
In Vitro Hypoxic Environment Enhances Volatile Compound Production in Persian Violet Flowers


Abstract
:
1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Volatile Organic Compound Analysis
- = retention index;
- = retention time of sample peak;
- = retention time of n-alkane peak eluting immediately before sample peak;
- = retention time of n-alkane peak eluting immediately after sample peak;
- = carbon number of n-alkane peak eluting immediately before sample peak.

2.3. Odor-Active Compounds Determination
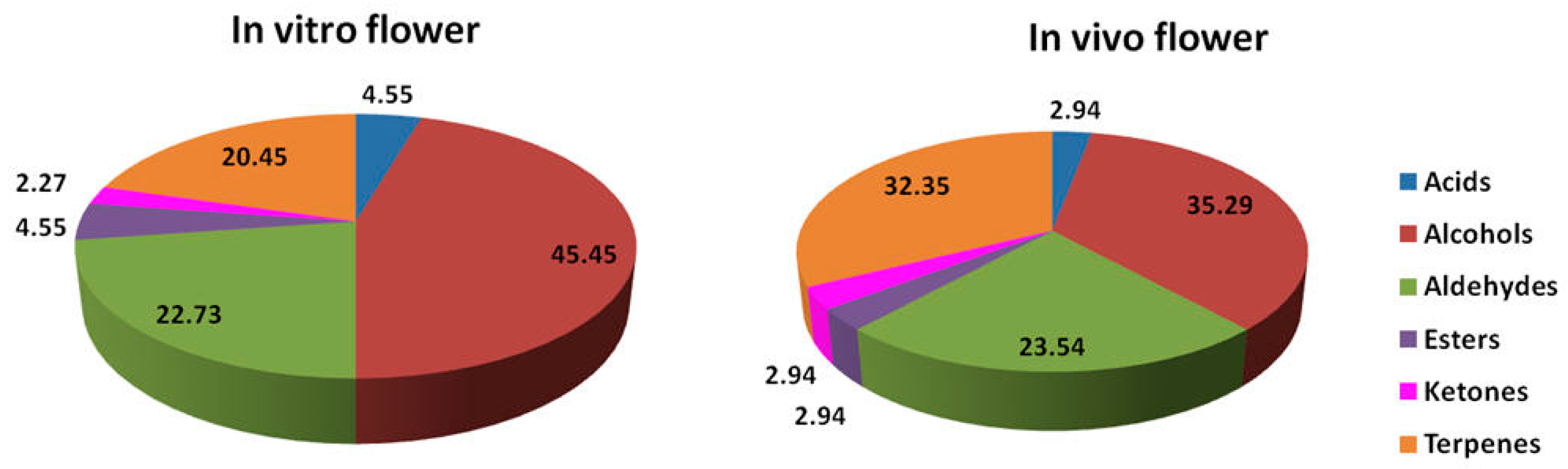
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Riseman, A.; Sumanasinghe, V.A.; Craig, R. Cytology, crossability, and pollen fertility of Sri Lankan Exacum (Gentianaceae) and their hybrids. Int. J. Plant Sci. 2006, 167, 191–199. [Google Scholar] [CrossRef]
- Skrzypczak-Pietraszek, E. Phytochemistry and biotechnology approaches of the genus Exacum. In The Gentianaceae; Rybczyński, J., Davey, M., Mikuła, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2015; Volume 2, pp. 383–401. [Google Scholar]
- Srichuay, W.; Promchan, T.; Te-chato, S. Effect of chlorine dioxide (ClO2) on culture medium sterilization on micropropagation of Persian violet (Exacum affine Balf.f. ex Regel). Int. J. Agric. Technol. 2018, 14, 259–270. [Google Scholar]
- Phanomchai, S.; Noichinda, S.; Kachonpadungkitti, Y.; Bodhipadma, K. Differing in vitro rooting and flowering responses of the Persian violet to low and high UV-C irradiation. Plants 2021, 10, 2671. [Google Scholar] [CrossRef] [PubMed]
- Gilman, E.F.; Howe, T. Exacum affine Persian Violet, German Violet; IFAS Extension; University of Florida: Gainesville, FL, USA, 2023; pp. 1–2. [Google Scholar]
- Riseman, A. Exacum: Exacum affine and related species. In Flower Breeding and Genetics: Issues, Challenges, and Opportunities for the 21st Century; Anderson, N.O., Ed.; Springer: Dordrecht, The Netherlands, 2007; pp. 601–622. [Google Scholar]
- Iwasaki, N.; Sata, Y.; Hisajima, S. Life cycle of Arabidopsis (Arabidopsis thaliana) plant in vitro. J. Shita 2005, 17, 34–38. [Google Scholar] [CrossRef]
- Haque, S.M.; Halder, T.; Ghosh, B. In vitro completion of sexual life cycle—Production of next sporophytic generation through in vitro flowering and fruiting in Solanum americanum and Solanum villosum. S. Afr. J. Bot. 2018, 118, 112–119. [Google Scholar] [CrossRef]
- Sukthavornthum, W.; Bodhipadma, K.; Noichinda, S.; Phanomchai, S.; Deelueak, U.; Kachonpadungkitti, Y.; Leung, D.W.M. UV-C irradiation induced alterations in shoot proliferation and in vitro flowering in plantlets developed from encapsulated and non-encapsulated microshoots of Persian violet. Sci. Hortic. 2018, 233, 9–13. [Google Scholar] [CrossRef]
