
Performance Evaluation of New Table Grape Varieties under High Light Intensity Conditions Based on the Photosynthetic and Chlorophyll Fluorescence Characteristics


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Site Overview
2.2. Experimental Plant Materials
2.3. Test Equipment and Test Reagents
2.4. Test Methods
2.4.1. Photo-Response Curve Determination
2.4.2. Measurement of Chlorophyll Fluorescence Parameters
2.4.3. Measurement of Leaf Appearance Traits
2.4.4. Measurement of Chlorophyll Content
2.4.5. TOPSIS Evaluation Method
2.5. Data Processing and Statistical Analysis
3. Results
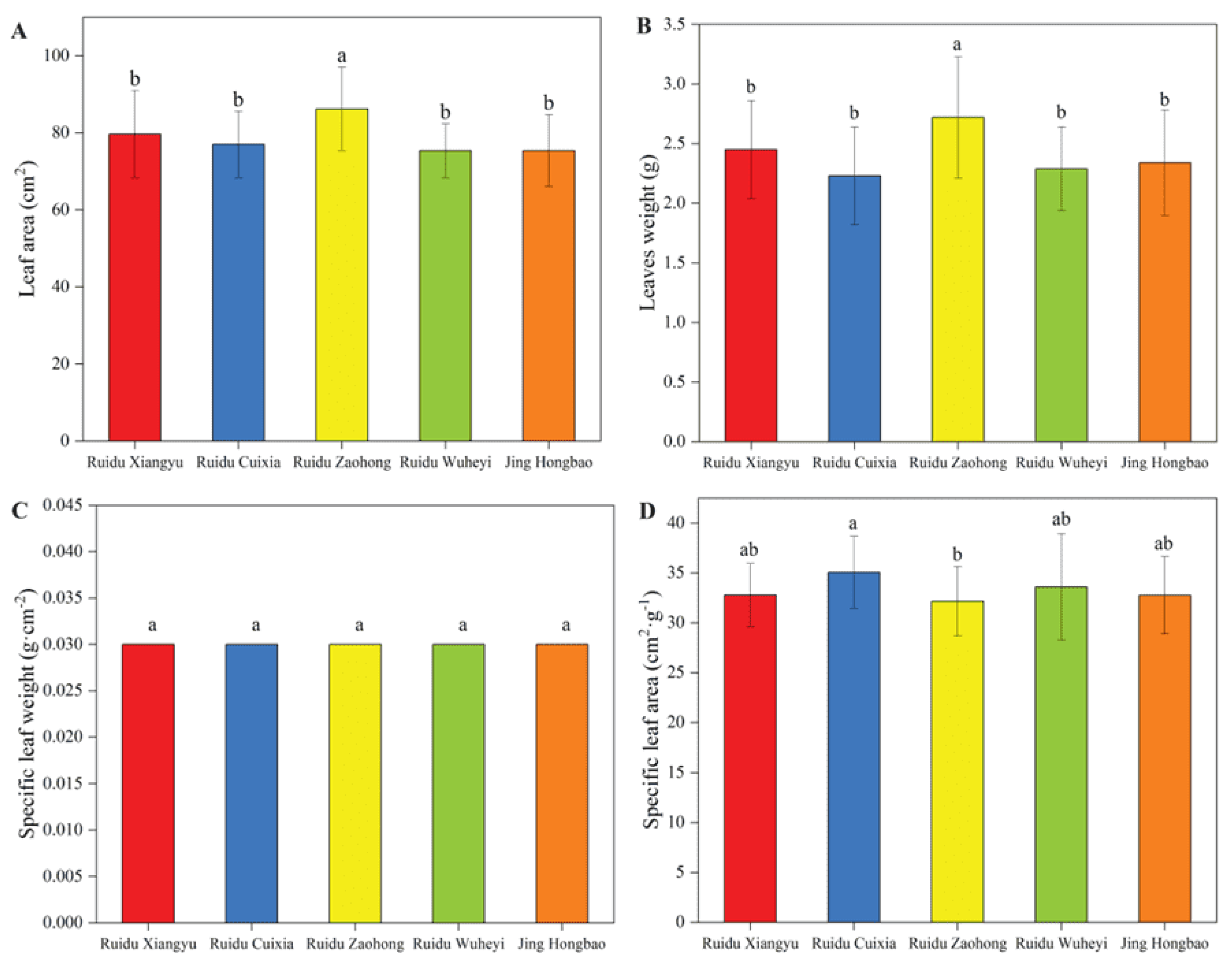
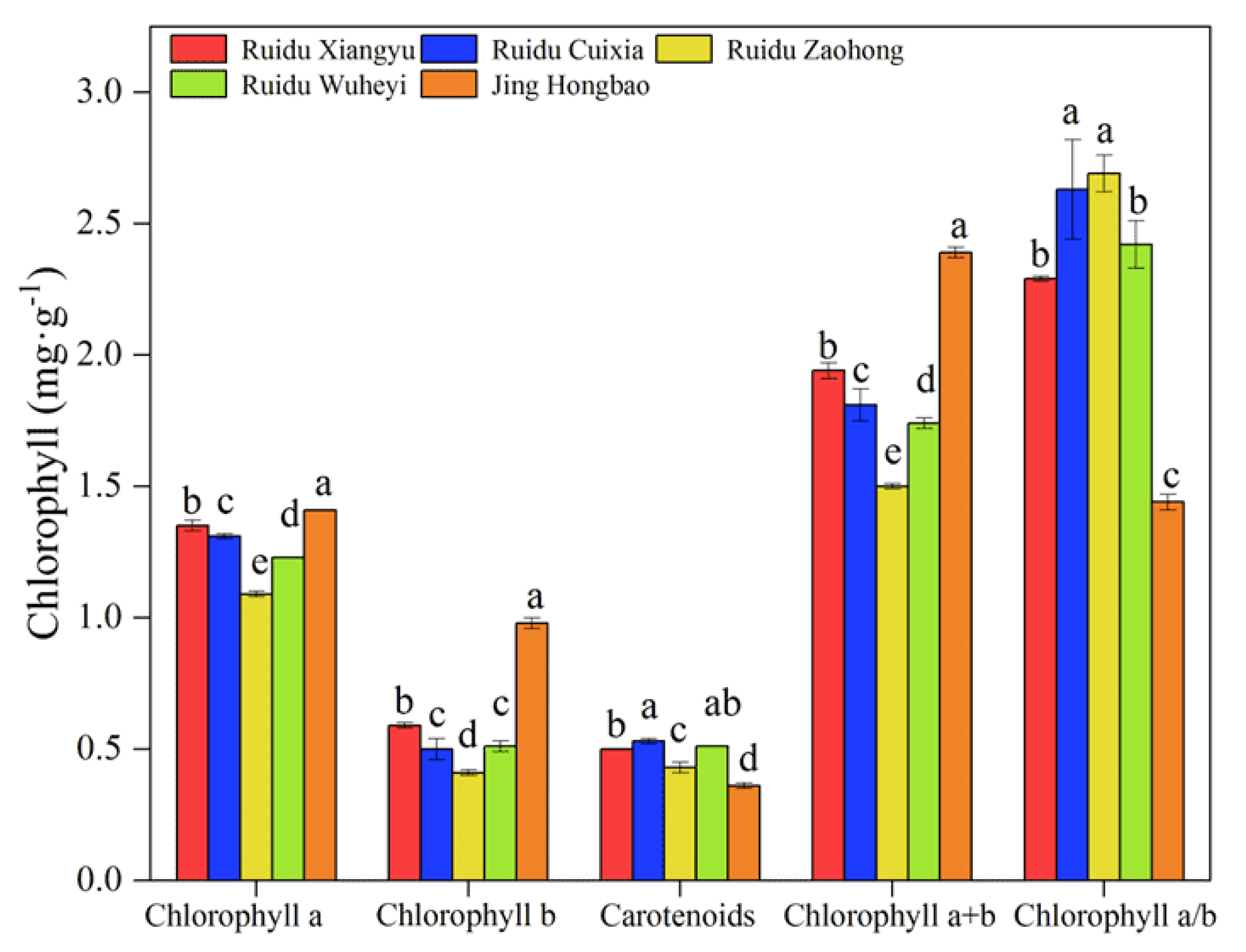
3.1. Chlorophyll Content and Leaf Appearance Traits
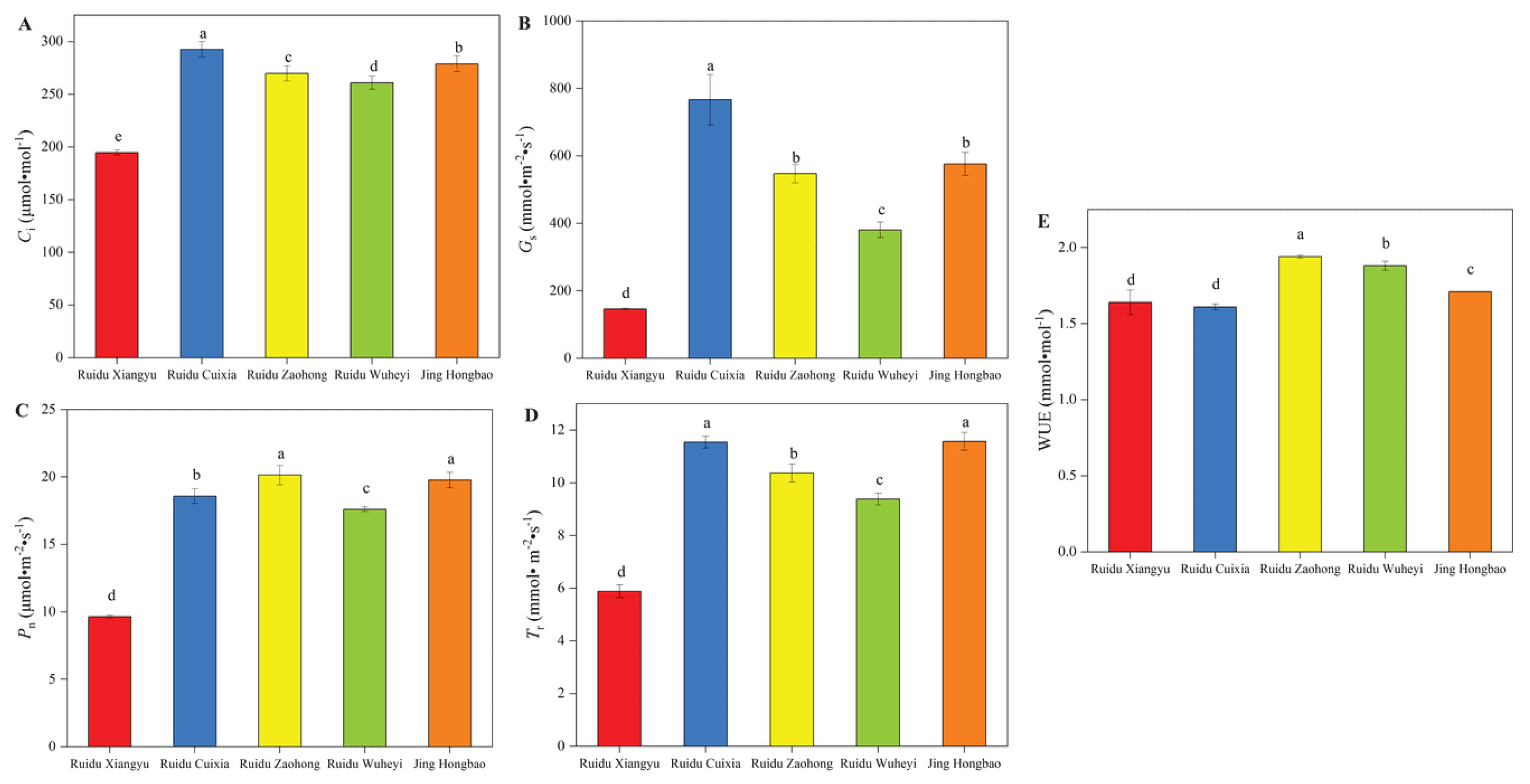
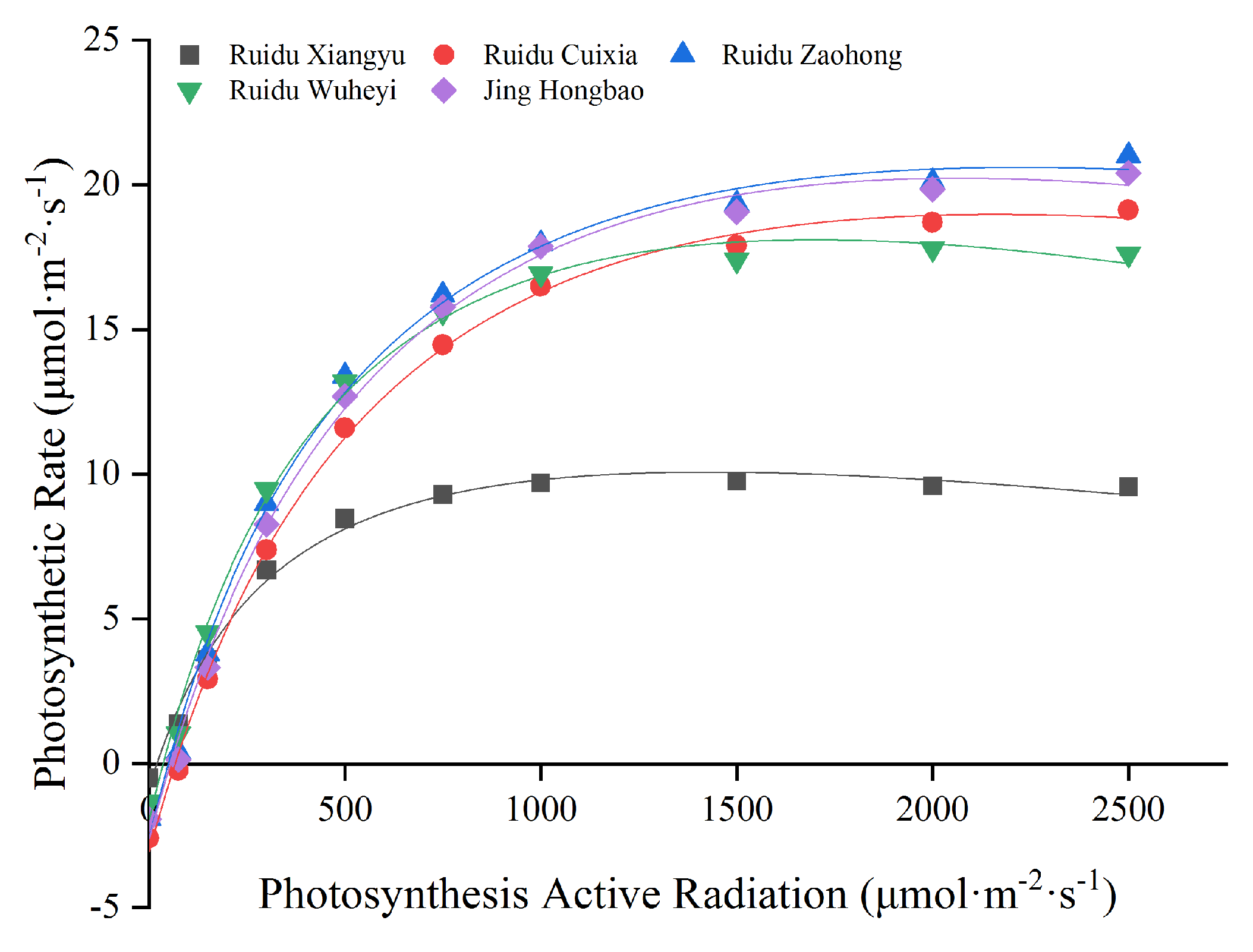
3.2. Photosynthetic Parameters and Photo-Response Curve
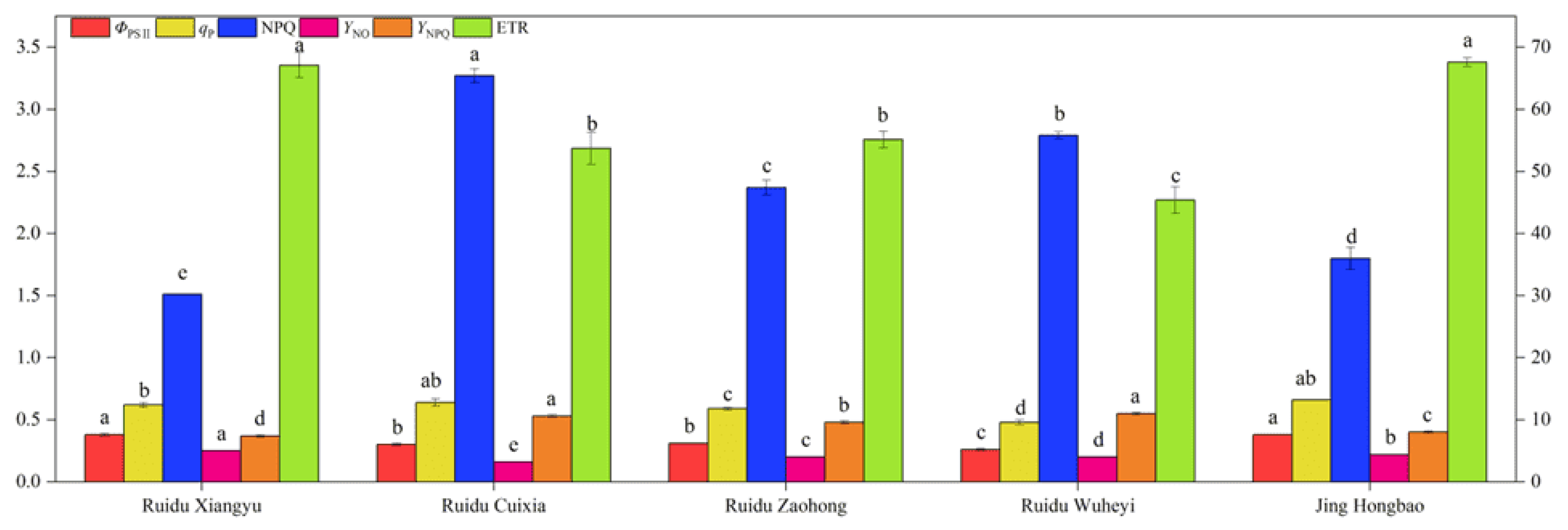
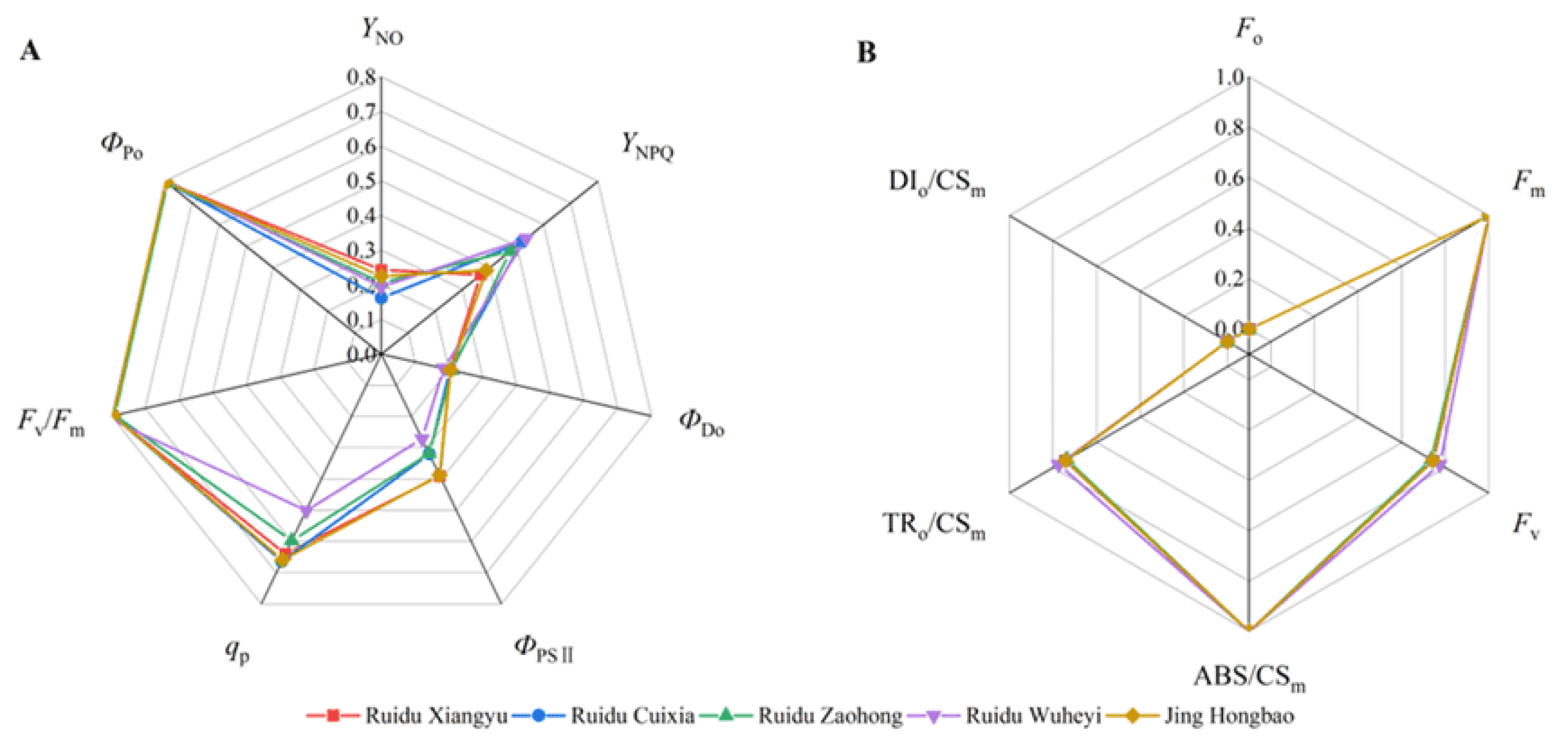
3.3. Chlorophyll Fluorescence Parameters
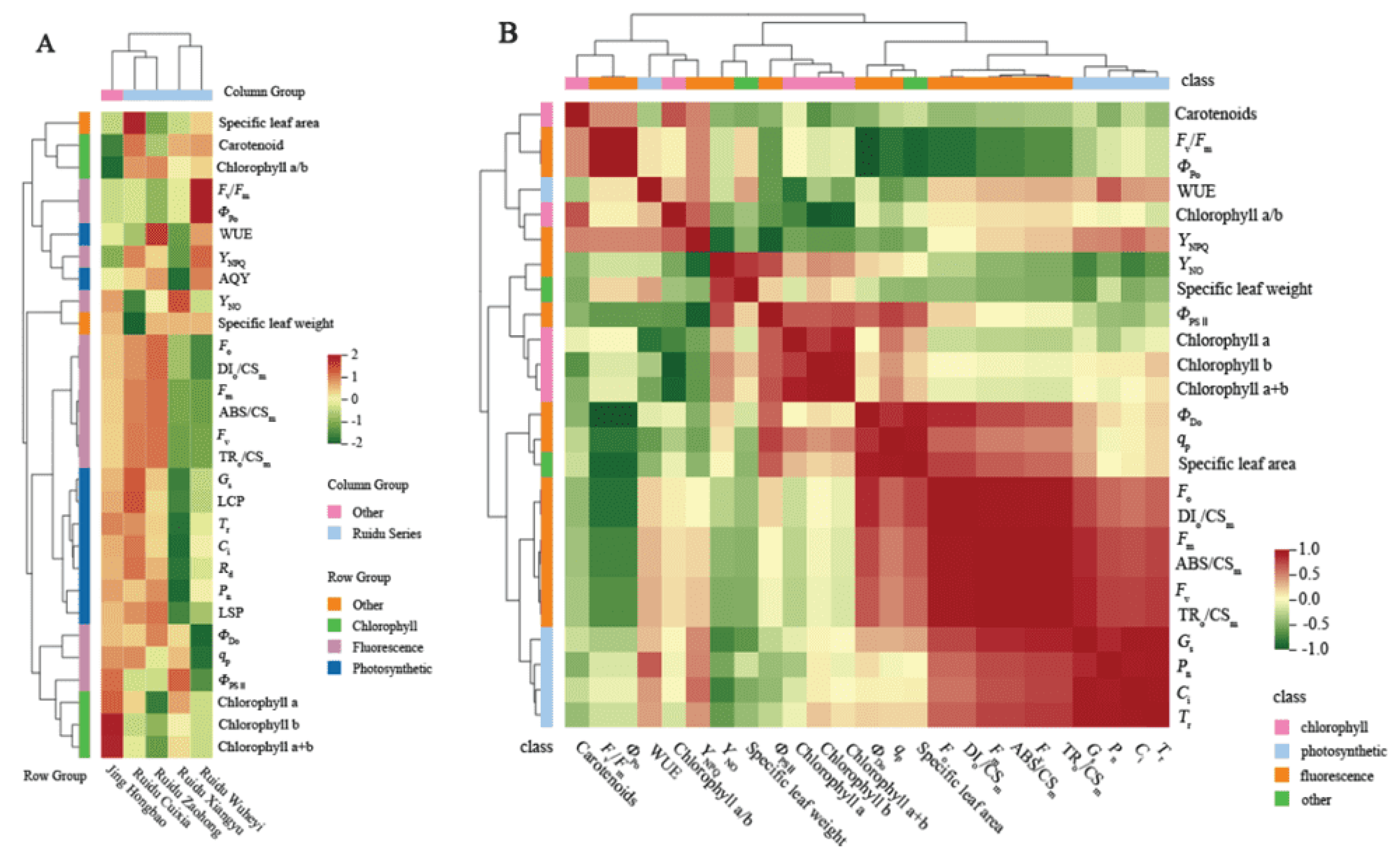
3.4. Correlation Analysis and Hierarchical Cluster Analysis
3.5. Principal Component Analysis
3.6. Comprehensive Evaluation of Photosynthetic Capacity
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- He, B.B.; Sheng, Y.; Cao, W.; Wu, J.C. Characteristics of Climate Change in Northern Xinjiang in 1961–2017, China. Chin. Geogr. Sci. 2019, 30, 249–265. [Google Scholar] [CrossRef]
- Ivashchenko, I.B.A.; Rakhmetov, D.; Fishchenko, V. Biochemical peculiarities of Glebionis coronaria (Asteraceae) introduced in Central Polissya of Ukraine. Plant Fungal Res. 2019, 2, 32–39. [Google Scholar] [CrossRef]
- Ivashchenko, I.; Kotyuk, L.; Bakalova, A. Morphology and productivity of tarragon (Artemisia dracunculus L.) in Central Polissya (Ukraine). Ukr. J. Ecol. 2020, 10, 48–55. [Google Scholar]
- Tsai, Y.C.; Chen, K.C.; Cheng, T.S.; Lee, C.; Lin, S.H.; Tung, C.W. Chlorophyll fluorescence analysis in diverse rice varieties reveals the positive correlation between the seedlings salt tolerance and photosynthetic efficiency. BMC Plant Biol. 2019, 19, 403. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.; Harris, P.J.C. Photosynthesis under stressful environments: An overview. Photosynthetica 2013, 51, 163–190. [Google Scholar] [CrossRef]
