

Control of Nematodes in Organic Horticulture Exploiting the Multifunctional Capacity of Microorganisms

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
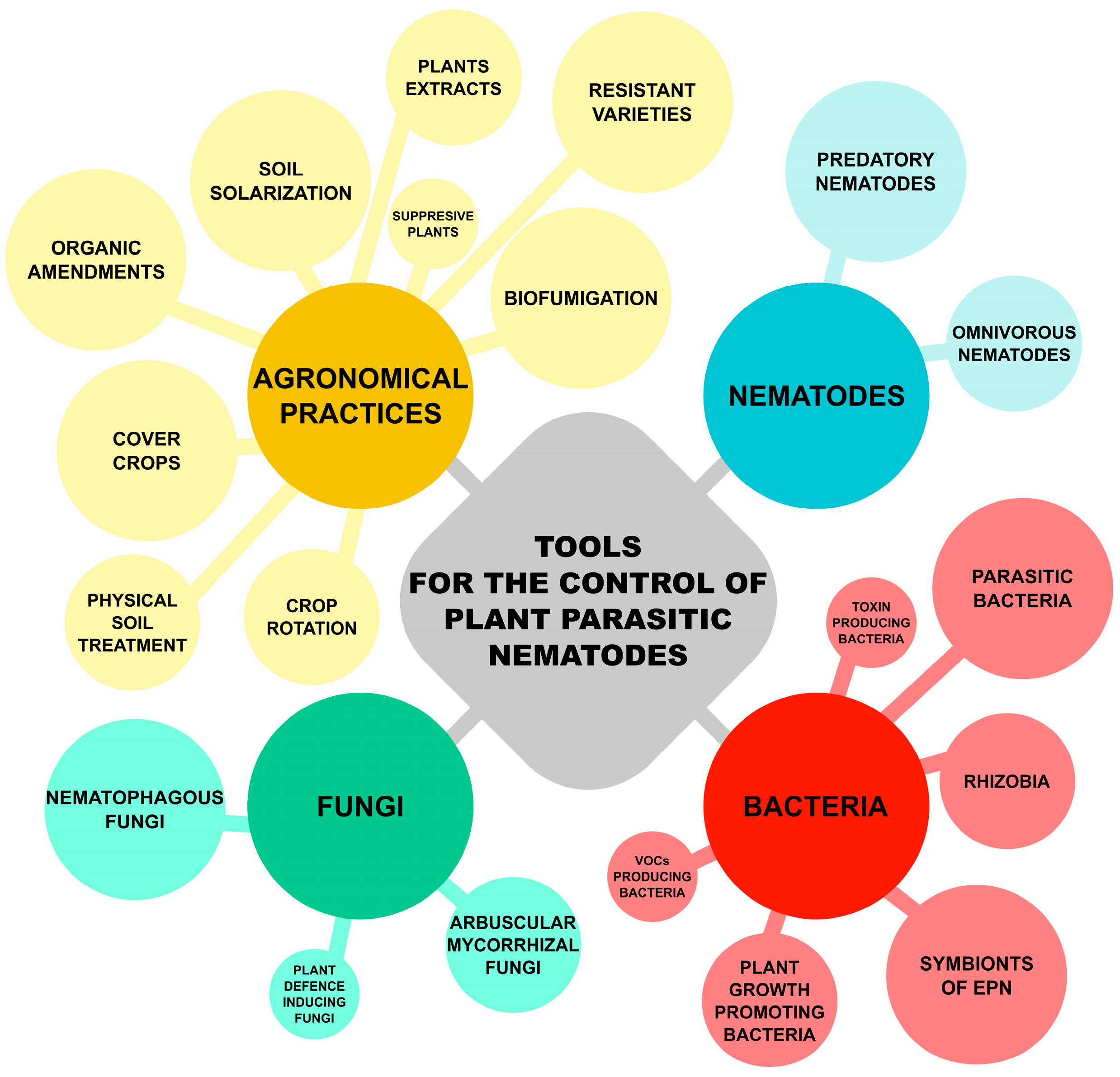
2. Biocontrol of Plant-Parasitic Nematodes
2.1. Bacteria
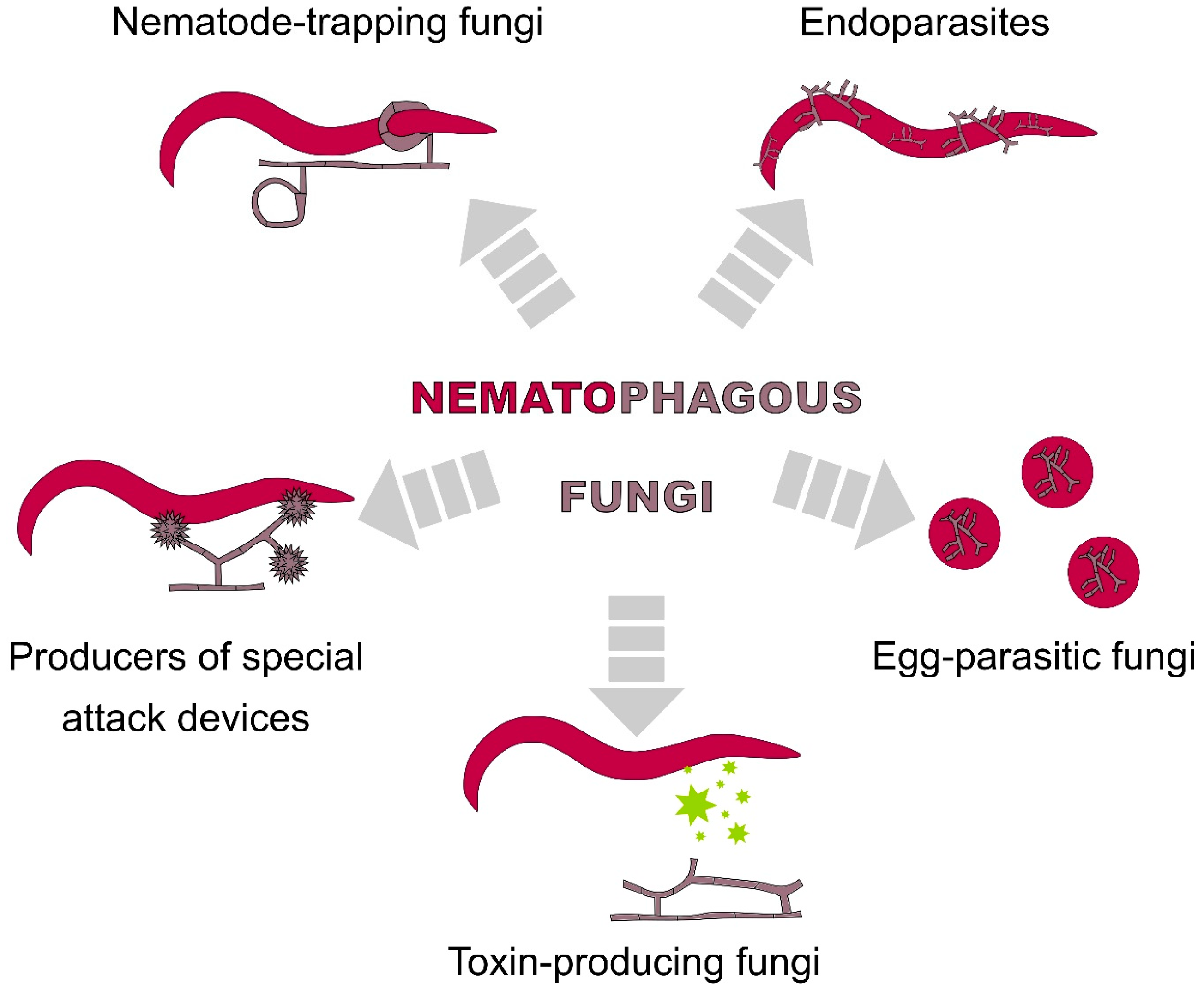
2.2. Fungi
2.2.1. Nematophagous Fungi
2.2.2. Multifunctional Fungi with PPN Control Capacity
2.3. Entomopathogenic Nematodes
2.4. Combination of Bioinocula
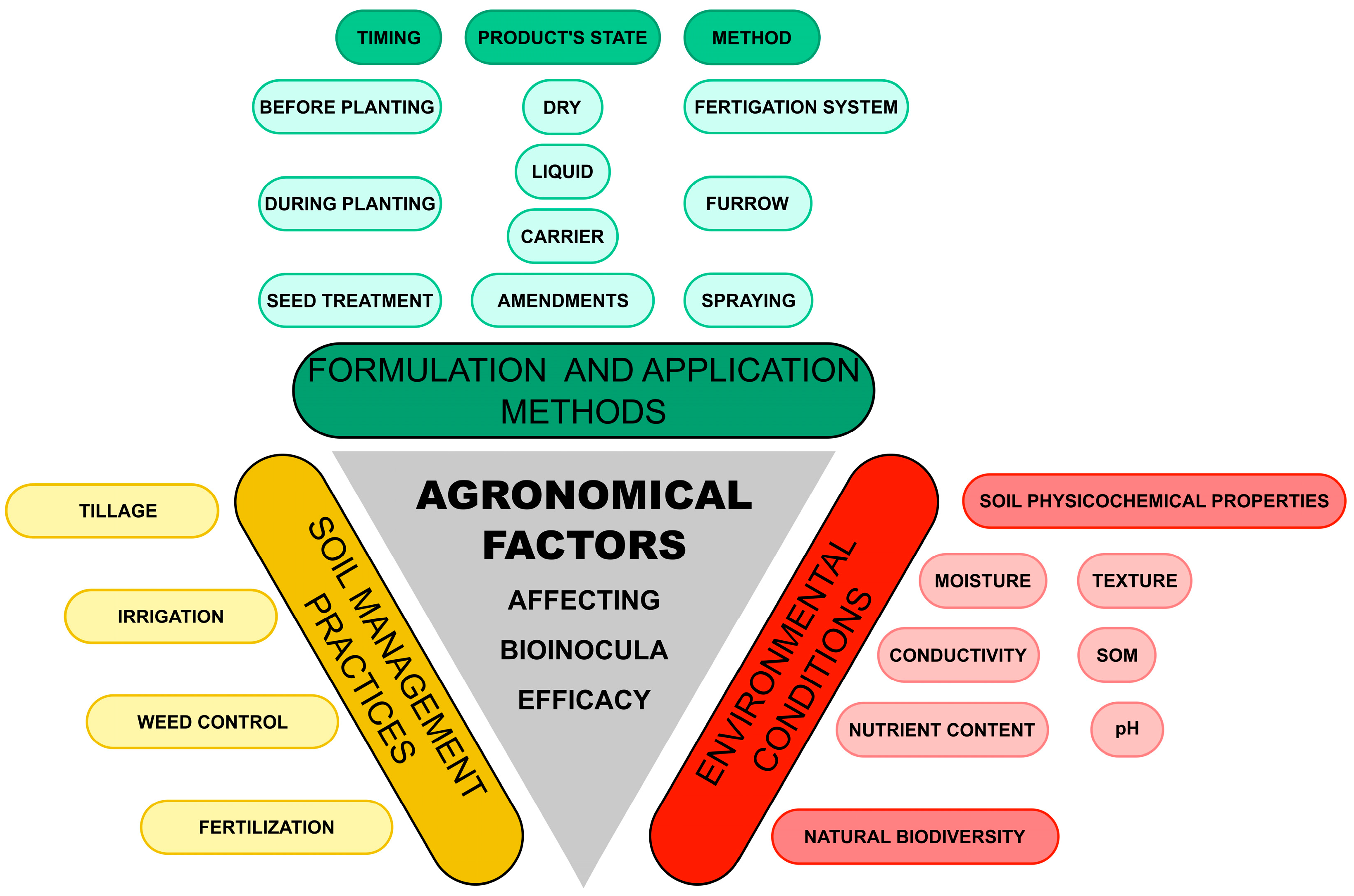
3. Agronomical Factors Affecting the Efficacy of Microbial Inocula for PPN Control
3.1. Environmental Conditions
3.2. Formulation and Application Methods
3.3. Soil Management Practices
4. Conclusions and Future Prospects
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- The European Parliament and the Council of the European Union. Regulation (EU) 2018/848 of the European Par-Liament and of the Council of 30 May 2018 on Organic Production and Lbelling of Organic Products and Repealing Council Regulation (EC) No 834/2007. J. Eur. Union 2018, L150/1. [Google Scholar]
- 2. European Commission. Communication from the Commission to the European Parliament, the Council, the European Eco-nomic and Social Committee and the Committee of the Regions: EU Soil Strategy for 2030—Reaping the Benefits of Healthy Soils for People, Food, Nature and Climate. COM2021699 Final. 2021. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX%3A52021DC0699 (accessed on 3 August 2023).
