

Anthocyanin Accumulation and Its Corresponding Gene Expression, Total Phenol, Antioxidant Capacity, and Fruit Quality of ‘Crimson Seedless’ Grapevine (Vitis vinifera L.) in Response to Grafting and Pre-Harvest Applications
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Evaluated Treatments
2.2. Measurements and Analysis
2.2.1. Fruit Yield and Physical Attributes
2.2.2. Berry Color
2.2.3. Soluble Solids Content (SSC) and Titratable Acidity (TA)
2.2.4. Total Phenols and Antioxidative Capacity
2.2.5. Anthocyanins Accumulation and Content
2.2.6. Determination of Enzymes Activities
2.2.7. RNA Extraction, cDNA Synthesis, and Quantitative Real-Time PCR
2.3. Statistical Analysis
3. Results
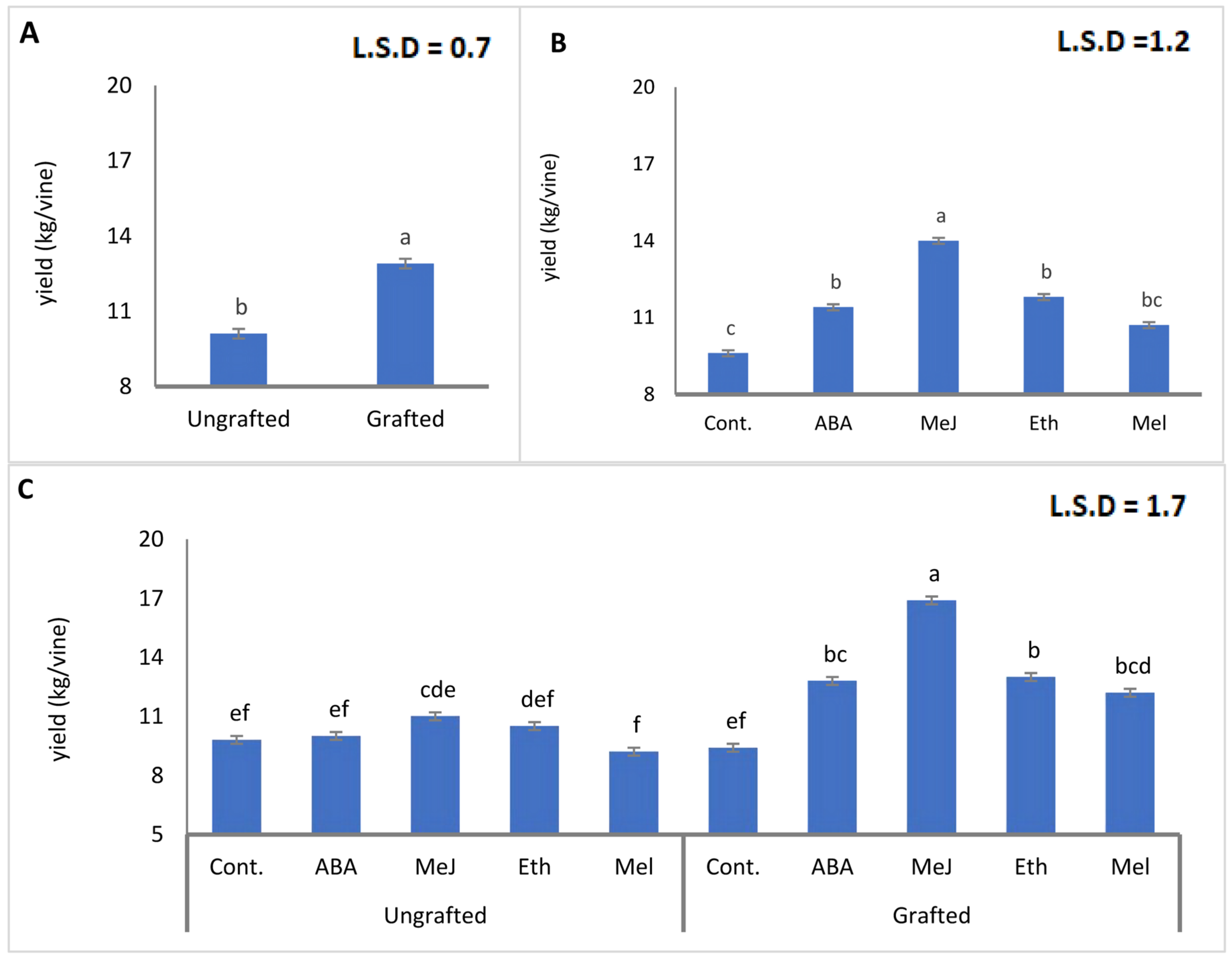
3.1. Yield (kg/Vine)
3.2. Bunch Weight, Weight, and Size of 50 Berries
3.3. Berry Firmness, Removal Force, and Berry Diameter