- Wei, S.S.; Wang, X.Y.; Shi, D.; Li, Y.H.; Zhang, J.W.; Liu, P.; Zhao, B.; Dong, S. The mechanisms of low nitrogen induced weakened photosynthesis in summer maize (Zea mays L.) under field conditions. Plant Physiol. Biochem. 2016, 105, 118–128. [Google Scholar] [CrossRef] [PubMed]
- Du, X.H.; Peng, F.R.; Jiang, J.; Tan, P.P.; Wu, Z.Z.; Liang, Y.W.; Zhong, Z.K. Inorganic nitrogen fertilizers induce changes in ammonium assimilation and gas exchange in Camellia sinensis L. Turk. J. Agric. For. 2015, 39, 28–38. [Google Scholar] [CrossRef]
- Mathur, S.; Tomar, R.S.; Jajoo, A. Arbuscular Mycorrhizal fungi (AMF) protects photosynthetic apparatus of wheat under drought stress. Photosynth. Res. 2019, 139, 227–238. [Google Scholar] [CrossRef]
- Wang, Y.; Jie, W.; Peng, X.; Hua, X.; Yan, X.; Zhou, Z.; Lin, J. Physiological adaptive strategies of oil seed crop Ricinus communis early seedlings (cotyledon vs. true leaf) under salt and alkali stresses: From the growth, photosynthesis and chlorophyll fluorescence. Front. Plant Sci. 2019, 9, 1939. [Google Scholar] [CrossRef]
- Tantray, A.Y.; Bashir, S.S.; Ahmad, A. Low nitrogen stress regulates chlorophyll fluorescence in coordination with photosynthesis and Rubisco efficiency of rice. Physiol. Mol. Biol. Plants 2020, 26, 83–94. [Google Scholar] [CrossRef]
- Yan, Z.; Ma, T.; Guo, S.; Liu, R.; Li, M. Leaf anatomy, photosynthesis and chlorophyll fluorescence of lettuce as influenced by arbuscular mycorrhizal fungi under high temperature stress. Sci. Hortic. 2021, 280, 109933. [Google Scholar] [CrossRef]
- Kromdijk, J.; Głowacka, K.; Leonelli, L.; Gabilly, S.T.; Iwai, M.; Niyogi, K.K.; Long, S.P. Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science 2016, 354, 857–861. [Google Scholar] [CrossRef]
- Zhao, L.S.; Li, K.; Wang, Q.M.; Song, X.Y.; Su, H.N.; Xie, B.B.; Zhang, X.Y.; Huang, F.; Chen, X.L.; Zhou, B.C. Nitrogen Starvation Impacts the Photosynthetic Performance of Porphyridium cruentum as Revealed by Chlorophyll a Fluorescence. Sci. Rep. 2017, 7, 8542. [Google Scholar] [CrossRef] [PubMed]