- Huang, R.; McGrath, S.P.; Hirsch, P.R.; Clark, I.M.; Storkey, J.; Wu, L.; Zhou, J.; Liang, Y. Plant–microbe networks in soil are weakened by century-long use of inorganic fertilizers. Microb. Biotechnol. 2019, 12, 1464–1475. [Google Scholar] [CrossRef] [Green Version]
- de Vries, F.T.; Thébault, E.; Liiri, M.; Birkhofer, K.; Tsiafouli, M.A.; Bjørnlund, L.; Jørgensen, H.B.; Brady, M.V.; Christensen, S.; de Ruiter, P.C.; et al. Soil food web properties explain ecosystem services across European land use systems. Proc. Natl. Acad. Sci. USA 2013, 110, 14296–14301. [Google Scholar] [CrossRef] [PubMed]
- Maron, P.-A.; Sarr, A.; Kaisermann, A.; Lévêque, J.; Mathieu, O.; Guigue, J.; Karimi, B.; Bernard, L.; Dequiedt, S.; Terrat, S.; et al. High Microbial Diversity Promotes Soil Ecosystem Functioning. Appl. Environ. Microbiol. 2018, 84, e02738-17. [Google Scholar] [CrossRef] [Green Version]
- Friberg, H.; Lagerlöf, J.; Rämert, B. Influence of soil fauna on fungal plant pathogens in agricultural and horticultural systems. Biocontrol Sci. Technol. 2005, 15, 641–658. [Google Scholar] [CrossRef]
- Gebremikael, M.T.; Steel, H.; Buchan, D.; Bert, W.; De Neve, S. Nematodes enhance plant growth and nutrient uptake under C and N-rich conditions. Sci. Rep. 2016, 6, 32862. [Google Scholar] [CrossRef] [PubMed]
- Ferris, H.; Bongers, T. Nematode indicators of organic enrichment. J. Nematol. 2006, 38, 3–12. [Google Scholar]
- Malusa, E.; Sas-Paszt, L.; Ciesielska, J. Technologies for Beneficial Microorganisms Inocula Used as Biofertilizers. Sci. World J. 2012, 2012, 491206. [Google Scholar] [CrossRef] [PubMed]
- Bulluck, L.R., III; Barker, K.R.; Ristaino, J.B. Influences of organic and synthetic soil fertility amendments on nematode trophic groups and community dynamics under tomatoes. Appl. Soil Ecol. 2002, 21, 233–250. [Google Scholar] [CrossRef]
- Neher, D.A. Nematode communities in organically and conventionally managed agricultural soils. J. Nematol. 1999, 31, 142–154. [Google Scholar]
- Hallmann, J.; Frankenberg, A.; Paffrath, A.; Schmidt, H. Occurrence and importance of plant-parasitic nematodes in organic farming in Germany. Nematology 2007, 9, 869–879. [Google Scholar] [CrossRef]
- Jones, J.T.; Haegeman, A.; Danchin, E.G.J.; Gaur, H.S.; Helder, J.; Jones, M.G.K.; Kikuchi, T.; Manzanilla-López, R.; Palomares-Rius, J.E.; Wesemael, W.M.L.; et al. Top 10 plant-parasitic nematodes in molecular plant pathology. Mol. Plant Pathol. 2013, 14, 946–961. [Google Scholar] [CrossRef] [Green Version]
- Mia, M.J.; Furmanczyk, E.M.; Golian, J.; Kwiatkowska, J.; Malusá, E.; Neri, D. Living Mulch with Selected Herbs for Soil Management in Organic Apple Orchards. Horticulturae 2021, 7, 59. [Google Scholar] [CrossRef]
- Shilev, S.; Azaizeh, H.; Vassilev, N.; Georgiev, D.; Babrikova, I. Interactions in Soil-Microbe-Plant System: Adaptation to Stressed Agriculture. In Microbial Interventions in Agriculture and Environment: Volume 1: Research Trends, Priorities and Prospects; Singh, D.P., Gupta, V.K., Prabha, R., Eds.; Springer: Singapore, 2019; pp. 131–171. ISBN 9789811383915. [Google Scholar]
- Kowalska, J.; Tyburski, J.; Matysiak, K.; Tylkowski, B.; Malusá, E. Field Exploitation of Multiple Functions of Beneficial Microorganisms for Plant Nutrition and Protection: Real Possibility or Just a Hope? Front. Microbiol. 2020, 11, 1904. [Google Scholar] [CrossRef] [PubMed]
- Vassileva, M.; Flor-Peregrin, E.; Malusá, E.; Vassilev, N. Towards Better Understanding of the Interactions and Efficient Ap-plication of Plant Beneficial Prebiotics, Probiotics, Postbiotics and Synbiotics. Front. Plant Sci. 2020, 11, 1068. [Google Scholar] [CrossRef]
- Siddiqui, Z.A.; Mahmood, I. Biological control of plant parasitic nematodes by fungi: A review. Bioresour. Technol. 1996, 58, 229–239. [Google Scholar] [CrossRef]
- Vassilev, N.; Vassileva, M.; Martos, V.; Del Moral, L.F.G.; Kowalska, J.; Tylkowski, B.; Malusá, E. Formulation of Microbial Inoculants by Encapsulation in Natural Polysaccharides: Focus on Beneficial Properties of Carrier Additives and Derivatives. Front. Plant Sci. 2020, 11, 270. [Google Scholar] [CrossRef]
- Walia, R.K.; Sharma, S.B.; Vats, R. Bacterial Antagonists of Phytonematodes. In Biocontrol Potential and Its Exploitation in Sus-tainable Agriculture: Crop Diseases, Weeds, and Nematodes; Upadhyay, R.K., Mukerji, K.G., Chamola, B.P., Eds.; Springer: Boston, MA, USA, 2000; pp. 173–186. ISBN 978-1-4615-4209-4. [Google Scholar]
- Xiao, T.-J.; Chen, F.; Gao, C.; Zhao, Q.-Y.; Shen, Q.-R.; Ran, W. Bacillus cereus X5 Enhanced Bio-Organic Fertilizers Effectively Control Root-Knot Nematodes (Meloidogyne sp.). Pedosphere 2013, 23, 160–168. [Google Scholar] [CrossRef]
- Terefe, M.; Tefera, T.; Sakhuja, P.K. Effect of a formulation of Bacillus firmus on root-knot nematode Meloidogyne incognita infestation and the growth of tomato plants in the greenhouse and nursery. J. Invertebr. Pathol. 2009, 100, 94–99. [Google Scholar] [CrossRef]