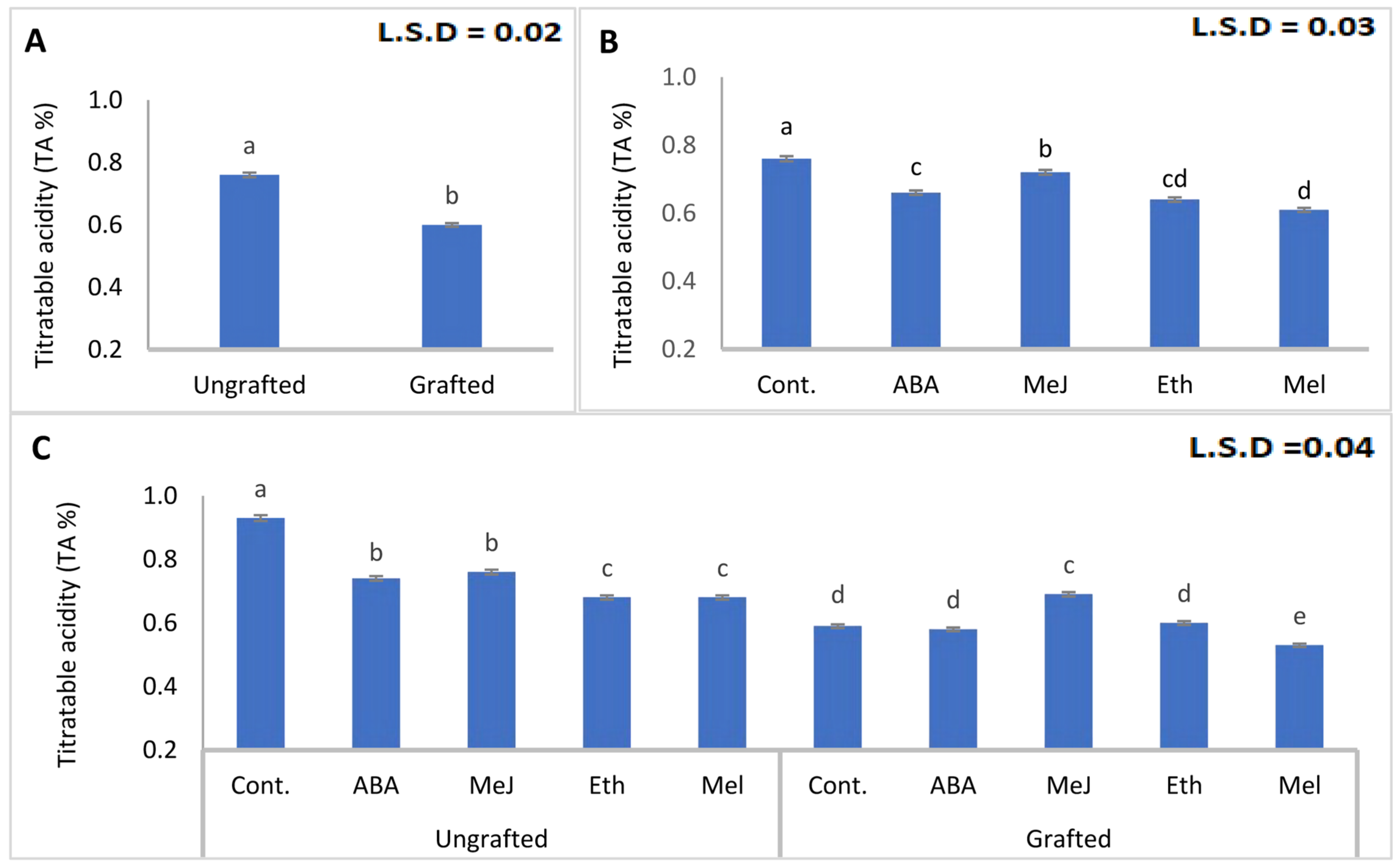
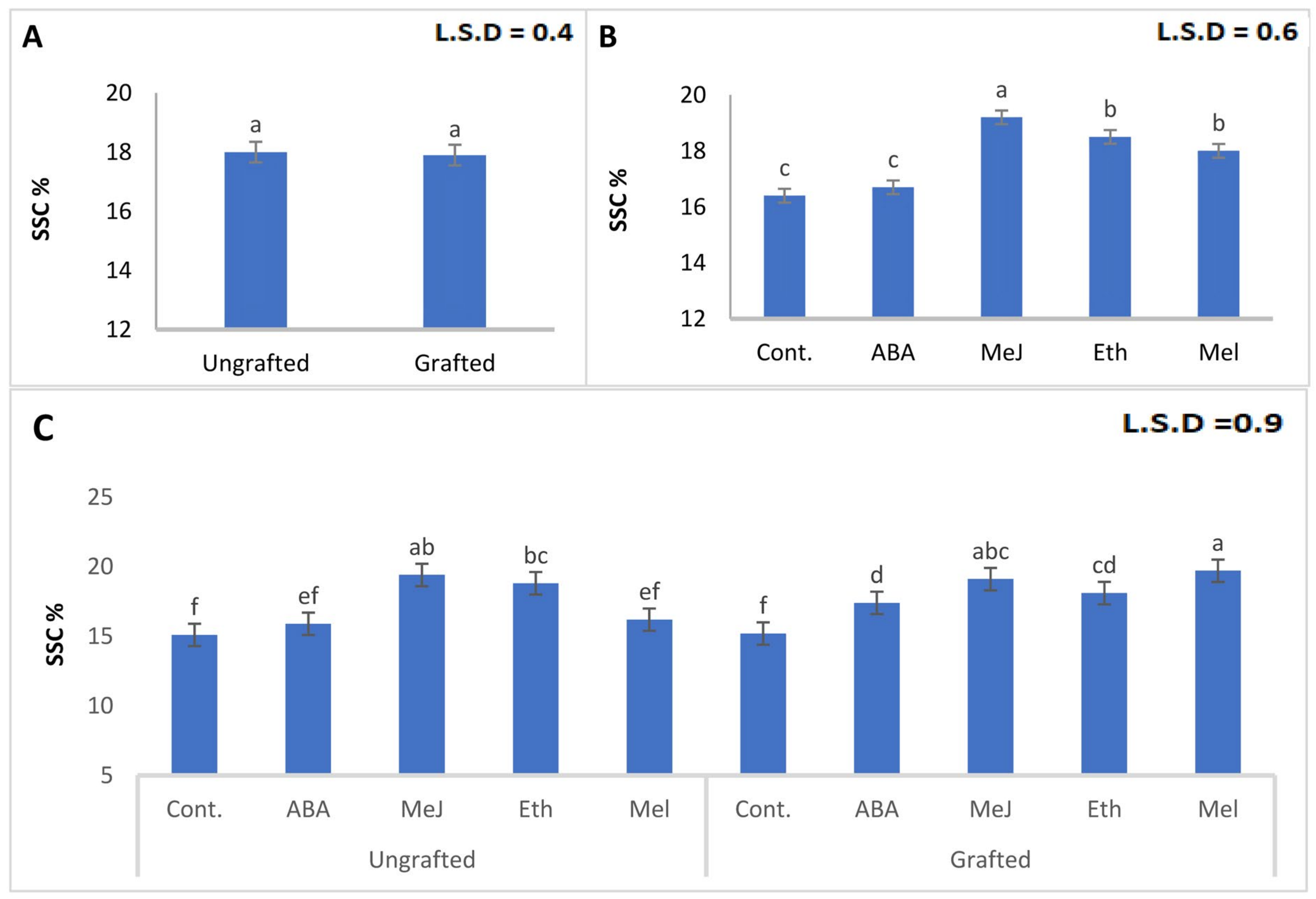
3.4. Titratable Acidity and Soluble Solids Content
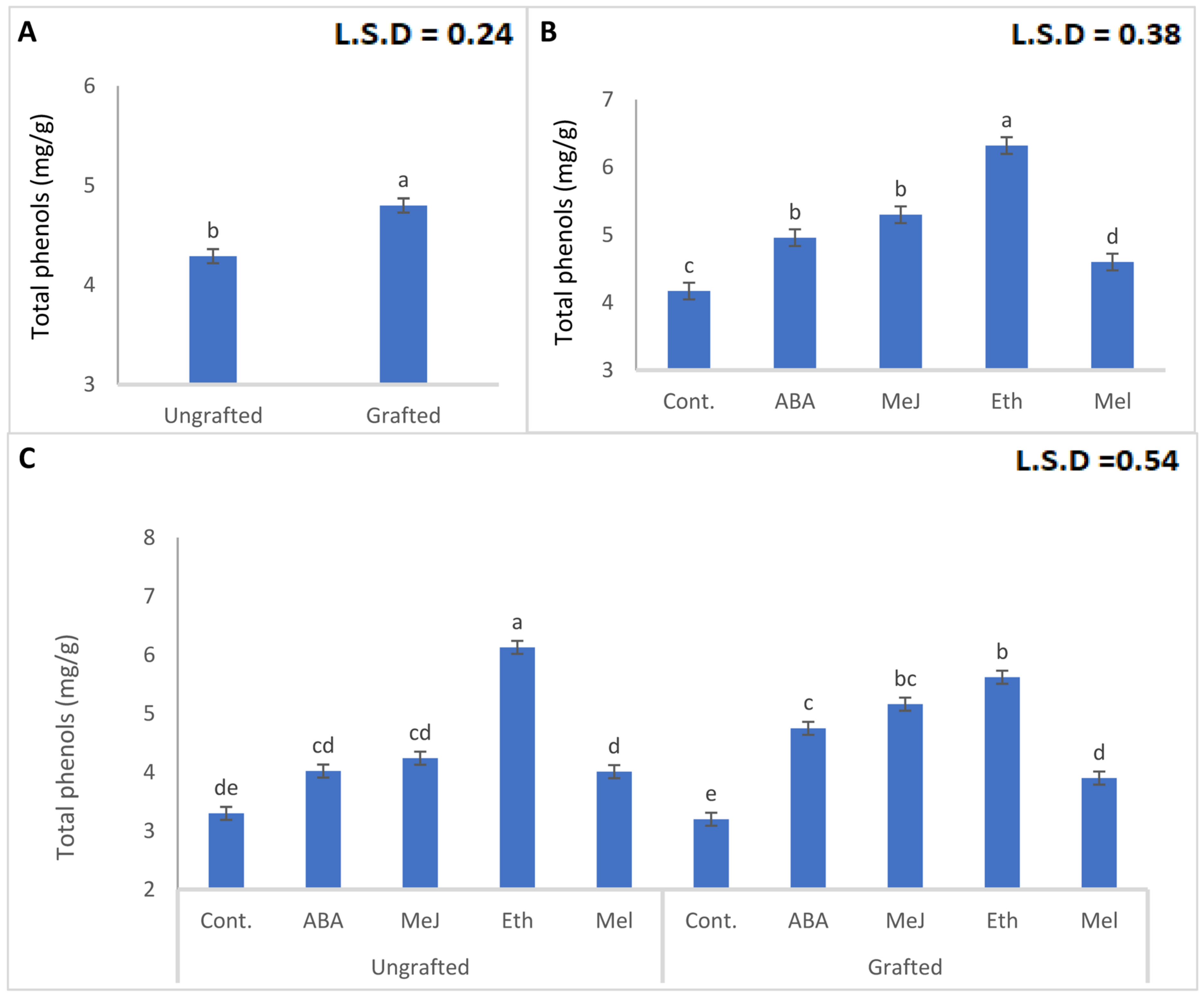
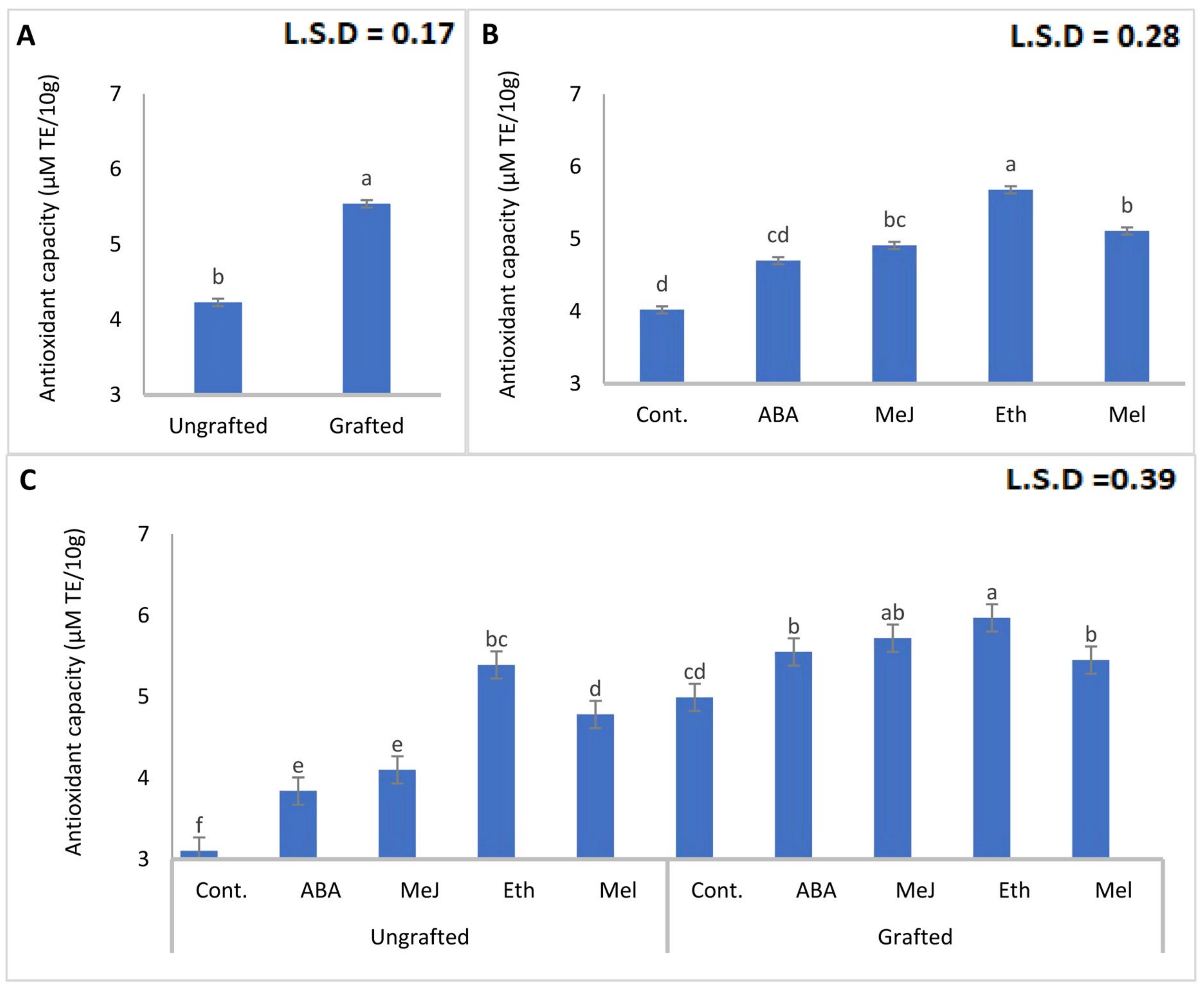
3.5. Total Phenols and Antioxidant Capacity
3.6. Color
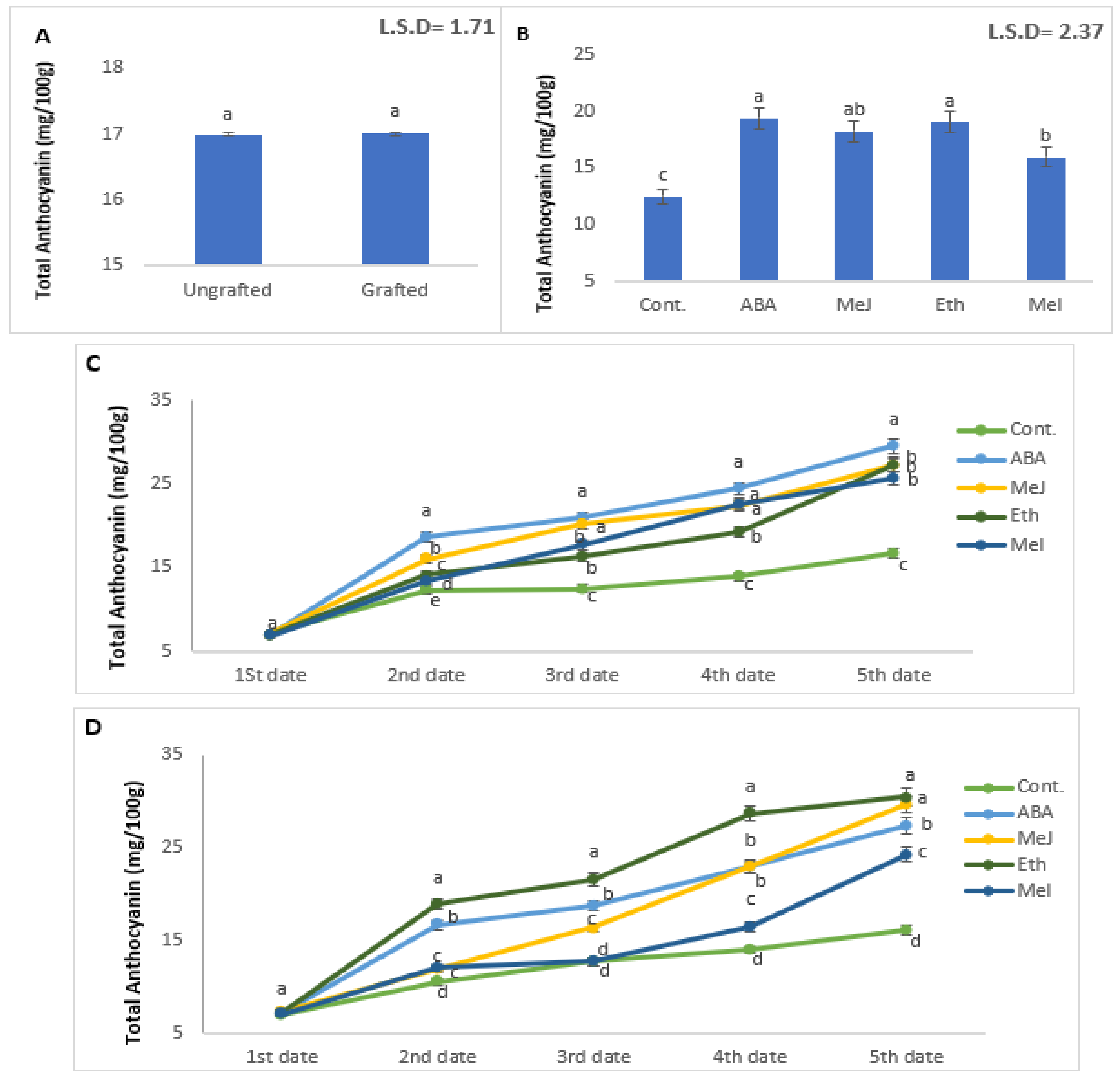
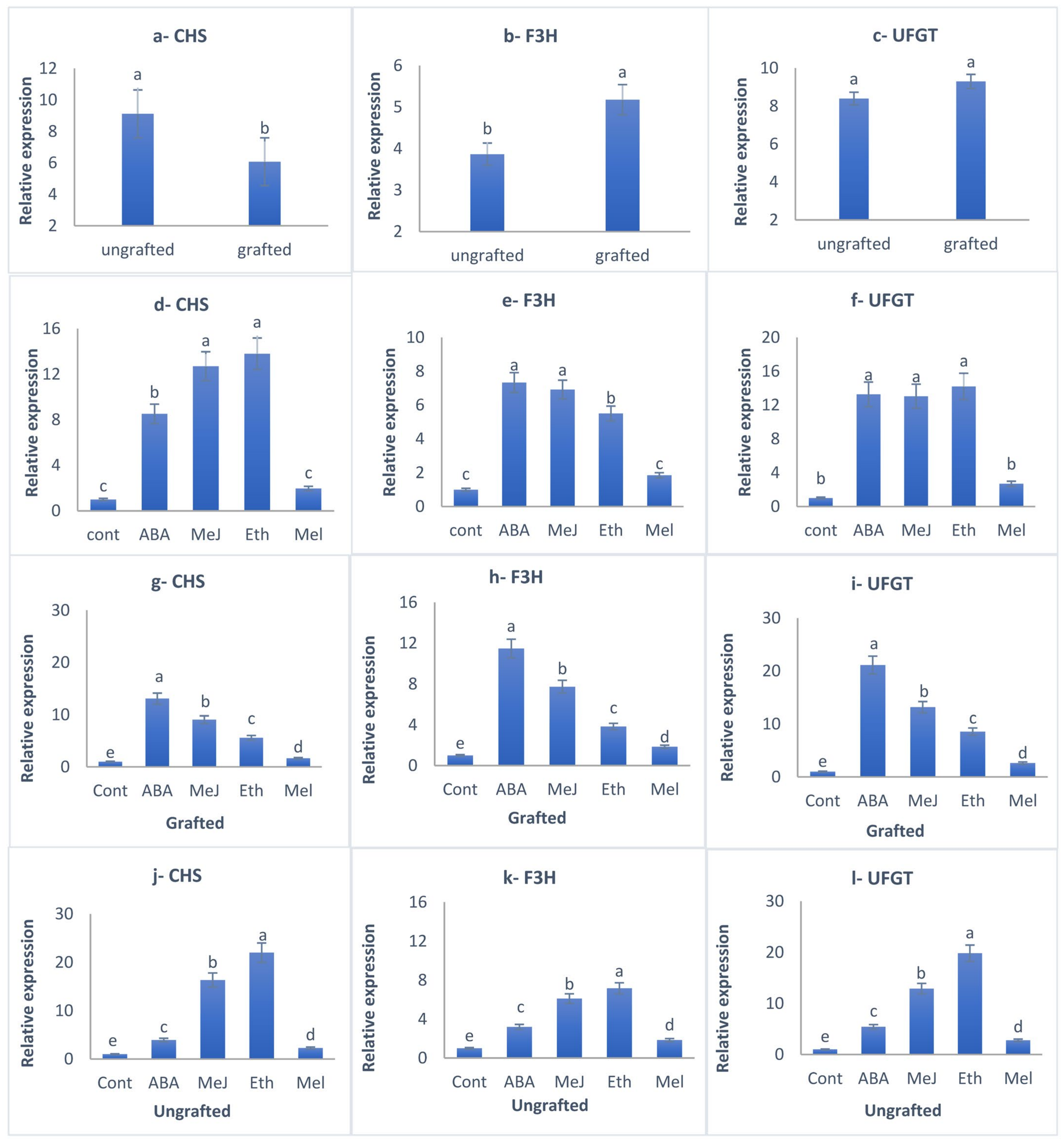
3.7. Total Anthocyanin Accumulation and Gene Expression