- Bark, K.M.; Forcé, R.K. Fluorescence properties of fluoranthene as a function of temperature and environment. Spectrochim. Acta Part A Mol. Spectrosc. 1993, 49, 1605–1611. [Google Scholar] [CrossRef]
- Hazrati, S.; Tahmasebi-Sarvestani, Z.; Modarres-Sanavy, S.A.M.; Mokhtassi-Bidgoli, A.; Nicola, S. Effects of water stress and light intensity on chlorophyll fluorescence parameters and pigments of Aloe vera L. Plant Physiol. Biochem. 2016, 106, 141–148. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Yuan, Y.; Wang, W.; Xie, Q.; Chen, Q. Effects of heat stress on chlorophyll fluorescence characteristics of Rhododendron leaves. Jiangsu Agric. Sci. 2019, 47, 144–148. [Google Scholar]
- Li, Z.; Ji, W.; Hong, E.; Fan, Z.; Lin, B.; Xia, X.; Chen, X.; Zhu, X. Study on Heat Resistance of Peony Using Photosynthetic Indexes and Rapid Fluorescence Kinetics. Horticulturae 2023, 9, 100. [Google Scholar] [CrossRef]
- Van der Westhuizen, M.M.; Oosterhuis, D.M.; Berner, J.M.; Boogaers, N. Chlorophyll a fluorescence as an indicator of heat stress in cotton (Gossypium hirsutum L.). S. Afr. J. Plant Soil 2020, 37, 116–119. [Google Scholar] [CrossRef]
- Carlson, J.E.; Adams, C.A.; Holsinger, K.E. Intraspecific variation in stomatal traits, leaf traits and physiology reflects adaptation along aridity gradients in a South African shrub. Ann. Bot. 2016, 117, 195–207. [Google Scholar] [CrossRef]
- Liu, Q.Q.; Huang, Z.J.; Wang, Z.N.; Chen, Y.F.; Wen, Z.M.; Liu, B.; Tigabu, M. Responses of leaf morphology, NSCs contents and C:N:P stoichiometry of Cunninghamia lanceolata and Schima superba to shading. BMC Plant Biol. 2020, 20, 354. [Google Scholar] [CrossRef]
- Wu, J.A.R.; Zhong, H.; Yadav, V.; Zhang, C.; Ma, Y.; Liu, X.; Zhang, F.; Zha, Q.; Wang, X. The Impact of High Temperatures in the Field on Leaf Tissue Structure in Different Grape Cultivars. Horticulturae 2023, 9, 731. [Google Scholar] [CrossRef]
- Altaf, M.A.; Hao, Y.; Shu, H.; Jin, W.; Chen, C.; Li, L.; Zhang, Y.; Mumtaz, M.A.; Fu, H.; Cheng, S.; et al. Melatonin mitigates cold-induced damage to pepper seedlings by promoting redox homeostasis and regulating antiox-idant profiling. Hortic. Plant J. 2023, 9, 1–13. [Google Scholar]