- Zuckerman, B.M.; Dicklow, M.B.; Acosta, N. A Strain of Bacillus thuringiensis for the Control of Plant-parasitic Nematodes. Biocontrol Sci. Technol. 1993, 3, 41–46. [Google Scholar] [CrossRef]
- Du, J.; Gao, Q.; Ji, C.; Song, X.; Liu, Y.; Li, H.; Li, C.; Zhang, P.; Li, J.; Liu, X. Bacillus licheniformis JF-22 to Control Meloidogyne incognita and Its Effect on Tomato Rhizosphere Microbial Community. Front. Microbiol. 2022, 13, 863341. [Google Scholar] [CrossRef]
- Bo, T.; Kong, C.; Zou, S.; Mo, M.; Liu, Y. Bacillus nematocida B16 Enhanced the Rhizosphere Colonization of Pochonia chlamydosporia ZK7 and Controlled the Efficacy of the Root-Knot Nematode Meloidogyne incognita. Microorganisms 2022, 10, 218. [Google Scholar] [CrossRef]
- Siddiqui, I.A.; Shaukat, S.S. Systemic Resistance in Tomato Induced by Biocontrol Bacteria Against the Root-Knot Nematode, Meloidogyne javanicais Independent of Salicylic Acid Production. J. Phytopathol. 2004, 152, 48–54. [Google Scholar] [CrossRef]
- Siddiqui, Z.A.; Iqbal, A.; Mahmood, I. Effects of Pseudomonas fluorescens and fertilizers on the reproduction of Meloidogyne incognita and growth of tomato. Appl. Soil Ecol. 2001, 16, 179–185. [Google Scholar] [CrossRef]
- Seenivasan, N.; Devrajan, K. Management of Meloidogyne Incognita on Medicinal Coleus by Commercial Biocontrol Formu-lations. Nematol. Mediterr. 2008, 36, 61–67. [Google Scholar]
- Zhang, R.; Ouyang, J.; Xu, X.; Li, J.; Rehman, M.; Deng, G.; Shu, J.; Zhao, D.; Chen, S.; Sayyed, R.Z.; et al. Nematicidal Activity of Burkholderia arboris J211 Against Meloidogyne incognita on Tobacco. Front. Microbiol. 2022, 13, 915546. [Google Scholar] [CrossRef]
- Siddiqui, Z.A. Effects of plant growth promoting bacteria and composed organic fertilizers on the reproduction of Meloidogyne incognita and tomato growth. Bioresour. Technol. 2004, 95, 223–227. [Google Scholar] [CrossRef]
- Costa, S.R.; Ng, J.L.P.; Mathesius, U. Interaction of Symbiotic Rhizobia and Parasitic Root-Knot Nematodes in Legume Roots: From Molecular Regulation to Field Application. Mol. Plant Microbe Interact. 2021, 34, 470–490. [Google Scholar] [CrossRef]
- Desaeger, J.; Odee, D.; Machua, J.; Esitubi, M. Interactions between Meloidogyne javanica (Treub) chitwood and rhizobia on growth of Sesbania sesban (L.) Merr. Appl. Soil Ecol. 2005, 29, 252–258. [Google Scholar] [CrossRef]
- Wood, C.W.; Pilkington, B.L.; Vaidya, P.; Biel, C.; Stinchcombe, J.R. Genetic conflict with a parasitic nematode disrupts the legume–rhizobia mutualism. Evol. Lett. 2018, 2, 233–245. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Lei, S.; Cheng, Z.; Jin, L.; Zhang, T.; Liang, L.-M.; Cheng, L.; Zhang, Q.; Xu, X.; Lan, C.; et al. Microbiota and functional analyses of nitrogen-fixing bacteria in root-knot nematode parasitism of plants. Microbiome 2023, 11, 48. [Google Scholar] [CrossRef] [PubMed]
- Rae, R.; Iatsenko, I.; Witte, H.; Sommer, R.J. A subset of naturally isolated Bacillus strains show extreme virulence to the free-living nematodes Caenorhabditis elegans and Pristionchus pacificus. Environ. Microbiol. 2010, 12, 3007–3021. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.X.; Dickson, D.W. Review of Pasteuria penetrans: Biology, Ecology, and Biological Control Potential. J. Nematol. 1998, 30, 313–340. [Google Scholar] [PubMed]
- Malusà, E.; Pinzari, F.; Canfora, L. Efficacy of Biofertilizers: Challenges to Improve Crop Production. In Microbial Inoculants in Sustainable Agricultural Productivity; Singh, D.P., Singh, H.B., Prabha, R., Singh, D.P., Singh, H.B., Prabha, R., Eds.; Springer: New Delhi, India, 2016; pp. 17–40. ISBN 978-81-322-2642-0. [Google Scholar]
- Khan, M.R.; Mohiddin, F.A.; Ahamad, F. Inoculant rhizobia suppressed root-knot disease, and enhanced plant productivity and nutrient uptake of some field-grown food legumes. Acta Agric. Scand. Sect. B—Soil Plant Sci. 2018, 68, 166–174. [Google Scholar] [CrossRef]
- Cheng, W.; Yang, J.; Nie, Q.; Huang, D.; Yu, C.; Zheng, L.; Cai, M.; Thomashow, L.S.; Weller, D.M.; Yu, Z.; et al. Volatile organic compounds from Paenibacillus polymyxa KM2501-1 control Meloidogyne incognita by multiple strategies. Sci. Rep. 2017, 7, 16213. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.; Yang, X.; Xue, H.; Huang, D.; Cai, M.; Huang, F.; Zheng, L.; Yu, Z.; Zhang, J. Reproductive Toxicity of Furfural Acetone in Meloidogyne incognita and Caenorhabditis elegans. Cells 2022, 11, 401. [Google Scholar] [CrossRef]
- Ayaz, M.; Ali, Q.; Farzand, A.; Khan, A.R.; Ling, H.; Gao, X. Nematicidal Volatiles from Bacillus atrophaeus GBSC56 Promote Growth and Stimulate Induced Systemic Resistance in Tomato against Meloidogyne incognita. Int. J. Mol. Sci. 2021, 22, 5049. [Google Scholar] [CrossRef]
- Ye, L.; Wang, J.-Y.; Liu, X.-F.; Guan, Q.; Dou, N.-X.; Li, J.; Zhang, Q.; Gao, Y.-M.; Wang, M.; Zhou, B. Nematicidal activity of volatile organic compounds produced by Bacillus altitudinis AMCC 1040 against Meloidogyne incognita. Arch. Microbiol. 2022, 204, 521. [Google Scholar] [CrossRef]