- Sreelekshmi, R.; Siril, E.A. Investigation on in vitro bouquets and flower longevity of micropropagated Dianthus chinensis L. Sci. Hortic. 2021, 275, 109708. [Google Scholar] [CrossRef]
- Bodhipadma, K.; Noichinda, S.; Tangtivaporn, C.; Phanomchai, S.; Leung, D.W.M. Micropropagation and in vitro inflorescence of Pentas lanceolata. Appl. Sci. Eng. Prog. 2023, 16, 5692. [Google Scholar] [CrossRef]
- Kumari, P.; Panwar, S.; Namita; Soni, A. Biosynthesis, composition and sources of floral scent in ornamental crops: A review. Chem. Sci. Rev. Lett. 2017, 6, 1502–1509. [Google Scholar]
- Fan, J.; Zhang, W.; Zhang, D.; Wang, G.; Cao, F. Flowering stage and daytime affect scent emission of Malus ioensis“Prairie Rose”. Molecules 2019, 24, 2356. [Google Scholar] [CrossRef]
- Picazo-Aragonés, J.; Terrab, A.; Balao, F. Plant volatile organic compounds evolution: Transcriptional regulation, epigenetics and polyploidy. Int. J. Mol. Sci. 2020, 21, 8956. [Google Scholar] [CrossRef]
- Giovannini, A.; Laura, M.; Nesi, B.; Savona, M.; Cardi, T. Genes and genome editing tools for breeding desirable phenotypes in ornamentals. Plant Cell Rep. 2021, 40, 461–478. [Google Scholar] [CrossRef] [PubMed]
- Pichersky, E.; Dudareva, N. Scent engineering: Toward the goal of controlling how flowers smell. Trends Biotechnol. 2007, 25, 105–110. [Google Scholar] [CrossRef]
- Wright, G.A.; Schiestl, F.P. The evolution of floral scent: The influence of olfactory learning by insect pollinators on the honest signalling of floral rewards. Funct. Ecol. 2009, 23, 841–851. [Google Scholar] [CrossRef]
- Mostafa, S.; Wang, Y.; Zeng, W.; Jin, B. Floral scents and fruit aromas: Functions, compositions, biosynthesis, and regulation. Front. Plant Sci. 2022, 13, 860157. [Google Scholar] [CrossRef] [PubMed]
- Gardner, E.M.; Gagné, R.J.; Kendra, P.E.; Montgomery, W.S.; Raguso, R.A.; McNeil, T.T.; Zerega, N.J.C. A flower in fruit’s clothing: Pollination of jackfruit (Artocarpus heterophyllus, Moraceae) by a new species of gall midge, Clinodiplosis ultracrepidata sp. nov. (Diptera: Cecidomyiidae). Int. J. Plant Sci. 2018, 179, 350–367. [Google Scholar] [CrossRef]
- Dudareva, N.; Murfitt, L.M.; Mann, C.J.; Gorenstein, N.; Kolosova, N.; Kish, C.M.; Bonham, C.; Wood, K. Developmental regulation of methyl benzoate biosynthesis and emission in snapdragon flowers. Plant Cell. 2000, 12, 949–961. [Google Scholar] [CrossRef]
- Engelmann, W. Rhythms in Organisms—Observing, Experimenting, Recording and Analyzing; Institute of Botany and Physiological Ecology of Plants, University of Tübingen: Tübingen, Germany, 2022; pp. 21–124. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio-assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Retention Index Mixture for GC (R8769)—Bulletin. Available online: https://www.sigmaaldrich.com/deepweb/assets/sigmaaldrich/product/documents/260/555/r8769bul.pdf (accessed on 21 June 2023).