- Altaf, A.; Nawaz, F.; Majeed, S.; Ahsan, M.; Ahmad, K.S.; Akhtar, G.; Shehzad, M.A.; Rashad Javeed, H.M.; Farman, M. Foliar Humic Acid and Salicylic Acid Application Stimulates Physiological Responses and Antioxidant Systems to Im-prove Maize Yield under Water Limitations. JSFA Rep. 2023, 3, 119–128. [Google Scholar] [CrossRef]
- Ye, Z.P.; Li, J.S. Comparative investigation light response of photosynthesis on non-rectangular hyperbola model and modified model of rectangular hyperbola. J. Jinggangshan Univ. 2010, 31, 38–44. [Google Scholar]
- Strasser, R.J.; Srivastava, A.; Tsimilli-Michael, M. The fluorescence transient as a tool to characterize and screen photosynthetic samples. Probing Photosynth. Mech. Regul. Adapt. 2000, 25, 445–483. [Google Scholar]
- Tsimilli-Michael, M. Revisiting JIP-test: An educative review on concepts, assumptions, approximations, definitions and terminology. Photosynthetica 2019, 57, 90–107. [Google Scholar]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. polyphenoloxidase in beta Vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef]
- Ma, X.H.; Song, L.L.; Yu, W.W.; Hu, Y.Y.; Liu, Y.; Wu, J.S.; Ying, Y.Q. Growth, physiological, and biochemical responses of Camptothecaacuminata seedlings to different light environments. Front. Plant Sci. 2015, 6, 321. [Google Scholar] [CrossRef]
- Chaves, M.M.; Flexas, J.; Pinheiro, C. Photosynthesis under drought and salt stress: Regulation mechanisms from whole plant to cell. Ann. Bot. 2009, 103, 551–560. [Google Scholar] [CrossRef]
- Tang, H.; Hu, Y.Y.; Yu, W.W.; Song, L.L.; Wu, J.S. Growth, photosynthetic and physiological responses of Torreya grandis seedlings to varied light environments. Trees 2015, 29, 1011–1022. [Google Scholar] [CrossRef]
- Huang, Z.; Liu, Q.; An, B.; Wu, X.; Sun, L.; Wu, P.; Liu, B.; Ma, X. Effects of Planting Density on Morphological and Photosynthetic Characteristics of Leaves in Different Positions on Cunninghamia lanceolata Saplings. Forests 2021, 12, 853. [Google Scholar] [CrossRef]
- Negi, S.; Barry, A.N.; Friedland, N.; Sudasinghe, N.; Subramanian, S.; Pieris, S.; Holguin, F.O.; Dungan, B.; Schaub, T.; Sayre, R. Impact of nitrogen limitation on biomass, photosynthesis, and lipid accumulation in Chlorella sorokiniana. J. Appl. Phycol. 2016, 28, 803–812. [Google Scholar] [CrossRef]