- Diyapoglu, A.; Chang, T.-H.; Chang, P.-F.L.; Yen, J.-H.; Chiang, H.-I.; Meng, M. Fumigant Activity of Bacterial Volatile Organic Compounds against the Nematodes Caenorhabditis elegans and Meloidogyne incognita. Molecules 2022, 27, 4714. [Google Scholar] [CrossRef]
- Pulavarty, A.; Singh, A.; Smyth, D.; Mehta, J.P.; Horgan, K.; Kakouli-Duarte, T. Sustainable management of the potato cyst nematode, Globodera rostochiensis, with two microbial fermentation products. Front. Plant Sci. 2022, 13, 987059. [Google Scholar] [CrossRef]
- Li, J.; Zou, C.; Xu, J.; Ji, X.; Niu, X.; Yang, J.; Huang, X.; Zhang, K.-Q. Molecular Mechanisms of Nematode-Nematophagous Microbe Interactions: Basis for Biological Control of Plant-Parasitic Nematodes. Annu. Rev. Phytopathol. 2015, 53, 67–95. [Google Scholar] [CrossRef] [PubMed]
- Su, H.; Zhao, Y.; Zhou, J.; Feng, H.; Jiang, D.; Zhang, K.-Q.; Yang, J. Trapping devices of nematode-trapping fungi: Formation, evolution, and genomic perspectives. Biol. Rev. 2017, 92, 357–368. [Google Scholar] [CrossRef] [PubMed]
- Nordbring-Hertz, B.; Jansson, H.-B.; Tunlid, A. Nematophagous Fungi. In Encyclopedia of Life Sciences; Wiley-Blackwell: Hoboken, NJ, USA, 2006; ISBN 978-0-470-01590-2. [Google Scholar]
- Lopez-Llorca, L.V.; Maciá-Vicente, J.G.; Jansson, H.-B. Mode of Action and Interactions of Nematophagous Fungi. In Integrated Management and Biocontrol of Vegetable and Grain Crops Nematodes; Ciancio, A., Mukerji, K.G., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 51–76. ISBN 978-1-4020-6063-2. [Google Scholar]
- Jaffee, B.A. Correlations Between Most Probable Number and Activity of Nematode-Trapping Fungi. Phytopathology 2003, 93, 1599–1605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaffee, B.A. Augmentation of Soil with the Nematophagous Fungi Hirsutella rhossiliensis and Arthrobotrys haptotyla. Phytopathology 2000, 90, 498–504. [Google Scholar] [CrossRef] [Green Version]
- Jaffee, B.A. Do Organic Amendments Enhance the Nematode-Trapping Fungi Dactylellina Haptotyla and Arthrobotrys Oli-gospora? J. Nematol. 2004, 36, 267–275. [Google Scholar]
- Jaffee, B.A.; Ferris, H.; Scow, K.M. Nematode-Trapping Fungi in Organic and Conventional Cropping Systems. Phytopathology 1998, 88, 344–350. [Google Scholar] [CrossRef] [Green Version]
- Giri, B.; Rawat, R.; Saxena, G.; Manchanda, P.; Wu, Q.-S.; Sharma, A. Effect of Rhizoglomus fasciculatum and Paecilomyces lilacinus in the biocontrol of root-knot nematode, Meloidogyne incognita in Capsicum annuum L. Commun. Integr. Biol. 2022, 15, 75–87. [Google Scholar] [CrossRef]
- EU Pesticide Database. Available online: https://ec.europa.eu/food/plant/pesticides/eu-pesticides-database/start/screen/active-substances (accessed on 14 April 2023).
- Khan, M.; Tanaka, K. Purpureocillium lilacinum for plant growth promotion and biocontrol against root-knot nematodes infecting eggplant. PLoS ONE 2023, 18, e0283550. [Google Scholar] [CrossRef]
- Lan, X.; Zhang, J.; Zong, Z.; Ma, Q.; Wang, Y. Evaluation of the Biocontrol Potential of Purpureocillium lilacinum QLP12 against Verticillium dahliae in Eggplant. BioMed Res. Int. 2017, 2017, 4101357. [Google Scholar] [CrossRef] [Green Version]
- Manzanilla-López, R.H.; Esteves, I.; Finetti-Sialer, M.M.; Hirsch, P.R.; Ward, E.; Devonshire, J.; Hidalgo-Díaz, L. Pochonia Chlamydosporia: Advances and Challenges to Improve Its Performance as a Biological Control Agent of Sedentary En-do-Parasitic Nematodes. J. Nematol. 2013, 45, 1–7. [Google Scholar]
- Yang, J.; Liang, L.; Li, J.; Zhang, K.-Q. Nematicidal enzymes from microorganisms and their applications. Appl. Microbiol. Biotechnol. 2013, 97, 7081–7095. [Google Scholar] [CrossRef]
- Lopez-Llorca, L.V.; Olivares-Bernabeu, C.; Salinas, J.; Jansson, H.-B.; Kolattukudy, P.E. Pre-penetration events in fungal parasitism of nematode eggs. Mycol. Res. 2002, 106, 499–506. [Google Scholar] [CrossRef]
- Bontempo, A.F.; Lopes, E.A.; Fernandes, R.H.; DE Freitas, L.G.; Dallemole-Giaretta, R. DOSE-RESPONSE EFFECT OF Pochonia chlamydosporia AGAINST Meloidogyne incognita ON CARROT UNDER FIELD CONDITIONS. Rev. Caatinga 2017, 30, 258–262. [Google Scholar] [CrossRef] [Green Version]
- Viggiano, J.R.; de Freitas, L.G.; Lopes, E.A. Use of Pochonia chlamydosporia to control Meloidogyne javanica in cucumber. Biol. Control 2014, 69, 72–77. [Google Scholar] [CrossRef]
- Tylkowski, B.; Olkiewicz, M.; Montane, X.; Nogalska, A.; Haponska, M.; Montornes, J.M.; Kowalska, J.; Malusá, E. Encapsulation Technologies in Agriculture; De Gruyter: Berlin, Germany, 2020; pp. 287–302. ISBN 978-3-11-064207-0. [Google Scholar]