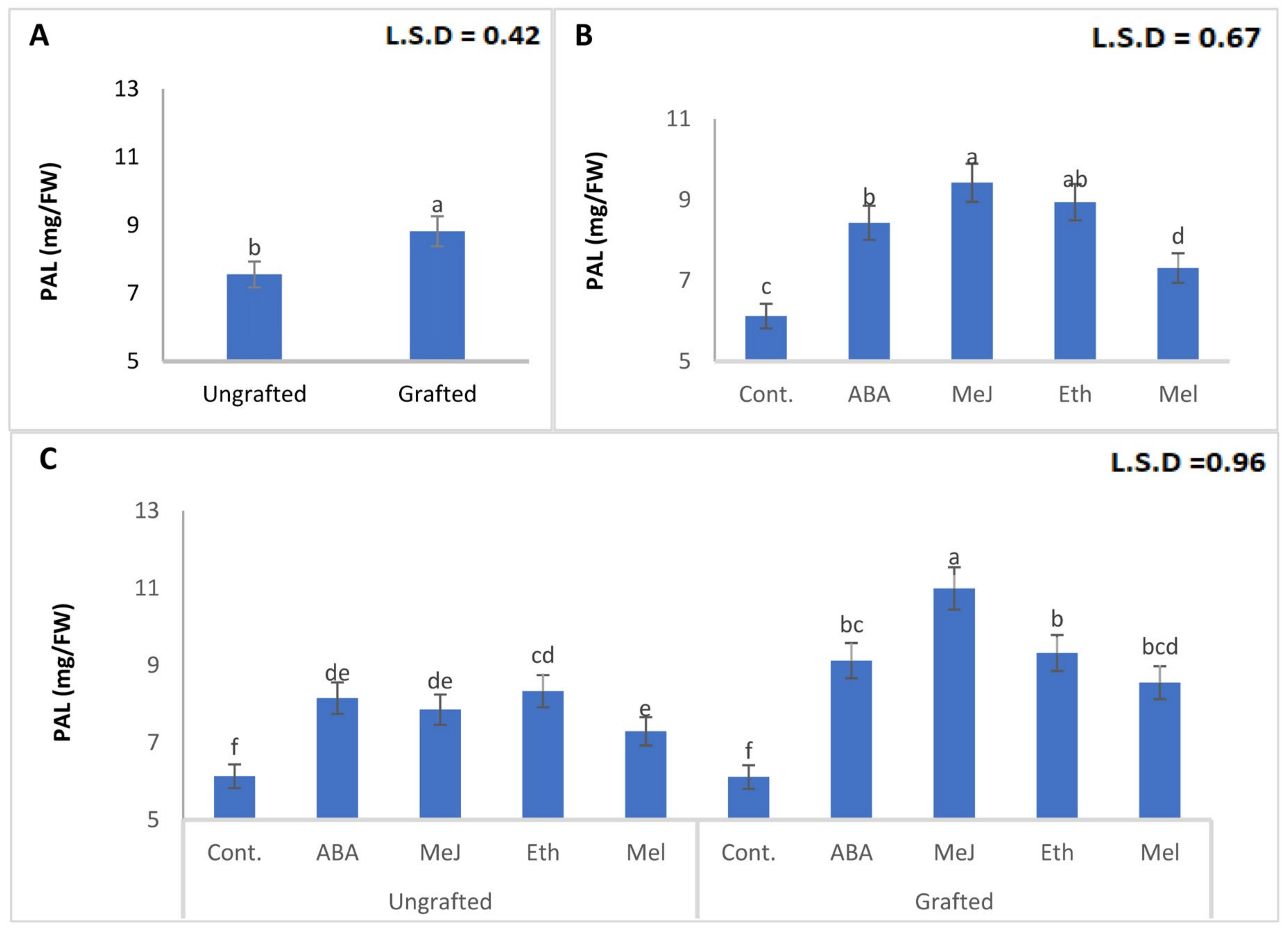
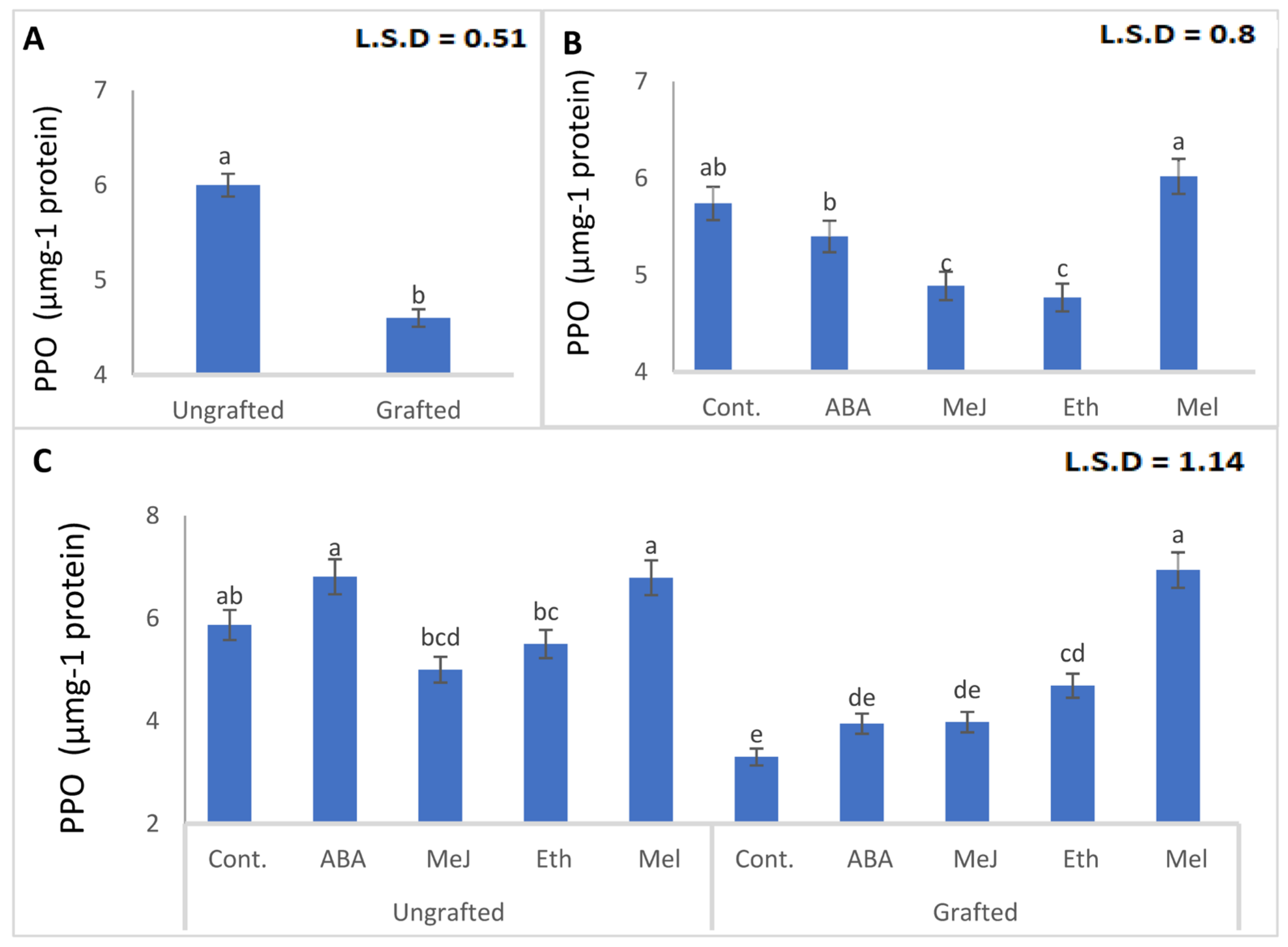
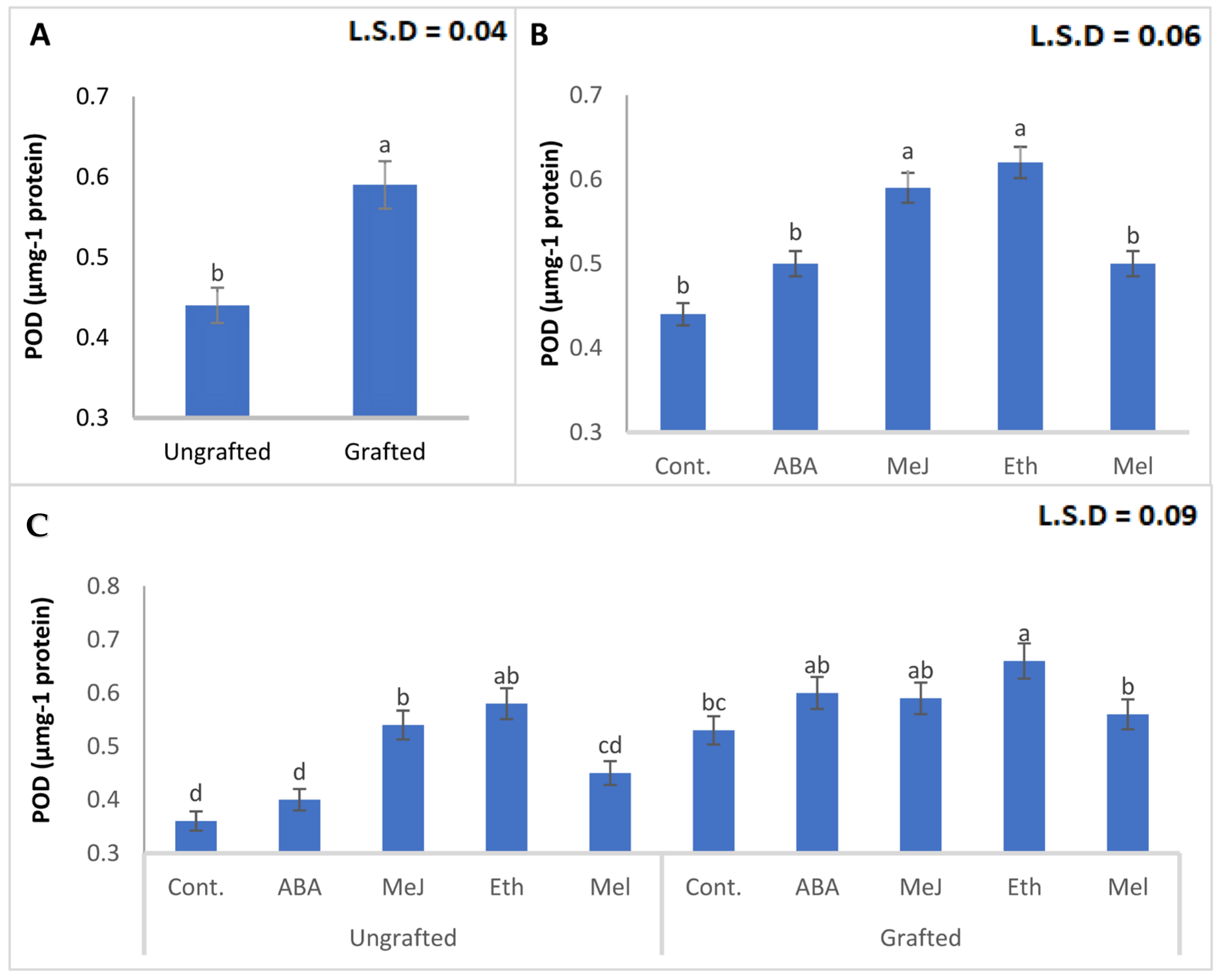
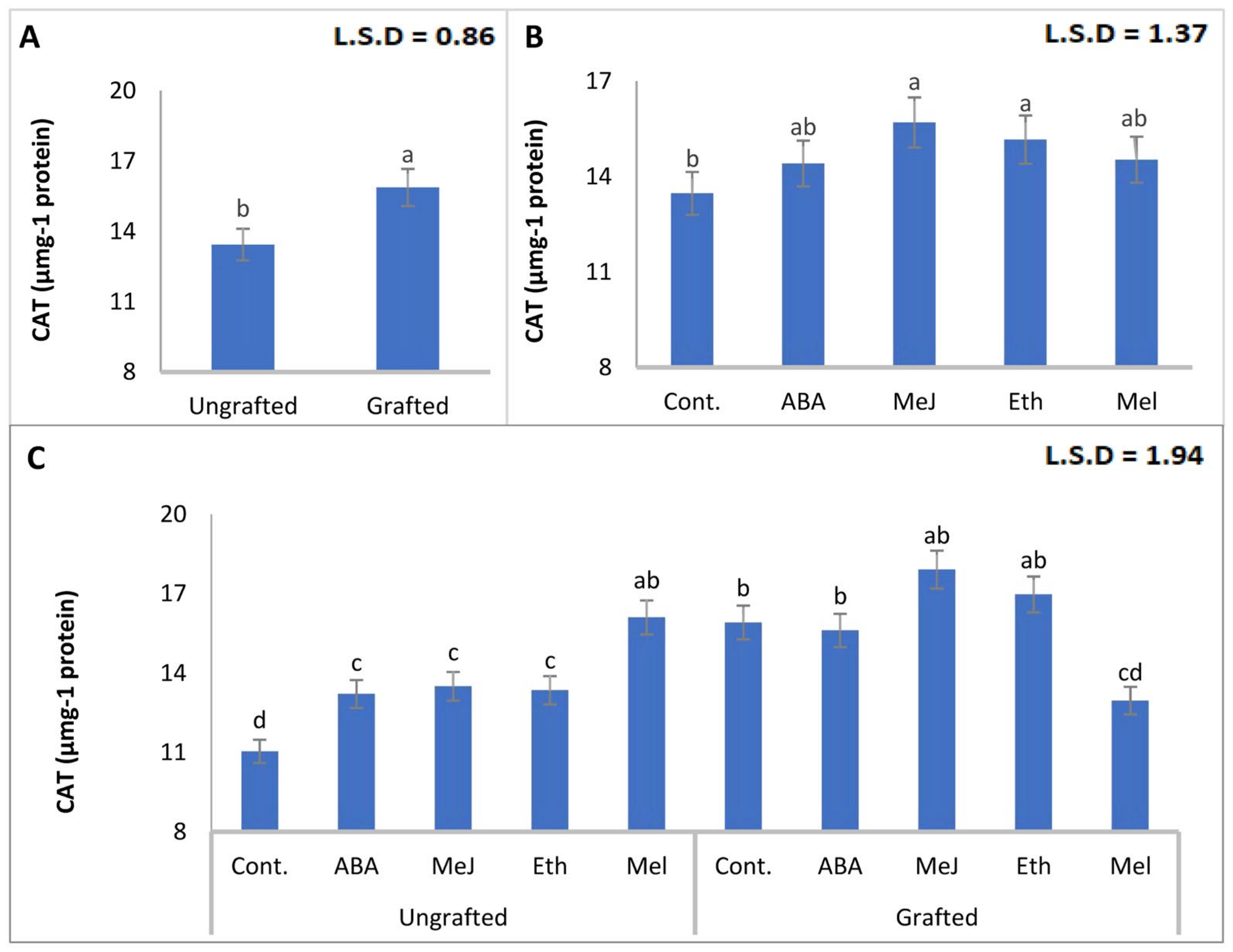
3.8. Enzymes (PAL, PPO, POD, and CAT)
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Food and Agriculture Organisation of the United Nations. Crops and Livestock Products. 2021. Available online: http://www.fao.org/faostat/en/#data/qc (accessed on 30 July 2023).