- Jiang, Y.; Ding, X.; Zhang, D.; Deng, Q.; Yu, C.L.; Zhou, S.; Hui, D. Soil salinity increases the tolerance of excessive sulfur fumigation stress in tomato plants. Environ. Exp. Bot. 2017, 133, 70–77. [Google Scholar] [CrossRef]
- Hichem, H.; Naceur, A.E.; Mounir, D. Effects of salt stress on photosynthesis, PSII photochemistry and thermal energy dissipation in leaves of two corn (Zea mays L.) varieties. Photosynthetica 2009, 47, 517–526. [Google Scholar] [CrossRef]
- Lin, Z.H.; Zhong, Q.S.; Chen, C.S.; Ruan, Q.C.; Chen, Z.H.; You, X.M. Carbon dioxide assimilation and photosynthetic electron transport of tea leaves under nitrogen deficiency. Bot. Stud. 2016, 57, 37. [Google Scholar] [CrossRef]
- Lin, Z.H.; Chen, L.S.; Chen, R.B.; Zhang, F.Z.; Jiang, H.X.; Tang, N. CO2 assimilation, ribulose-1,5-bisphosphate carboxylase/oxygenase, carbohydrates and photosynthetic electron transport probed by the JIP-test, of tea leaves in response to phosphorus supply. BMC Plant Biol. 2009, 9, 43. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll fluorescence analysis: A guide to good practice and understanding some new applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef] [PubMed]
- Hanachi, S.; Van Labeke, M.C.; Mehouachi, T. Application of chlorophyll fluorescence to screen eggplant (Solanum melongena L.) cultivars for salt tolerance. Photosynthetica 2014, 52, 57–62. [Google Scholar] [CrossRef]
- Hanelt, D. Photosynthesis Assessed by Chlorophyll Fluorescence; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Mathur, S.; Allakhverdiev, S.I.; Jajoo, A. Analysis of high temperature stress on the dynamics of antenna size and reducing side heterogeneity of Photosystem II in wheat leaves (Triticum aestivum). Biochim. Biophys. Acta (BBA) Bioenerg. 2011, 1807, 22–29. [Google Scholar] [CrossRef]
- Martins, S.C.; Galmés, J.; Molins, A.; DaMatta, F.M. Improving the estimation of mesophyll conductance to CO2: On the role of electron transport rate correction and respiration. J. Exp. Bot. 2013, 64, 3285–3298. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Varieties | Species | Parent | Breeding Units | Breeding Year |