- Escudero, N.; Ferreira, S.R.; Lopez-Moya, F.; Naranjo-Ortiz, M.A.; Marin-Ortiz, A.I.; Thornton, C.R.; Lopez-Llorca, L.V. Chitosan enhances parasitism of Meloidogyne javanica eggs by the nematophagous fungus Pochonia chlamydosporia. Fungal Biol. 2016, 120, 572–585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escudero, N.; Lopez-Moya, F.; Ghahremani, Z.; Zavala-Gonzalez, E.A.; Alaguero-Cordovilla, A.; Ros-Ibañez, C.; Lacasa, A.; Sorribas, F.J.; Lopez-Llorca, L.V. Chitosan Increases Tomato Root Colonization by Pochonia chlamydosporia and Their Combination Reduces Root-Knot Nematode Damage. Front. Plant Sci. 2017, 8, 1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iqbal, M.; Broberg, M.; Haarith, D.; Broberg, A.; Bushley, K.E.; Durling, M.B.; Viketoft, M.; Jensen, D.F.; Dubey, M.; Karlsson, M. Natural variation of root lesion nematode antagonism in the biocontrol fungus Clonostachys rosea and identification of biocontrol factors through genome-wide association mapping. Evol. Appl. 2020, 13, 2264–2283. [Google Scholar] [CrossRef]
- Sahebani, N.; Hadavi, N. Biological control of the root-knot nematode Meloidogyne javanica by Trichoderma harzianum. Soil Biol. Biochem. 2008, 40, 2016–2020. [Google Scholar] [CrossRef]
- Sun, M.-H.; Gao, L.; Shi, Y.-X.; Li, B.-J.; Liu, X.-Z. Fungi and actinomycetes associated with Meloidogyne spp. eggs and females in China and their biocontrol potential. J. Invertebr. Pathol. 2006, 93, 22–28. [Google Scholar] [CrossRef]
- Zhang, J.; Fu, B.; Lin, Q.; Riley, I.T.; Ding, S.; Chen, L.; Cui, J.; Yang, L.; Li, H. Colonization of Beauveria bassiana 08F04 in root-zone soil and its biocontrol of cereal cyst nematode (Heterodera filipjevi). PLoS ONE 2020, 15, e0232770. [Google Scholar] [CrossRef]
- Wonganu, B.; Pootanakit, K.; Boonyapakron, K.; Champreda, V.; Tanapongpipat, S.; Eurwilaichitr, L. Cloning, expression and characterization of a thermotolerant endoglucanase from Syncephalastrum racemosum (BCC18080) in Pichia pastoris. Protein Expr. Purif. 2008, 58, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Hashem, M.; Abo-Elyousr, K.A. Management of the root-knot nematode Meloidogyne incognita on tomato with combinations of different biocontrol organisms. Crop Prot. 2011, 30, 285–292. [Google Scholar] [CrossRef]
- Huang, W.-K.; Sun, J.-H.; Cui, J.-K.; Wang, G.-F.; Kong, L.-A.; Peng, H.; Chen, S.-L.; Peng, D.-L. Efficacy Evaluation of Fungus Syncephalastrum racemosum and Nematicide Avermectin against the Root-Knot Nematode Meloidogyne incognita on Cucumber. PLoS ONE 2014, 9, e89717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.-K.; Cui, J.-K.; Liu, S.-M.; Kong, L.-A.; Wu, Q.-S.; Peng, H.; He, W.-T.; Sun, J.-H.; Peng, D.-L. Testing various biocontrol agents against the root-knot nematode (Meloidogyne incognita) in cucumber plants identifies a combination of Syncephalastrum racemosum and Paecilomyces lilacinus as being most effective. Biol. Control 2016, 92, 31–37. [Google Scholar] [CrossRef]
- Affokpon, A.; Coyne, D.L.; Lawouin, L.; Tossou, C.; Agbèdè, R.D.; Coosemans, J. Effectiveness of native West African arbuscular mycorrhizal fungi in protecting vegetable crops against root-knot nematodes. Biol. Fertil. Soils 2011, 47, 207–217. [Google Scholar] [CrossRef]
- Castillo, P.; Nico, A.I.; Azcón-Aguilar, C.; Rincón, C.D.R.; Calvet, C.; Jiménez-Díaz, R.M. Protection of olive planting stocks against parasitism of root-knot nematodes by arbuscular mycorrhizal fungi. Plant Pathol. 2006, 55, 705–713. [Google Scholar] [CrossRef] [Green Version]
- Marro, N.; Lax, P.; Cabello, M.; Doucet, M.E.; Becerra, A.G. Use of the arbuscular mycorrhizal fungus Glomus intraradices as biological control agent of the nematode Nacobbus aberrans parasitizing tomato. Braz. Arch. Biol. Technol. 2014, 57, 668–674. [Google Scholar] [CrossRef] [Green Version]
- Schouteden, N.; De Waele, D.; Panis, B.; Vos, C.M. Arbuscular Mycorrhizal Fungi for the Biocontrol of Plant-Parasitic Nematodes: A Review of the Mechanisms Involved. Front. Microbiol. 2015, 6, 1280. [Google Scholar] [CrossRef] [Green Version]
- de Sá, C.S.B.; Campos, M.A.S. Arbuscular mycorrhizal fungi decrease Meloidogyne enterolobii infection of Guava seedlings. J. Helminthol. 2020, 94, e183. [Google Scholar] [CrossRef]
- Hao, Z.; Fayolle, L.; van Tuinen, D.; Chatagnier, O.; Li, X.; Gianinazzi, S.; Gianinazzi-Pearson, V. Local and systemic mycorrhiza-induced protection against the ectoparasitic nematode Xiphinema index involves priming of defence gene responses in grapevine. J. Exp. Bot. 2012, 63, 3657–3672. [Google Scholar] [CrossRef] [Green Version]
- Vos, C.; Tesfahun, A.; Panis, B.; De Waele, D.; Elsen, A. Arbuscular mycorrhizal fungi induce systemic resistance in tomato against the sedentary nematode Meloidogyne incognita and the migratory nematode Pratylenchus penetrans. Appl. Soil Ecol. 2012, 61, 1–6. [Google Scholar] [CrossRef]
- Hao, Z.; van Tuinen, D.; Fayolle, L.; Chatagnier, O.; Li, X.; Chen, B.; Gianinazzi, S.; Gianinazzi-Pearson, V. Arbuscular mycorrhiza affects grapevine fanleaf virus transmission by the nematode vector Xiphinema index. Appl. Soil Ecol. 2018, 129, 107–111. [Google Scholar] [CrossRef]
- Garita, S.A.; Bernardo, V.F.; Guimarães, M.D.A.; Arango, M.C.; Ruscitti, M.F. Mycorrhization and grafting improve growth in the tomato and reduce the population of Nacobbus aberrans. Rev. Ciênc. Agron. 2019, 50, 609–615. [Google Scholar] [CrossRef]