- Aschariyaphotha, W.; Wongs-Aree, C.; Bodhipadma, K.; Noichinda, S. Fruit volatile fingerprints characterized among four commercial cultivars of Thai durian (Duriozibethinus). J. Food Qual. 2021, 2021, 1383927. [Google Scholar] [CrossRef]
- van Gemert, L.J. Odour Thresholds: Compilations of Odour Threshold Values in Air, Water and Other Media; Oliemans Punter & Partners BV: Utrecht, The Netherlands, 2011; p. 486. [Google Scholar]
- Hasnain, A.; Naqvi, S.A.H.; Ayesha, S.I.; Khalid, F.; Ellahi, M.; Iqbal, S.; Hassan, M.Z.; Abbas, A.; Adamski, R.; Markowska, D.; et al. Plants in vitro propagation with its applications in food, pharmaceuticals and cosmetic industries; current scenario and future approaches. Front. Plant Sci. 2022, 13, 1009395. [Google Scholar] [CrossRef] [PubMed]
- Kaviani, B.; Deltalab, B.; Kulus, D.; Tymoszuk, A.; Bagheri, H.; Azarinejad, T. In vitro propagation of Pyracantha angustifolia (Franch.) C.K. Schneid. Horticulturae 2022, 8, 964. [Google Scholar] [CrossRef]
- Ørnstrup, H.; MØlgaard, J.P.; Farestveit, B. Somatic embryogenesis and plant regeneration from cell suspensions of Exacum affine. Plant Cell Tissue Organ Cult. 1993, 35, 37–41. [Google Scholar] [CrossRef]
- Sarai, N.; Bodhipadma, K.; Noichinda, S.; Luangsriumporn, P.; Leung, D.W.M. Microshoot culture of Persian violet: Plant regeneration and in vitro flowering. Ann. Agric. Sci. 2017, 62, 105–111. [Google Scholar] [CrossRef]
- Kadohisa, M. Effects of odor on emotion, with implications. Front. Syst. Neurosci. 2013, 7, 66. [Google Scholar] [CrossRef] [PubMed]
- Najar, B.; Marchioni, I.; Ruffoni, B.; Copetta, A.; Pistelli, L.; Pistelli, L. Volatilomic analysis of four edible flowers from Agastache genus. Molecules 2019, 24, 4480. [Google Scholar] [CrossRef]
- Wendin, K.; Pálsdóttir, A.M.; Spendrup, S.; Mårtensson, L. Odor perception and descriptions of rose-scented geranium Pelargonium graveolens ‘Dr. Westerlund’–sensory and chemical analyses. Molecules 2023, 28, 4511. [Google Scholar] [CrossRef]
- Peng, H.-P.; Chan, C.-S.; Shih, M.-C.; Yang, S.F. Signaling events in the hypoxic induction of alcohol dehydrogenase gene in Arabidopsis. Plant Physiol. 2001, 126, 742–749. [Google Scholar] [CrossRef]
- Midzi, J.; Jeffery, D.W.; Baumann, U.; Rogiers, S.; Tyerman, S.D.; Pagay, V. Stress-induced volatile emissions and signalling in inter-plant communication. Plants 2022, 11, 2566. [Google Scholar] [CrossRef]
- Nagegowda, D.A. Plant volatile terpenoid metabolism: Biosynthetic genes, transcriptional regulation and subcellular compartmentation. FEBS Lett. 2010, 584, 2965–2973. [Google Scholar] [CrossRef]
- Couillaud, J.; Leydet, L.; Duquesne, K.; Iacazio, G. The terpene mini-path, a new promising alternative for terpenoids bio-production. Genes 2021, 12, 1974. [Google Scholar] [CrossRef]