|---|---|---|---|---|
| Ruidu Xiangyu | Eurasian | Jingxiu × Xiangfei | Institute of Forestry and Fruit Science, Beijing Academy of Agriculture and Forestry Science | In December 2007, it was approved by Beijing Forest Variety Examination and Approval Committee |
| Ruidu Cuixia | Eurasian | Jingxiu × Xiangfei | Institute of Forestry and Fruit Science, Beijing Academy of Agriculture and Forestry Science | In December 2007, it was approved by Beijing Forest Variety Examination and Approval Committee |
| Ruidu Zaohong | Eurasian | Jingxiu × Xiangfei | Institute of Forestry and Fruit Science, Beijing Academy of Agriculture and Forestry Science | In December 2014, it was approved by Beijing Forest Variety Examination and Approval Committee |
| Ruidu Wuheyi | Eurasian | Xiangfei × Hongbaoshi seedless | Institute of Forestry and Fruit Science, Beijing Academy of Agriculture and Forestry Science | In 2009, it was approved by the Beijing Forest Variety Examination and Approval Committee |
| Jinghongbao | Eurasian | Guibao × Wuhebai Jixin | Fruit research institute of Shanxi Academy of Agricultural Sciences | In 2012, it was approved by Shanxi Provincial Crop Variety Examination and Approval Committee |
| Varieties | Right Angle Hyperbolic Modified Model | Apparent Quantum Efficiency | Adjust R-Square | Dark Respiration Rate/ (μmol·m−2·s−1) | Light Saturation Point/ (μmol·m−2·s−1) | Light Compensation Point/(μmol·m−2·s−1) | Maximum Net Photosynthetic Rate/ (μmol·m−2·s−1) |
|---|---|---|---|---|---|---|---|
| Ruidu Xiangyu | y = 0.04488x− 0.85047 | 0.0298 | 0.991 | 0.85 | 1437.1 | 20 | 9.8 |
| Ruidu Cuixia | y = 0.05172x− 3.12694 | 0.0371 | 0.998 | 3.13 | 2290.5 | 66.8 | 19.2 |
| Ruidu Zaohong | y = 0.05921x− 2.69752 | 0.0384 | 0.995 | 2.70 | 2377.4 | 49.6 | 21.0 |
| Ruidu Wuheyi | y = 0.05966x− 2.02594 | 0.0391 | 0.995 | 2.03 | 1734.3 | 36.4 | 17.8 |
| Jing Hongbao | y = 0.05297x − 2.70940 | 0.0356 | 0.996 | 2.71 | 2171.3 | 55.4 | 20.4 |
| Principal Component Number | Eigenvalue | Rate of Contribution/% | Accumulating Contribution Rate/% |
|---|---|---|---|
| 1 | 10.9385 | 43.75% | 43.75% |
| 2 | 7.3320 | 29.33% | 73.08% |
| 3 | 3.5369 | 14.15% | 87.23% |
| 4 | 3.1926 | 12.77% | 100.00% |