- Pawlowski, M.L.; Hartman, G.L. Impact of Arbuscular Mycorrhizal Species on Heterodera glycines. Plant Dis. 2020, 104, 2406–2410. [Google Scholar] [CrossRef]
- Ferreira, B.S.; Santana, M.V.; Macedo, R.S.; Silva, J.O.; Carneiro, M.A.C.; Rocha, M.R. Co-occurrence patterns between plant-parasitic nematodes and arbuscular mycorrhizal fungi are driven by environmental factors. Agric. Ecosyst. Environ. 2018, 265, 54–61. [Google Scholar] [CrossRef]
- Kepenekci, I.; Hazir, S.; Lewis, E.E. Evaluation of entomopathogenic nematodes and the supernatants of the in vitro culture medium of their mutualistic bacteria for the control of the root-knot nematodes Meloidogyne incognita and M. arenaria. Pest Manag. Sci. 2016, 72, 327–334. [Google Scholar] [CrossRef] [PubMed]
- El Aimani, A.; Houari, A.; Laasli, S.-E.; Mentag, R.; Iraqi, D.; Diria, G.; Khayi, S.; Lahlali, R.; Dababat, A.A.; Mokrini, F. Antagonistic potential of Moroccan entomopathogenic nematodes against root-knot nematodes, Meloidogyne javanica on tomato under greenhouse conditions. Sci. Rep. 2022, 12, 2915. [Google Scholar] [CrossRef]
- Lacey, L.A.; Georgis, R. Entomopathogenic Nematodes for Control of Insect Pests Above and Below Ground with Comments on Commercial Production. J. Nematol. 2012, 44, 218–225. [Google Scholar]
- Del Valle, E.E.; Lax, P.; Dueñas, J.R.; Doucet, M.E. Effects of insect cadavers infected by Heterorhabditis bacteriophora and Steinernema diaprepesi on Meloidogyne incognita parasitism in pepper and summer squash plants. Cienc. Investig. Agrar. 2013, 40, 109–118. [Google Scholar] [CrossRef] [Green Version]
- Caccia, M.; Marro, N.; Dueñas, J.R.; Doucet, M.E.; Lax, P. Effect of the entomopathogenic nematode-bacterial symbiont complex on Meloidogyne hapla and Nacobbus aberrans in short-term greenhouse trials. Crop Prot. 2018, 114, 162–166. [Google Scholar] [CrossRef] [Green Version]
- Sharma, M.P.; Sharma, A.N.; Hussaini, S.S. Entomopathogenic nematodes, a potential microbial biopesticide: Mass production and commercialisation status—A mini review. Arch. Phytopathol. Plant Prot. 2011, 44, 855–870. [Google Scholar] [CrossRef]
- Vyas, R.V.; Patel, B.; Maghodia, A.; Patel, D.J. Significance of Metabolites of Native Xenorhabdus, a Bacterial Symbiont of Steinernema, for Suppression of Collar Rot and Root Knot Diseases of Groundnut. Indian J. Biotechnol. 2008, 7, 371–377. [Google Scholar]
- Vagelas, I.K.; Pembroke, B.; Gowen, S.R.; Davies, K.G. The control of root-knot nematodes (Meloidogyne spp.) by Pseudomonas oryzihabitans and its immunological detection on tomato roots. Nematology 2007, 9, 363–370. [Google Scholar] [CrossRef]
- Orozco, R.A.; Molnár, I.; Bode, H.; Stock, S.P. Bioprospecting for secondary metabolites in the entomopathogenic bacterium Photorhabdus luminescens subsp. sonorensis. J. Invertebr. Pathol. 2016, 141, 45–52. [Google Scholar] [CrossRef]
- Kusakabe, A.; Wang, C.; Xu, Y.-M.; Molnár, I.; Stock, S.P. Selective Toxicity of Secondary Metabolites from the Entomopathogenic Bacterium Photorhabdus luminescens sonorensis against Selected Plant Parasitic Nematodes of the Tylenchina Suborder. Microbiol. Spectr. 2022, 10, e0257721. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.L.F.; Roberts, D.P. Combinations of biocontrol agents for management of plant-parasitic nematodes and soilborne plant-pathogenic fungi. J. Nematol. 2002, 34, 1–8. [Google Scholar]
- Siddiqui, Z.A.; Baghel, G.; Akhtar, M.S. Biocontrol of Meloidogyne javanica by Rhizobium and plant growth-promoting rhizobacteria on lentil. World J. Microbiol. Biotechnol. 2007, 23, 435–441. [Google Scholar] [CrossRef]
- Díaz-Manzano, F.E.; Amora, D.X.; Martínez-Gómez, Á.; Moelbak, L.; Escobar, C. Biocontrol of Meloidogyne spp. in Solanum lycopersicum using a dual combination of Bacillus strains. Front. Plant Sci. 2023, 13, 1077062. [Google Scholar] [CrossRef]
- Sharma, I.P.; Sharma, A.K. Physiological and biochemical changes in tomato cultivar PT-3 with dual inoculation of mycorrhiza and PGPR against root-knot nematode. Symbiosis 2017, 71, 175–183. [Google Scholar] [CrossRef]
- Van der Veken, L.; Cabasan, M.T.N.; Elsen, A.; Swennen, R.; De Waele, D. Effect of single or dual inoculation of the arbuscular mycorrhizal fungus Glomus mosseae and root-nodulating rhizobacteria on reproduction of the burrowing nematode Radopholus similis on non-leguminous and leguminous banana intercrops. J. Plant Dis. Prot. 2021, 128, 961–971. [Google Scholar] [CrossRef]
- Flor-Peregrín, E.; Azcón, R.; Martos, V.; Verdejo-Lucas, S.; Talavera, M. Effects of dual inoculation of mycorrhiza and endophytic, rhizospheric or parasitic bacteria on the root-knot nematode disease of tomato. Biocontrol Sci. Technol. 2014, 24, 1122–1136. [Google Scholar] [CrossRef]
- Nafady, N.A.; Sultan, R.; El-Zawahry, A.M.; Mostafa, Y.S.; Alamri, S.; Mostafa, R.G.; Hashem, M.; Hassan, E.A. Effective and Promising Strategy in Management of Tomato Root-Knot Nematodes by Trichoderma harzianum and Arbuscular Mycorrhizae. Agronomy 2022, 12, 315. [Google Scholar] [CrossRef]