- Borges, C.V.; Orsi, R.O.; Maraschin, M.; Lima, G.P.P. Oxidative stress in plants and the biochemical response mechanism. In Plant Stress Mitigators: Types, Techniques and Functions; Ghorbanpour, M., Shahid, M.A., Eds.; Academic Press: Cambridge, MA, USA, 2023; pp. 455–468. [Google Scholar]
- Wongs-Aree, C.; Noichinda, S. Glycolysis fermentative by-products and secondary metabolites involved in plant adaptation under hypoxia during pre- and postharvest. In Hypoxia and Anoxia; Das, K., Biradar, M.S., Eds.; IntechOpen: London, UK, 2018; pp. 59–72. [Google Scholar]
- Wolf, S.J.; Craig, R. Inheritance of flower and stem color in Exacum affine Balf. J. Hered. 1988, 79, 303–306. [Google Scholar]
- Buchbauer, G.; Jirovetz, L.; Wasicky, M. Volatile constituents of Exacum affine Balf. f. flowers obtained by dynamic headspace sampling and as the essential oil. Flav. Fragr. J. 1994, 9, 55–58. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Volatile Component | Relative Content (mg Thiophene/gFW) | OAV | Odor Characteristics | ||
|---|---|---|---|---|---|
| In Vitro | In Vivo | In Vitro | In Vivo | ||
| Acids | |||||
| Acetic acid | 0.0039 | 0.0016 | 0.75 | 0.1231 | Vinegar |
| tran-2-Pentenoic acid | 0.0005 | nd | na | na | Sour, caramel |
| Aldehydes | |||||
| Acetaldehyde | 0.1318 | 0.0729 | 48.81 | 27.0 | Pungent |
| 3-Methylbutanal | 0.013 | 0.0074 | 20.312 | 4.628 | Peach-like |
| 2-Butenal | 0.0139 | 0.0057 | 0.516 | 0.0521 | Sweet |
| 2-Pentenal | 0.0004 | nd | 0.0008 | - | Tomato-like |
| Hexanal | 0.0011 | 0.0038 | 0.012 | 0.165 | Green apple |
| 2-Hexenal | 0.0028 | 0.004 | 0.012 | 0.165 | Vegetable-like |
| Heptanal | 0.0008 | nd | 0.0081 | - | Fatty, oily |
| Octanal | nd | 0.001 | - | 0.0059 | Honey-like |
| Nonanal | nd | 0.0044 | - | 0.419 | Fatty |
| 2-Methylbenzaldehyde | 0.0005 | 0.0013 | 0.115 | 0.01354 | Cherry-like |
| Benzaldehyde | 0.0017 | nd | 0.049 | - | Fruity |
| Benzeneacetaldehyde | 0.0004 | nd | 158.73 | - | Hyacinth-like |
| Ketone | |||||
| Acetone | 0.0206 | 0.0145 | 0.026 | 0.0077 | Sweet |
| Alcohols | |||||
| Ethanol | 0.0865 | 0.0708 | 0.349 | 0.1142 | Wine |
| 3-Penten-2-one | 0.0077 | nd | 15.917 | - | Fruity |
| 3-Methyl-2-butanol | 0.0189 | nd | 23.049 | - | Fruity |
| 4-Methyl-2-pentanol | 0.0006 | nd | 0.0011 | - | Pungent |
| 1-Penten-3-ol | 0.0023 | 0.004 | 0.014 | 0.0009 | Grassy-green |
| 3-Hexanol | 0.0004 | nd | 0.0012 | - | Alcoholic |
| 3-Methyl-1-butanol | 0.0015 | nd | 0.623 | - | Pungent |
| 4-Methyl-2-pentanol | 0.0014 | nd | 0.003 | - | Pungent |
| 3,5-Dimethylphenol | 0.0011 | nd | 67.5 | - | Fruity |
| 1-Pentanol | nd | 0.0014 | - | 0.0092 | Sweet |
| 1-Hexanol | 0.0067 | 0.031 | 0.494 | 0.9118 | Green-fruity |
| 2-Heptanol | 0.0008 | nd | 0.02 | - | Lemon |