| Varieties | F1 | F2 | F3 | F4 | F | Rank |
|---|---|---|---|---|---|---|
| Ruidu Xiangyu | −3.6070 | 2.3626 | −0.6990 | −1.8853 | −1.2248 | 4 |
| Ruidu Cuixia | 2.9872 | −1.5209 | 1.9314 | −1.8010 | 0.9041 | 2 |
| Ruidu Zaohong | 2.8615 | −0.7777 | −2.8697 | 0.3004 | 0.6561 | 3 |
| Ruidu Wuheyi | −3.4800 | −3.2592 | 0.4389 | 1.3790 | −2.2403 | 5 |
| Jinghongbao | 1.2383 | 3.1953 | 1.1984 | 2.0069 | 1.9048 | 1 |
| Indexes | F1 | F2 | F3 | F4 | F | Rank |
|---|---|---|---|---|---|---|
| Chlorophyll a | −0.0683 | 0.2336 | 0.3939 | −0.0271 | 0.0673 | 16 |
| Chlorophyll b | −0.0022 | 0.2730 | 0.2428 | 0.2771 | 0.1105 | 1 |
| Carotenoids | −0.1108 | −0.2200 | 0.1283 | −0.3760 | −0.1063 | 25 |
| Chlorophyll a + b | −0.0272 | 0.2729 | 0.3128 | 0.1776 | 0.1003 | 3 |
| Chlorophyll a/b | 0.0285 | −0.2691 | −0.1971 | −0.3176 | −0.1002 | 24 |
| Ci | 0.2296 | −0.1361 | 0.2000 | 0.2138 | 0.0863 | 12 |
| gs | 0.2706 | −0.0866 | 0.1834 | 0.0891 | 0.0968 | 5 |
| Pn | 0.2299 | −0.1105 | 0.0299 | 0.3205 | 0.0843 | 13 |
| Tr | 0.2424 | −0.0692 | 0.1815 | 0.2534 | 0.1069 | 2 |
| WUE | 0.0835 | −0.1826 | −0.3069 | 0.3293 | −0.0134 | 19 |
| Fv/Fm | −0.2076 | −0.2165 | 0.1546 | 0.1772 | −0.0816 | 22 |
| ΦPo | −0.2076 | −0.2165 | 0.1546 | 0.1772 | −0.0816 | 23 |
| ΦDo | 0.2076 | 0.2165 | −0.1546 | −0.1772 | 0.0816 | 14 |
| ΦPSII | −0.0033 | 0.3635 | 0.0179 | −0.0962 | 0.0708 | 15 |
| qp | 0.1554 | 0.2587 | 0.1377 | −0.2358 | 0.0988 | 4 |
| YNO | −0.1478 | 0.2805 | −0.2165 | 0.0768 | −0.0023 | 18 |
| YNPQ | 0.0621 | −0.3569 | 0.0759 | 0.0332 | −0.0465 | 21 |
| Fo | 0.2910 | 0.0678 | −0.0871 | −0.0637 | 0.0942 | 6 |
| Fm | 0.3011 | 0.0068 | −0.0422 | −0.0205 | 0.0930 | 8 |
| Fv | 0.3017 | −0.0156 | −0.0254 | −0.0046 | 0.0916 | 10 |
| ABS/CSm | 0.3011 | 0.0068 | −0.0422 | −0.0205 | 0.0930 | 9 |
| Tro/CSm | 0.3017 | −0.0156 | −0.0254 | −0.0046 | 0.0916 | 11 |
| DIo/CSm | 0.2910 | 0.0678 | −0.0871 | −0.0637 | 0.0942 | 7 |
| Specific leaf area | 0.0419 | −0.1743 | 0.4141 | −0.2195 | −0.0019 | 17 |
| Specific leaf weight | −0.1528 | 0.1154 | −0.3097 | 0.3106 | −0.0273 | 20 |
| Indexes | Positive Ideal Solution A+ | Negative Ideal Solution A− | Indexes | Positive Ideal Solution A+ | Negative Ideal Solution A− |