- Domingues Neto, F.J.; Pimentel Junior, A.; Borges, C.V.; Cunha, S.R.; Callili, D.; Lima, G.P.P.; Roberto, S.R.; Leonel, S.; Tecchio, M.A. The Exogenous Application of Abscisic Acid Induce Accumulation of Anthocyanins and Phenolic Compounds of the ‘Rubi’ Grape. Am. J. Plant Sci. 2017, 8, 2422–2432. [Google Scholar] [CrossRef]
- Arias, I.; Lacau, B.; Astrain, J.; Barón, C.; Fenandez-Zurbano, P.; Ferreira, V.; Escudero, A. Effects of Vineyard ‘Potential’ and Grape Maturation on the Aroma-Volatile Profile of Grenache Wines. Oeno One 2019, 53, 695–707. [Google Scholar] [CrossRef]
- Bouzas-Cid, Y.; Falqué, E.; Orriols, I.; Mirás-Avalos, J.M. Effects of Irrigation over Three Years on the Amino Acid Composition of Treixadura (Vitis Vinifera L.) Musts and Wines, and on the Aromatic Composition and Sensory Profiles of Its Wines. Food Chem. 2018, 240, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.H.; Wang, B.L.; Wang, X.D.; Shi, X.B.; Liu, P.P.; Liu, F.Z.; Wang, H.B. Effects of Different Color Paper Bags on Aroma Development of Kyoho Grape Berries. J. Integr. Agric. 2019, 18, 70–82. [Google Scholar] [CrossRef]
- Suehiro, Y.; Mochida, K.; Tsuma, M.; Yasuda, Y.; Itamura, H.; Esumi, T. Effects of Gibberellic Acid/Cytokinin Treatments on Berry Development and Maturation in the Yellow-Green Skinned ‘Shine Muscat’ Grape. Hortic. J. 2019, 88, 202–213. [Google Scholar] [CrossRef]
- Chong, Z.; Hao, C.; Lianjun, W. Effect of ABA Treatment on the Expression of ADH Gene Family and C6 Volatile Production in Table Grape (V. Vinifera Cv. Muscat Hamburg) during Postharvest Storage. Acta Physiol. Plant. 2020, 42, 55. [Google Scholar] [CrossRef]
- Gao, X.T.; Wu, M.H.; Sun, D.; Li, H.Q.; Chen, W.K.; Yang, H.Y.; Liu, F.Q.; Wang, Q.C.; Wang, Y.Y.; Wang, J.; et al. Effects of Gibberellic Acid (GA3) Application before Anthesis on Rachis Elongation and Berry Quality and Aroma and Flavour Compounds in Vitis vinifera L. ‘Cabernet Franc’ and ‘Cabernet Sauvignon’ Grapes. J. Sci. Food Agric. 2020, 100, 3729–3740. [Google Scholar] [CrossRef]
- Xi, X.; Zha, Q.; He, Y.; Tian, Y.; Jiang, A. Influence of Cluster Thinning and Girdling on Aroma Composition in ‘Jumeigui’ Table Grape. Sci. Rep. 2020, 10, 6877. [Google Scholar] [CrossRef]
- Downey, M.O.; Dokoozlian, N.K.; Krstic, M.P. Cultural Practice and Environmental Impacts on the Flavonoid Composition of Grapes and Wine: A Review of Recent Research. Am. J. Enol. Vitic. 2006, 57, 257–268. [Google Scholar] [CrossRef]
- Kuhn, N.; Guan, L.; Dai, Z.W.; Wu, B.H.; Lauvergeat, V.; Gomès, E.; Li, S.H.; Godoy, F.; Arce-Johnson, P.; Delrot, S. Berry Ripening: Recently Heard through the Grapevine. J. Exp. Bot. 2014, 65, 4543–4559. [Google Scholar] [CrossRef]
- Nakazawa, A.; Nozue, M.; Yasuda, H.; Takeba, G.; Kubo, H. Expression Pattern and Gene Structure of Phenylalanine Ammonia-Lyase in Pharbitis Nil. J. Plant Res. 2001, 114, 323–328. [Google Scholar] [CrossRef]
- Jaakola, L. New Insights into the Regulation of Anthocyanin Biosynthesis in Fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef]
- Sicilia, A.; Scialò, E.; Puglisi, I.; Lo Piero, A.R. Anthocyanin Biosynthesis and DNA Methylation Dynamics in Sweet Orange Fruit [Citrus sinensis L. (Osbeck)] under Cold Stress. J. Agric. Food Chem. 2020, 68, 7024–7031. [Google Scholar] [CrossRef]
- Wen, W.; Alseekh, S.; Fernie, A.R. Conservation and Diversification of Flavonoid Metabolism in the Plant Kingdom. Curr. Opin. Plant Biol. 2020, 55, 100–108. [Google Scholar] [CrossRef] [PubMed]
- Tian, M.; Ma, W.; Xia, N.; Peng, J.; Hu, R.; Duan, C.; He, F. Soil Variables and Reflected Light Revealed the Plasticity of Grape and Wine Composition: Regulation of the Flavoromics under Inner Row Gravel Covering. Food Chem. 2023, 414, 135659. [Google Scholar] [CrossRef]
- Wang, Y.U.; Li, H.Q.; Gao, X.T.; Lu, H.C.; Peng, W.T.; Chen, W.U.; Li, S.P.; Duan, C.Q.; Wang, J. Influence of Attenuated Reflected Solar Radiation from the Vineyard Floor on Volatile Compounds in Cabernet Sauvignon Grapes and Wines of the North Foot of Mt. Tianshan. Food Res. Int. 2020, 137, 109688. [Google Scholar] [CrossRef]
- Samaan, M.; Nasser, M.A. Effect of Spraying Paclobutrazol (PP333) on Yield and Fruit Quality of Crimson Seedless Grape. J. Plant Prod. 2020, 11, 1031–1034. [Google Scholar] [CrossRef]
- Peppi, M.C.; Fidelibus, M.W. Effects of Forchlorfenuron and Abscisic Acid on the Quality of “Flame Seedless” Grapes. HortScience 2008, 43, 173–176. [Google Scholar] [CrossRef]
- Setha, S. Roles of Abscisic Acid in Fruit Ripening. Walailak J. Sci. Technol. 2012, 9, 297–308. [Google Scholar]
- Yang, F.W.; Feng, X.Q. Abscisic Acid Biosynthesis and Catabolism and Their Regulation Roles in Fruit Ripening. Phyton-Int. J. Exp. Bot. 2015, 84, 444–453. [Google Scholar] [CrossRef]
- Marzouk, H.A.; Kassem, H.A. Improving Fruit Quality, Nutritional Value and Yield of Zaghloul Dates by the Application of Organic and/or Mineral Fertilizers. Sci. Hortic. 2011, 127, 249–254. [Google Scholar] [CrossRef]
- Rattanakon, S.; Ghan, R.; Gambetta, G.A.; Deluc, L.G.; Schlauch, K.A.; Cramer, G.R. Abscisic Acid Transcriptomic Signaling Varies with Grapevine Organ. BMC Plant Biol. 2016, 16, 72. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Butelli, E.; Martin, C. Engineering Anthocyanin Biosynthesis in Plants. Curr. Opin. Plant Biol. 2014, 19, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Ferrara, G.; Mazzeo, A.; Matarrese, A.M.S.; Pacucci, C.; Punzi, R.; Faccia, M.; Trani, A.; Gambacorta, G. Application of Abscisic Acid (S-ABA) and Sucrose to Improve Colour, Anthocyanin Content and Antioxidant Activity of Cv. Crimson Seedless Grape Berries. Aust. J. Grape Wine Res. 2015, 21, 18–29. [Google Scholar] [CrossRef]
- Olivares, D.; Contreras, C.; Muñoz, V.; Rivera, S.; González-Agüero, M.; Retamales, J.; Defilippi, B.G. Relationship among Color Development, Anthocyanin and Pigment-Related Gene Expression in ‘Crimson Seedless’ Grapes Treated with Abscisic Acid and Sucrose. Plant Physiol. Biochem. 2017, 115, 286–297. [Google Scholar] [CrossRef]
- Dar, T.A.; Uddin, M.; Khan, M.M.A.; Hakeem, K.R.; Jaleel, H. Jasmonates Counter Plant Stress: A Review. Environ. Exp. Bot. 2015, 115, 49–57. [Google Scholar] [CrossRef]
- Portu, J.; López, R.; Santamaría, P.; Garde-Cerdán, T. Methyl Jasmonate Treatment to Increase Grape and Wine Phenolic Content in Tempranillo and Graciano Varieties during Two Growing Seasons. Sci. Hortic. 2018, 240, 378–386. [Google Scholar] [CrossRef]
- Gómez-Plaza, E.; Bautista-Ortín, A.B.; Ruiz-García, Y.; Fernández-Fernández, J.I.; Gil-Muñoz, R. Effect of Elicitors on the Evolution of Grape Phenolic Compounds during the Ripening Period. J. Sci. Food Agric. 2017, 97, 977–983. [Google Scholar] [CrossRef]