| 2-Penten-1-ol | 0.0013 | 0.0017 | 3.0 | 1.544 | Green note |
| 2-Hexen-1-ol | 0.0026 | nd | 18.37 | - | Fruity-green |
| 3-Hexen-1-ol | 0.0365 | 0.1096 | 7.015 | 8.4308 | Grassy-green |
| 4-Hexen-1-ol | 0.0005 | 0.0014 | 12 | 14 | Pungent-oily |
| 3-Methyl-2-heptanol | nd | 0.0005 | - | 0.005 | Citrus |
| 1-Octen-3-ol | nd | 0.0046 | - | 0.46 | Herbaceous |
| 6-Methyl-5-hepten-2-ol | 0.0005 | nd | 0.6 | - | Fruity |
| 2-Ethyl-1-hexanol | 0.0008 | nd | 0.0025 | - | Floral |
| 1-Octanol | 0.0003 | 0.0021 | 0.036 | 0.0955 | Orange-rose |
| Benzyl alcohol | 0.0032 | 0.0017 | 0.0031 | 0.0007 | Fruity |
| Phenylethyl alcohol | 0.0017 | 0.0005 | 0.2048 | 0.0238 | Rose |
| Esters | |||||
| Geranyl acetate | 0.0006 | nd | 0.015 | - | Floral |
| Methyl salicylate | 0.001 | 0.0015 | 0.00002 | 0.00003 | Minty, wintergreen |
| Terpenes | |||||
| γ-Terpinene | 0.0013 | nd | 0.000049 | - | Lemon |
| 2-Carene | 0.0003 | 0.0008 | 0.2 | 0.2 | Floral |
| 3-Carene | nd | 0.0018 | - | 0.0005 | Citrus |
| Caryophyllene | nd | 0.0045 | - | 0.0571 | Floral |
| Humulene | nd | 0.0008 | - | 5.0 | Woody |
| β-Ionone | nd | 0.0004 | - | 0.025 | Floral |
| Eucalyptol | 0.0033 | 0.0052 | 0.0041 | 0.0026 | Camphoraceous |
| Terpinen-4-ol | 0.0015 | 0.0006 | 3.17 | 0.5 | Spicy |
| α-Terpineol | 0.0007 | 0.006 | 0.0198 | 0.0007 | Floral |
| β-Citronellol | 0.0392 | 0.0117 | 0.035 | 0.0042 | Rose |
| Linalool | 0.0013 | 0.0006 | 0.0005 | 0.0001 | Floral |
| β-Farnesene | 0.0003 | nd | 9.195 | - | Woody |
| α-Farnesene | 0.0005 | 0.0016 | 14.94 | 18.3908 | Woody |
| Terpene | In Vitro | In Vivo | Terpene Type |
|---|---|---|---|
| γ-Terpinene | ++ | - | Monoterpene |
| 2-Carene | + | + | Monoterpene |
| 3-Carene | - | ++ | Monoterpene |
| Caryophyllene | - | ++ | Sesquiterpene |
| Humulene | - | + | Sesquiterpene |
| β-Ionone | - | + | Monoterpene |
| Eucalyptol | + | + | Monoterpene |
| Terpinen-4-ol | ++ | + | Terpene alcohol |
| α-Terpineol | + | ++ | Terpene alcohol |
| β-Citronellol | +++ | ++ | Terpene alcohol |
| Linalool | ++ | + | Terpene alcohol |
| β-Farnesene | + | - | Sesquiterpene |
| α-Farnesene | + | ++ | Sesquiterpene |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noichinda, S.; Bodhipadma, K. In Vitro Hypoxic Environment Enhances Volatile Compound Production in Persian Violet Flowers. Horticulturae 2023, 9, 981. https://doi.org/10.3390/horticulturae9090981
Noichinda S, Bodhipadma K. In Vitro Hypoxic Environment Enhances Volatile Compound Production in Persian Violet Flowers. Horticulturae. 2023; 9(9):981. https://doi.org/10.3390/horticulturae9090981
Chicago/Turabian StyleNoichinda, Sompoch, and Kitti Bodhipadma. 2023. "In Vitro Hypoxic Environment Enhances Volatile Compound Production in Persian Violet Flowers" Horticulturae 9, no. 9: 981. https://doi.org/10.3390/horticulturae9090981