|---|---|---|---|---|---|
| Chlorophyll a | 0.492 | 0.38 | ΦPo | 0.457 | 0.443 |
| Chlorophyll b | 0.695 | 0.291 | ΦDo | 0.465 | 0.409 |
| Carotenoids | 0.506 | 0.344 | ΦPSII | 0.513 | 0.356 |
| Chlorophyll a + b | 0.563 | 0.353 | qp | 0.482 | 0.364 |
| Chlorophyll a/b | 0.513 | 0.276 | YNO | 0.523 | 0.349 |
| Ci | 0.499 | 0.341 | YNPQ | 0.518 | 0.355 |
| gs | 0.63 | 0.135 | Fo | 0.508 | 0.357 |
| Pn | 0.521 | 0.237 | Fm | 0.492 | 0.391 |
| Tr | 0.521 | 0.269 | Fv | 0.487 | 0.397 |
| WUE | 0.503 | 0.397 | ABS/CSm | 0.492 | 0.391 |
| Specific leaf area | 0.462 | 0.424 | Tro/CSm | 0.487 | 0.397 |
| Specific leaf weight | 0.453 | 0.424 | DIo/CSm | 0.508 | 0.357 |
| Fv/Fm | 0.457 | 0.443 |
| Varieties | Positive Ideal Solution Distance D+ | Negative Ideal Solution Distance D− | Degree of Relative Proximity C | Sorting Result |
|---|---|---|---|---|
| Ruidu Xiangyu | 0.779 | 0.389 | 0.333 | 5 |
| Ruidu Cuixia | 0.434 | 0.769 | 0.639 | 2 |
| Ruidu Zaohong | 0.53 | 0.663 | 0.556 | 3 |
| Ruidu Wuheyi | 0.624 | 0.464 | 0.426 | 4 |
| Jing Hongbao | 0.367 | 0.78 | 0.68 | 1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, Y.; Yadav, V.; Bai, S.; Wu, J.; Zhou, X.; Zhang, W.; Han, S.; Wang, M.; Zeng, B.; Wu, X.; et al. Performance Evaluation of New Table Grape Varieties under High Light Intensity Conditions Based on the Photosynthetic and Chlorophyll Fluorescence Characteristics. Horticulturae 2023, 9, 1035. https://doi.org/10.3390/horticulturae9091035
He Y, Yadav V, Bai S, Wu J, Zhou X, Zhang W, Han S, Wang M, Zeng B, Wu X, et al. Performance Evaluation of New Table Grape Varieties under High Light Intensity Conditions Based on the Photosynthetic and Chlorophyll Fluorescence Characteristics. Horticulturae. 2023; 9(9):1035. https://doi.org/10.3390/horticulturae9091035
Chicago/Turabian StyleHe, Yawen, Vivek Yadav, Shijian Bai, Jiuyun Wu, Xiaoming Zhou, Wen Zhang, Shouan Han, Min Wang, Bin Zeng, Xinyu Wu, and et al. 2023. "Performance Evaluation of New Table Grape Varieties under High Light Intensity Conditions Based on the Photosynthetic and Chlorophyll Fluorescence Characteristics" Horticulturae 9, no. 9: 1035. https://doi.org/10.3390/horticulturae9091035