkett, C.H.; Bao, R.; Choi Hong, S.M.; Culleen, E.; Belda-Ferre, P.; Campbell, E.; Aitoro, R.; Nocerino, R.; Paparo, L.; et al. Healthy infants harbor intestinal bacteria that protect against food allergy. Nat. Med. 2019, 25, 448–453. [Google Scholar] [CrossRef]
- Jones, S.E.; Knight, K.L. Bacillus subtilis-Mediated Protection from Citrobacter rodentium-Associated Enteric Disease Requires espH and Functional Flagella. Infect. Immun. 2012, 80, 710–719. [Google Scholar] [CrossRef]
- Pawankar, R.; Canonica, G.W.; Holgate, S.T.; Lockey, R.F.; Blaiss, M. World Allergy Organization (WAO) white book on allergy. Nihon Shoni Arerugi Gakkaishi Jpn. J. Pediatr. Allergy Clin. Immunol. 2011, 25, 341. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monroy Del Toro, R.; Incrocci, R.; Negris, O.; McGrath, S.; Swartzendruber, J.A. Bacillus subtilis Provides Long-Term Protection in a Murine Model of Allergic Lung Disease by Influencing Bacterial Composition. Allergies 2023, 3, 1-10. https://doi.org/10.3390/allergies3010001
Monroy Del Toro R, Incrocci R, Negris O, McGrath S, Swartzendruber JA. Bacillus subtilis Provides Long-Term Protection in a Murine Model of Allergic Lung Disease by Influencing Bacterial Composition. Allergies. 2023; 3(1):1-10. https://doi.org/10.3390/allergies3010001
Chicago/Turabian StyleMonroy Del Toro, Rosalinda, Ryan Incrocci, Olivia Negris, Shaina McGrath, and Julie A. Swartzendruber. 2023. "Bacillus subtilis Provides Long-Term Protection in a Murine Model of Allergic Lung Disease by Influencing Bacterial Composition" Allergies 3, no. 1: 1-10. https://doi.org/10.3390/allergies3010001