- Bellincontro, A.; Fardelli, A.; De Santis, D.; Botondi, R.; Mencarelli, F. Postharvest Ethylene and 1-MCP Treatments Both Affect Phenols, Anthocyanins, and Aromatic Quality of Aleatico Grapes and Wine. Aust. J. Grape Wine Res. 2006, 12, 141–149. [Google Scholar] [CrossRef]
- An, J.P.; Wang, X.F.; Li, Y.Y.; Song, L.Q.; Zhao, L.L.; You, C.X.; Hao, Y.J. EIN3-LIKE1, MYB1, and Ethylene Response Factor3 Act in a Regulatory Loop That Synergistically Modulates Ethylene Biosynthesis and Anthocyanin Accumulation. Plant Physiol. 2018, 178, 808–823. [Google Scholar] [CrossRef]
- Cheng, Y.; Liu, L.; Yuan, C.; Guan, J. Molecular Characterization of Ethylene-Regulated Anthocyanin Biosynthesis in Plums During Fruit Ripening. Plant Mol. Biol. Report. 2016, 34, 777–785. [Google Scholar] [CrossRef]
- An, J.P.; Yao, J.F.; Xu, R.R.; You, C.X.; Wang, X.F.; Hao, Y.J. Apple BZIP Transcription Factor MdbZIP44 Regulates Abscisic Acid-Promoted Anthocyanin Accumulation. Plant Cell Environ. 2018, 41, 2678–2692. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Yang, S.; Ni, J.; Teng, Y.; Bai, S. Advances of Anthocyanin Synthesis Regulated by Plant Growth Regulators in Fruit Trees. Sci. Hortic. 2023, 307, 111476. [Google Scholar] [CrossRef]
- Ni, J.B.; Bai, S.L.; Zhao, Y.; Tao, R.Y.; Teng, Y.W. Ethylene Suppresses Anthocyanin Accumulation in “Red Zaosu” Pear. In Proceedings of the Acta Horticulturae; International Society for Horticultural Science (ISHS): Leuven, Belgium, 2020; pp. 513–520. [Google Scholar]
- Figueroa, N.E.; Gatica-Meléndez, C.; Figueroa, C.R. Ethylene Application at the Immature Stage of Fragaria Chiloensis Fruit Represses the Anthocyanin Biosynthesis with a Concomitant Accumulation of Lignin. Food Chem. 2021, 358, 129913. [Google Scholar] [CrossRef] [PubMed]
- Arnao, M.B.; Hernández-Ruiz, J. Functions of Melatonin in Plants: A Review. J. Pineal Res. 2015, 59, 133–150. [Google Scholar] [CrossRef]
- Zhang, H.M.; Zhang, Y. Melatonin: A Well-Documented Antioxidant with Conditional pro-Oxidant Actions. J. Pineal Res. 2014, 57, 131–146. [Google Scholar] [CrossRef]
- Murch, S.J.; Hall, B.A.; Le, C.H.; Saxena, P.K. Changes in the Levels of Indoleamine Phytochemicals during Véraison and Ripening of Wine Grapes. J. Pineal Res. 2010, 49, 95–100. [Google Scholar] [CrossRef]
- Meng, J.F.; Yu, Y.; Shi, T.C.; Fu, Y.S.; Zhao, T.; Zhang, Z.W. Melatonin Treatment of Pre-Veraison Grape Berries Modifies Phenolic Components and Antioxidant Activity of Grapes and Wine. Food Sci. Technol. 2019, 39, 35–42. [Google Scholar] [CrossRef]
- Rouphael, Y.; Kyriacou, M.C.; Colla, G. Vegetable Grafting: A Toolbox for Securing Yield Stability under Multiple Stress Conditions. Front. Plant Sci. 2018, 8, 10–13. [Google Scholar] [CrossRef]
- Kumar, R.M.S.; Ji, G.; Guo, H.; Zhao, L.; Zheng, B. Over-Expression of a Grafting-Responsive Gene from Hickory Increases Abiotic Stress Tolerance in Arabidopsis. Plant Cell Rep. 2018, 37, 541–552. [Google Scholar] [CrossRef]
- Lana, G.; Modica, G.; Las Casas, G.; Siracusa, L.; La Malfa, S.; Gentile, A.; Sicilia, A.; Distefano, G.; Continella, A. Molecular Insights into the Effects of Rootstocks on Maturation of Blood Oranges. Horticulturae 2021, 7, 468. [Google Scholar] [CrossRef]
- Parvaneh, T.; Abedi, B.; Davarynejad, G.H.; Ganji Moghadam, E. Enzyme Activity, Phenolic and Flavonoid Compounds in Leaves of Iranian Red Flesh Apple Cultivars Grown on Different Rootstocks. Sci. Hortic. 2019, 246, 862–870. [Google Scholar] [CrossRef]
- Font i Forcada, C.; Gogorcena, Y.; Moreno, M.A. Fruit Sugar Profile and Antioxidants of Peach and Nectarine Cultivars on Almond×peach Hybrid Rootstocks. Sci. Hortic. 2013, 164, 563–572. [Google Scholar] [CrossRef]
- Milošević, T.; Milošević, N.; Mladenović, J. Combining Fruit Quality and Main Antioxidant Attributes in the Sour Cherry: The Role of New Clonal Rootstock. Sci. Hortic. 2020, 265, 109236. [Google Scholar] [CrossRef]
- Cheng, J.; Wei, L.; Mei, J.; Wu, J. Effect of Rootstock on Phenolic Compounds and Antioxidant Properties in Berries of Grape (Vitis vinifera L.) Cv. ‘Red Alexandria’. Sci. Hortic. 2017, 217, 137–144. [Google Scholar] [CrossRef]
- Bascuñán-Godoy, L.; Franck, N.; Zamorano, D.; Sanhueza, C.; Carvajal, D.E.; Ibacache, A. Rootstock Effect on Irrigated Grapevine Yield under Arid Climate Conditions Are Explained by Changes in Traits Related to Light Absorption of the Scion. Sci. Hortic. 2017, 218, 284–292. [Google Scholar] [CrossRef]
- Da Silva, M.J.R.; Vedoato, B.T.F.; Lima, G.P.P.; Moura, M.F.; Coser, G.M.D.A.G.; Watanabe, C.Y.; Tecchio, M.A. Phenolic Compounds and Antioxidant Activity of Red and White Grapes on Different Rootstocks. Afr. J. Biotechnol. 2017, 16, 664–671. [Google Scholar] [CrossRef]
- Balic, I.; Ejsmentewicz, T.; Sanhueza, D.; Silva, C.; Peredo, T.; Olmedo, P.; Barros, M.; Verdonk, J.C.; Paredes, R.; Meneses, C.; et al. Biochemical and Physiological Study of the Firmness of Table Grape Berries. Postharvest Biol. Technol. 2014, 93, 15–23. [Google Scholar] [CrossRef]
- Lancaster, J.E.; Lister, C.E.; Reay, P.F.; Triggs, C.M. Influence of Pigment Composition on Skin Color in a Wide Range of Fruit and Vegetables. J. Am. Soc. Hortic. Sci. 1997, 122, 594–598. [Google Scholar] [CrossRef]
- Chironi, S.; Sortino, G.; Allegra, A.; Saletta, F.; Caviglia, V.; Ingrassia, M. Consumer Assessment on Sensory Attributes of Fresh Table Grapes Cv “Italia” and “Red Globe” after Long Cold Storage Treatment. Chem. Eng. Trans. 2017, 58, 421–426. [Google Scholar] [CrossRef]
- Martínez-Esplá, A.; Zapata, P.J.; Castillo, S.; Guillén, F.; Martínez-Romero, D.; Valero, D.; Serrano, M. Preharvest Application of Methyl Jasmonate (MeJA) in Two Plum Cultivars. 1. Improvement of Fruit Growth and Quality Attributes at Harvest. Postharvest Biol. Technol. 2014, 98, 98–105. [Google Scholar] [CrossRef]
- Binsan, W.; Benjakul, S.; Visessanguan, W.; Roytrakul, S.; Tanaka, M.; Kishimura, H. Antioxidative Activity of Mungoong, an Extract Paste, from the Cephalothorax of White Shrimp (Litopenaeus vannamei). Food Chem. 2008, 106, 185–193. [Google Scholar] [CrossRef]
- Von Wettstein, D. Chlorophyll-Letale Und Der Submikroskopische Formwechsel Der Plastiden. Exp. Cell Res. 1957, 12, 427–506. [Google Scholar] [CrossRef] [PubMed]
- Assis, J.S.; Maldonado, R.; Muñoz, T.; Escribano, M.I.; Merodio, C. Effect of High Carbon Dioxide Concentration on PAL Activity and Phenolic Contents in Ripening Cherimoya Fruit. Postharvest Biol. Technol. 2001, 23, 33–39. [Google Scholar] [CrossRef]
- Kar, M.; Mishra, D. Catalase, Peroxidase, and Polyphenoloxidase Activities during Rice Leaf Senescence. Plant Physiol. 1976, 57, 315–319. [Google Scholar] [CrossRef]
- Chance, B.; Meahly, A.C.I. Catalase Assay by Disappearance of Peroxide. Methods Enzymol. 1955, 2, 764–775. [Google Scholar]
- Singh, R.K.; Martins, V.; Soares, B.; Castro, I.; Falco, V. Chitosan Application in Vineyards (Vitis vinifera L. Cv. Tinto Cão) Induces Accumulation of Anthocyanins and Other Phenolics in Berries, Mediated by Modifications in the Transcription of Secondary Metabolism Genes. Int. J. Mol. Sci. 2020, 21, 306. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2-ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Duncan, D.B. A Bayesian Approach to Multiple Comparisons. Technometrics 1965, 7, 171–222. [Google Scholar] [CrossRef]
- Jin, Z.X.; Sun, T.Y.; Sun, H.; Yue, Q.Y.; Yao, Y.X. Modifications of ‘Summer Black’ Grape Berry Quality as Affected by the Different Rootstocks. Sci. Hortic. 2016, 210, 130–137. [Google Scholar] [CrossRef]
- Shahda, M. Effect of Rootstocks on Growth, Yield and Fruit Quality of Red Globe Grape. Ann. Agric. Sci. Moshtohor 2016, 54, 339–344. [Google Scholar] [CrossRef]
- García-Pastor, M.E.; Serrano, M.; Guillén, F.; Castillo, S.; Martínez-Romero, D.; Valero, D.; Zapata, P.J. Methyl Jasmonate Effects on Table Grape Ripening, Vine Yield, Berry Quality and Bioactive Compounds Depend on Applied Concentration. Sci. Hortic. 2019, 247, 380–389. [Google Scholar] [CrossRef]
- Mohamed, A.; El-Salhy, A.; Mostafa, R.; El-Mahdy, M.; Hussein, A. Effect of Exogenous Abscisic Acid (ABA), Gibberellic Acid (GA3) and Cluster Thinning on Yield of Some Grape Cultivars. J. Plant Prod. 2019, 10, 101–105. [Google Scholar] [CrossRef]
- Jia, H.; Zhang, C.; Pervaiz, T.; Zhao, P.; Liu, Z.; Wang, B.; Wang, C.; Zhang, L.; Fang, J.; Qian, J. Jasmonic Acid Involves in Grape Fruit Ripening and Resistant against Botrytis Cinerea. Funct. Integr. Genom. 2016, 16, 79–94. [Google Scholar] [CrossRef] [PubMed]
- Shahab, M.; Roberto, S.R.; Ahmed, S.; Colombo, R.C.; Silvestre, J.P.; Koyama, R.; De Souza, R.T. Anthocyanin Accumulation and Color Development of ‘Benitaka’ Table Grape Subjected to Exogenous Abscisic Acid Application at Different Timings of Ripening. Agronomy 2019, 9, 164. [Google Scholar] [CrossRef]
- Cantín, C.M.; Fidelibus, M.W.; Crisosto, C.H. Application of Abscisic Acid (ABA) at Veraison Advanced Red Color Development and Maintained Postharvest Quality of “Crimson Seedless” Grapes. Postharvest Biol. Technol. 2007, 46, 237–241. [Google Scholar] [CrossRef]
- Ulas, F.; Fricke, A.; Stützel, H. Effect of Grafting on Yield and Quality of Tomato (Solanum Lycopersicum Mill.) Cultivars. Tanzan. J. Agric. Sci. 2018, 17, 39–45. [Google Scholar]
- Rouphael, Y.; Schwarz, D.; Krumbein, A.; Colla, G. Impact of Grafting on Product Quality of Fruit Vegetables. Sci. Hortic. 2010, 127, 172–179. [Google Scholar] [CrossRef]
- Xia, H.; Shen, Y.; Shen, T.; Wang, X.; Zhang, X.; Hu, P.; Liang, D.; Lin, L.; Deng, H.; Wang, J.; et al. Melatonin Accumulation in Sweet Cherry and Its Influence on Fruit Quality and Antioxidant Properties. Molecules 2020, 25, 753. [Google Scholar] [CrossRef]
- El-Anany, A.M.; Hassan, G.F.A.; Ali, F.R. Effects of Edible Coatings on the Shelf-Life and Quality of Anna Apple during Cold Storage. J. Food Technol. 2009, 7, 5–11. [Google Scholar]
- Gong, T.; Zhang, X.; Brecht, J.K.; Serrano, T.; Sims, C.A.; Colee, J.; Zhao, X. Instrumental and Sensory Analyses of Fruit Quality Attributes of Grafted Grape Tomatoes in High Tunnel Organic Production Systems. J. Sci. Food Agric. 2022, 102, 7268–7281. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhong, H.; Zhou, X.; Pan, M.; Xu, J.; Liu, M.; Wang, M.; Liu, G.; Xu, T.; Wang, Y.; et al. Grafting with Rootstocks Promotes Phenolic Compound Accumulation in Grape Berry Skin during Development Based on Integrative Multi-Omics Analysis. Hortic. Res. 2022, 9, uhac055. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.H.; Gemma, H.; Iwahori, S. Stimulation of “Fuji” Apple Skin Color by Ethephon and Phosphorus-Calcium Mixed Compounds in Relation to Flavonoid Synthesis. Sci. Hortic. 2002, 94, 193–199. [Google Scholar] [CrossRef]
- Zhang, P.; Zhou, Z. Postharvest Ethephon Degreening Improves Fruit Color, Flavor Quality and Increases Antioxidant Capacity in ‘Eureka’ Lemon (Citrus Limon (L.) Burm. F.). Sci. Hortic. 2019, 248, 70–80. [Google Scholar] [CrossRef]
- Ribeiro, T.P.; de Lima, M.A.C.; Alves, R.E. Maturação e Qualidade de Uvas Para Suco Em Condições Tropicais, Nos Primeiros Ciclos de Produção. Pesqui. Agropecu. Bras. 2012, 47, 1057–1065. [Google Scholar] [CrossRef]
- Wang, Y.W.; Malladi, A.; Doyle, J.W.; Scherm, H.; Nambeesan, S.U. The Effect of Ethephon, Abscisic Acid, and Methyl Jasmonate on Fruit Ripening in Rabbiteye Blueberry (Vaccinium virgatum). Horticulturae 2018, 4, 24. [Google Scholar] [CrossRef]
- Tecchio, M.A.; Neto, F.J.D.; Junior, A.P.; da Silva, M.J.R.; Roberto, S.R.; Smarsi, R.C. Improvement of Color and Increase in Anthocyanin Content of Niagara Rosada Grapes with Application of Abscisic Acid. Afr. J. Biotechnol. 2017, 16, 1400–1403. [Google Scholar] [CrossRef]
- Gambetta, G.A.; Matthews, M.A.; Shaghasi, T.H.; McElrone, A.J.; Castellarin, S.D. Sugar and Abscisic Acid Signaling Orthologs Are Activated at the Onset of Ripening in Grape. Planta 2010, 232, 219–234. [Google Scholar] [CrossRef]
- Zhang, Z.; Cao, B.; Li, N.; Chen, Z.; Xu, K. Comparative Transcriptome Analysis of the Regulation of ABA Signaling Genes in Different Rootstock Grafted Tomato Seedlings under Drought Stress. Environ. Exp. Bot. 2019, 166, 103814. [Google Scholar] [CrossRef]
- Liu, M.Y.; Song, C.Z.; Chi, M.; Wang, T.M.; Zuo, L.L.; Li, X.L.; Zhang, Z.W.; Xi, Z.M. The Effects of Light and Ethylene and Their Interaction on the Regulation of Proanthocyanidin and Anthocyanin Synthesis in the Skins of Vitis vinifera Berries. Plant Growth Regul. 2016, 79, 377–390. [Google Scholar] [CrossRef]
- Goulas, V.; Manganaris, G.A. Exploring the Phytochemical Content and the Antioxidant Potential of Citrus Fruits Grown in Cyprus. Food Chem. 2012, 131, 39–47. [Google Scholar] [CrossRef]
- Lurie, S.; Ovadia, R.; Nissim-Levi, A.; Oren-Shamir, M.; Kaplunov, T.; Zutahy, Y.; Weksler, H.; Lichter, A. Abscisic Acid Improves Colour Development in “Crimson Seedless” Grapes in the Vineyard and on Detached Berries. J. Hortic. Sci. Biotechnol. 2009, 84, 639–644. [Google Scholar] [CrossRef]
- Dong, T.; Zheng, T.; Fu, W.; Guan, L.; Jia, H.; Fang, J. The Effect of Ethylene on the Color Change and Resistance to Botrytis cinerea Infection in “kyoho” Grape Fruits. Foods 2020, 9, 892. [Google Scholar] [CrossRef] [PubMed]
- An, J.P.; Zhang, X.W.; Bi, S.Q.; You, C.X.; Wang, X.F.; Hao, Y.J. The ERF Transcription Factor MdERF38 Promotes Drought Stress-Induced Anthocyanin Biosynthesis in Apple. Plant J. 2020, 101, 573–589. [Google Scholar] [CrossRef]
- Singh, R.; Rastogi, S.; Dwivedi, U.N. Phenylpropanoid Metabolism in Ripening Fruits. Compr. Rev. Food Sci. Food Saf. 2010, 9, 398–416. [Google Scholar] [CrossRef] [PubMed]
- Sarabandi, M.; Farokhzad, A.; Mandoulakani, B.A.; Ghasemzadeh, R. Biochemical and Gene Expression Responses of Two Iranian Grape Cultivars to Foliar Application of Methyl Jasmonate under Boron Toxicity Conditions. Sci. Hortic. 2019, 249, 355–363. [Google Scholar] [CrossRef]
- Dong, Y.; Liu, L.; Zhao, Z.; Zhi, H.; Guan, J. Effects of 1-MCP on Reactive Oxygen Species, Polyphenol Oxidase Activity, and Cellular Ultra-Structure of Core Tissue in ‘Yali’ Pear (Pyrus Bretschneideri Rehd.) during Storage. Hortic. Environ. Biotechnol. 2015, 56, 207–215. [Google Scholar] [CrossRef]
- Molla, S.M.H.; Rastegar, S.; Omran, V.G.; Khademi, O. Ameliorative Effect of Melatonin against Storage Chilling Injury in Pomegranate Husk and Arils through Promoting the Antioxidant System. Sci. Hortic. 2022, 295, 110889. [Google Scholar] [CrossRef]
- Jeong, S.T.; Goto-Yamamoto, N.; Kobayashi, S.; Esaka, M. Effects of Plant Hormones and Shading on the Accumulation of Anthocyanins and the Expression of Anthocyanin Biosynthetic Genes in Grape Berry Skins. Plant Sci. 2004, 167, 247–252. [Google Scholar] [CrossRef]
- Azuma, A.; Kobayashi, S.; Yakushiji, H.; Yamada, M.; Mitani, N.; Sato, A. VvmybA1 Genotype Determines Grape Skin Color. Vitis—J. Grapevine Res. 2007, 46, 154–155. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Abbreviation | Primer Sequence |
|---|---|---|
| Flavanone3-hydroxylase | F3H | F/5 CAGTGCAAGACTGGCGCGAGATCGTA/3 R/5 TAGCCTCAGACAACACCTCCAGCAACT/3 |
| Chalcone synthase | CHS | F/5 CACTCTTCGAACTCGTCTCT/3 R/5 CCACCAAGCTCTTCTCTATG/3 |
| flavonoid3-o-glucosyltransferase | UFGT | F/5 TGCAGGGCCTAACTCACTCT/3 R/5 GCAGTCGCCTTAGGTAGCAC/3 |
| Beta-Actin | β-actin | F/5 GTGCCTGCCATGTATGTTGCC R/5GCAAGGTCAAGACGAAGGATA |
| Treatment | Average Bunch Weight (g) | Weight 50 Berries (g) | Size 50 Berries (cm3) | |
|---|---|---|---|---|
| Ungrafted | 325.2 ± 4.6 b | 193.9 ± 2.6 a | 162 ± 1.6 a | |
| Grafted | 398.1 ± 5.2 a | 185.9 ± 3.3 b | 160 ± 2.3 a | |
| Cont. | 343.3 ± 5.1 b | 176.8 ± 3.1 b | 150.8 ± 3 c | |
| ABA | 336.8 ± 16.9 b | 191.8 ± 1.7 a | 174.1 ± 4.4 a | |
| MeJ | 388 ± 28.9 a | 193.3 ± 3.7 a | 155 ± 0 bc | |
| Eth | 400 ± 6.7 a | 199.3 ± 6.2 a | 170.8 ± 8.2 ab | |
| Mel | 340.2 ± 7.9 b | 188.3 ± 1.8 ab | 157.5 ± 7.2 bc | |
| Ungrafted | Cont. | 350 ± 10.4 b | 189.6 ± 2.8 cde | 166.6 ± 4.4 ab |
| ABA | 339.1 ± 8.7 b | 202.3 ± 5.2 bc | 178.3 ± 8.8 a | |
| MeJ | 284.7 ± 23.3 c | 192.3 ± 7.8 cd | 150 ± 7.6 bcd | |
| Eth | 331.7 ± 21.3 b | 222.6 ± 6.3 a | 176.6 ± 7.2 a | |
| Mel | 320.5 ± 14.9 bc | 162.6 ± 3.7 f | 141.6 ± 1.4 cd | |
| Grafted | Cont. | 336.6 ± 1.6 b | 164 ± 4 f | 135 ± 2.8 d |
| ABA | 334.4 ± 25.8 b | 181.3 ± 5.3 de | 170 ± 0 ab | |
| MeJ | 491.4 ± 46.9 a | 194.3 ± 0.3 cd | 160 ± 7.6 abc | |
| Eth | 468.3 ± 12.5 a | 176 ± 9.1 ef | 165 ± 10 ab | |
| Mel | 360 ± 16.4 b | 214 ± 2.6 ab | 173.3 ± 13 a | |
| L.S.D0.05% Grafting | 20.3 | 7.2 | 10.3 | |
| L.S.D0.05% Treatment | 32.1 | 11.5 | 16.3 | |
| L.S.D0.05% Interaction | 45.4 | 16.3 | 23 | |
| Treatment | Berries Firmness (N/cm2) | Removal Force (N) | Berries Diameter (mm) | |
|---|---|---|---|---|
| Ungrafted | 667.3 ± 8.8 a | 694.2 ± 15.6 a | 16.5 ± 0.1 a | |
| Grafted | 652.8 ± 31.4 a | 697.1 ± 38.4 a | 16.4 ± 0.1 a | |
| Cont. | 605.5 ± 31.3 b | 645.5 ± 27.4 b | 16.4 ± 0.08 a | |
| ABA | 694.4 ± 14.4 a | 676.6 ± 23.1 b | 16.8 ± 0.3 a | |
| MeJ | 668.8 ± 15.1 a | 697.7 ± 23.1 ab | 16.3 ± 0.5 a | |
| Eth | 672.2 ± 38.8 a | 675 ± 30.7 b | 16.6 ± 0.2 a | |
| Mel | 659.4 ± 26.6 a | 738.3 ± 51.1 a | 16.2 ± 0.2 a | |
| Ungrafted | Cont. | 567.7 ± 24.3 c | 628.8 ± 24.5 b | 16.7 ± 0.3 ab |
| ABA | 728.8 ± 7.7 a | 667.7 ± 20.4 b | 16.7 ± 0.3 ab | |
| MeJ | 681.1 ± 43.7 ab | 686.6 ± 33.3 b | 16.9 ± 0.4 a | |
| Eth | 690 ± 30.2 ab | 691.1 ± 56.1 b | 16.7 ± 0.4 ab | |
| Mel | 668.8 ± 65.4 ab | 796.6 ± 29.1 a | 15.9 ± 0.1 b | |
| Grafted | Cont. | 643.3 ± 48.5 b | 662.2 ± 77.1 b | 16.3 ± 0.2 ab |
| ABA | 660 ± 27.2 ab | 685.5 ± 25.9 b | 17.0 ± 0.6 a | |
| MeJ | 656.6 ± 24.1 b | 708.8 ± 43.3 b | 15.9 ± 0.6 b | |
| Eth | 654.4 ± 47.5 b | 658.8 ± 28 b | 16.5 ± 0.3 ab | |
| Mel | 650 ± 27.1 b | 680 ± 81.9 b | 16.5 ± 0.3 ab | |
| L.S.D0.05% Grafting | 32.2 | 37.6 | 0.3 | |
| L.S.D0.05% Treatment | 50.9 | 59.4 | 0.5 | |
| L.S.D0.05% Interaction | 72.1 | 84.1 | 0.8 | |
| Treatment | Color | |||
|---|---|---|---|---|
| L* | a* | b* | ||
| Ungrafted | 27.4 ± 0.5 b | 9.2 ± 0.4 b | 23.0 ± 0.2 a | |
| Grafted | 30.3 ± 0.6 a | 13.1 ± 0.8 a | 21.1 ± 0.6 b | |
| Cont. | 40 ± 2.2 a | 3.5 ± 0.9 d | 30.4 ± 0.1 a | |
| ABA | 20.2 ± 0.2 d | 12.5 ± 0.7 b | 16.7 ± 0.6 d | |
| MeJ | 23.9 ± 0.7 cd | 14.8 ± 0.7 ab | 21.7 ± 0.9 c | |
| Eth | 27.1 ± 0.7 c | 16 ± 0.6 a | 15.8 ± 1.2 d | |
| Mel | 32.9 ± 1.9 b | 8.9 ± 0.7 c | 25.6 ± 1 b | |
| Ungrafted | Cont. | 42.3 ± 0.4 a | 3.0 ± 0.6 c | 34.3 ± 2.1 a |
| ABA | 25.6 ± 1.3 cd | 8.0 ± 1.5 b | 20.8 ± 1.2 cd | |
| MeJ | 21.3 ± 1.6 de | 13.6 ± 1.1 a | 18.8 ± 0.1 de | |
| Eth | 18.6 ± 0.6 ef | 16.7 ± 1.5 a | 16.0 ± 1.7 ef | |
| Mel | 29.2 ± 2.1 c | 4.8 ± 1.1 bc | 25.2 ± 0.8 b | |
| Grafted | Cont. | 37.7 ± 2.8 ab | 4.1 ± 0.7 bc | 26.5 ± 0.6 b |
| ABA | 14.8 ± 0.8 f | 17 ± 2.1 a | 12.6 ± 1.3 f | |
| MeJ | 26.6 ± 0.9 cd | 16.1 ± 2.2 a | 24.6 ± 2.4 bc | |
| Eth | 35.6 ± 3.1 b | 15.3 ± 1.1 a | 15.6 ± 1.2 ef | |
| Mel | 36.7 ± 0.8 b | 13.1 ± 1.1 a | 26.0 ± 1.3 b | |
| L.S.D0.05% Grafting | 2.4 | 1.9 | 1.9 | |
| L.S.D0.05% Treatment | 3.8 | 3 | 3 | |
| L.S.D0.05% Interaction | 5.3 | 4.3 | 4.3 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salama, A.-M.; Abdelsalam, M.A.; Rehan, M.; Elansary, M.; El-Shereif, A. Anthocyanin Accumulation and Its Corresponding Gene Expression, Total Phenol, Antioxidant Capacity, and Fruit Quality of ‘Crimson Seedless’ Grapevine (Vitis vinifera L.) in Response to Grafting and Pre-Harvest Applications. Horticulturae 2023, 9, 1001. https://doi.org/10.3390/horticulturae9091001
Salama A-M, Abdelsalam MA, Rehan M, Elansary M, El-Shereif A. Anthocyanin Accumulation and Its Corresponding Gene Expression, Total Phenol, Antioxidant Capacity, and Fruit Quality of ‘Crimson Seedless’ Grapevine (Vitis vinifera L.) in Response to Grafting and Pre-Harvest Applications. Horticulturae. 2023; 9(9):1001. https://doi.org/10.3390/horticulturae9091001
Chicago/Turabian StyleSalama, Abdel-Moety, Mohamed A. Abdelsalam, Medhat Rehan, Mohamed Elansary, and Ali El-Shereif. 2023. "Anthocyanin Accumulation and Its Corresponding Gene Expression, Total Phenol, Antioxidant Capacity, and Fruit Quality of ‘Crimson Seedless’ Grapevine (Vitis vinifera L.) in Response to Grafting and Pre-Harvest Applications" Horticulturae 9, no. 9: 1001. https://doi.org/10.3390/horticulturae9091001