- Giovannini, L.; Palla, M.; Agnolucci, M.; Avio, L.; Sbrana, C.; Turrini, A.; Giovannetti, M. Arbuscular Mycorrhizal Fungi and Associated Microbiota as Plant Biostimulants: Research Strategies for the Selection of the Best Performing Inocula. Agronomy 2020, 10, 106. [Google Scholar] [CrossRef] [Green Version]
- Pandey, S. Can VAM Occurring in the Rhizosphere of Cowpea Be A Source of Natural Antagonist to Heterodera Cajani Pop-ulation? Indian J. Fundam. Appl. Life Sci. 2011, 1, 51–58. [Google Scholar]
- Banuelos, J.; Alarcón, A.; Larsen, J.; Cruz-Sánchez, S.; Trejo, D. Interactions between arbuscular mycorrhizal fungi and Meloidogyne incognitain the ornamental plant Impatiens balsamina. J. Soil Sci. Plant Nutr. 2014, 14, 63–74. [Google Scholar] [CrossRef] [Green Version]
- Lax, P.; Becerra, A.G.; Soteras, F.; Cabello, M.; Doucet, M.E. Effect of the arbuscular mycorrhizal fungus Glomus intraradices on the false root-knot nematode Nacobbus aberrans in tomato plants. Biol. Fertil. Soils 2011, 47, 591–597. [Google Scholar] [CrossRef] [Green Version]
- Marro, N.; Caccia, M.; Doucet, M.E.; Cabello, M.; Becerra, A.; Lax, P. Mycorrhizas reduce tomato root penetration by false root-knot nematode Nacobbus aberrans. Appl. Soil Ecol. 2018, 124, 262–265. [Google Scholar] [CrossRef]
- Gautam, A.; Siddiqui, A.; Mahmood, I. Integrated Management of Meloidogyne Incognita on Tomato. Nematol. Mediterr. 1995, 23, 245–247. [Google Scholar]
- Tiwari, S.; Pandey, S.; Chauhan, P.S.; Pandey, R. Biocontrol agents in co-inoculation manages root knot nematode [ Meloidogyne incognita (Kofoid & White) Chitwood] and enhances essential oil content in Ocimum basilicum L. Ind. Crops Prod. 2017, 97, 292–301. [Google Scholar] [CrossRef]
- dos Santos, M.C.V.; Esteves, I.; Kerry, B.; Abrantes, I. Biology, growth parameters and enzymatic activity of Pochonia chlamydosporia isolated from potato cyst and root-knot nematodes. Nematology 2013, 15, 493–504. [Google Scholar] [CrossRef]
- Kerry, B.R.; Irving, F.; Hornsey, J.C. Variation Between Strains of the Nematophagous Fungus, Verticillium Chlamydosporium Goddard. I. Factors Affecting Growth in Vitro. Nematologica 1986, 32, 461–473. [Google Scholar] [CrossRef]
- Leij, F.A.A.M.D.; Kerry, B.R.; Dennehy, J.A. Verticillium Chlamydosporium as a Biological Control Agent for Meloidogyne Incognita and M. Hapla in Pot and Micro-Plot Tests. Nematologica 1993, 39, 115–126. [Google Scholar] [CrossRef]
- Nasu, d.G.C.; Amora, D.X.; Monteiro, T.S.A.; Alves, P.S.; de Podestá, G.S.; Ferreira, F.C.; de Freitas, L.G. Pochonia chlamydosporia applied via seed treatment for nematode control in two soil types. Crop Prot. 2018, 114, 106–112. [Google Scholar] [CrossRef]
- Gray, N.F. Ecology of nematophagous fungi: Effect of soil moisture, organic matter, pH and nematode density on distribution. Soil Biol. Biochem. 1985, 17, 499–507. [Google Scholar] [CrossRef]
- Jamiołkowska, A.; Księżniak, A.; Gałązka, A.; Hetman, B.; Kopacki, M.; Skwaryło-Bednarz, B. Impact of abiotic factors on development of the community of arbuscular mycorrhizal fungi in the soil: A Review. Int. Agrophysics 2018, 32, 133–140. [Google Scholar] [CrossRef]
- Pathak, E.; Campos–Herrera, R.; El–Borai, F.E.; Duncan, L.W. Spatial relationships between entomopathogenic nematodes and nematophagous fungi in Florida citrus orchards. J. Invertebr. Pathol. 2017, 144, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Gray, N.F. Ecology of nematophagous fungi: Effect of the soil nutrients N, P and K, and seven major metals on distribution. Plant Soil 1988, 108, 286–290. [Google Scholar] [CrossRef]
- Eayre, C.G.; Jaffe, B.A.; Zehr, E.I. Suppression of Criconemella Xenoplax by the Fungus Hirsutella Rhossiliensis. Phytopathology 1983, 73, 500. [Google Scholar] [CrossRef]
- Sun, J.; Wang, H.; Lu, F.; Du, L.; Wang, G. The efficacy of nematicidal strain Syncephalastrum racemosum. Ann. Microbiol. 2008, 58, 369–373. [Google Scholar] [CrossRef]
- Podestá, G.S.; Amora, D.X.; Maffia, L.A.; Nasu, G.C.; Ferraz, S.; Freitas, L.G. Effect of time between soil infestation with Pochonia chlamidosporia and planting on the efficacy of the fungus in managing Meloidogyne javanica. Crop Prot. 2016, 90, 77–83. [Google Scholar] [CrossRef]
- Bontempo, A.F.; Fernandes, R.H.; Lopes, J.; Freitas, L.G.; Lopes, E.A. Pochonia Chlamydosporia Controls Meloidogyne Incog-nita on Carrot. Australas. Plant Pathol. 2014, 43, 421–424. [Google Scholar] [CrossRef]
- Malusá, E.; Tartanus, M.; Soika, G. Monitoring and possibilities of controlling nematodes and fruit damaging pests of Rosa spp. with microbial-derived products. J. Plant Prot. Res. 2019, 59, 334–340. [Google Scholar] [CrossRef]
- Anastasiadis, I.A.; Giannakou, I.O.; Prophetou-Athanasiadou, D.A.; Gowen, S.R. The combined effect of the application of a biocontrol agent Paecilomyces lilacinus, with various practices for the control of root-knot nematodes. Crop Prot. 2008, 27, 352–361. [Google Scholar] [CrossRef]
- Dababat, A.; Sikora, R.A. Importance of Application Time and Inoculum Density of Fusarium Ox-Ysporum 162 for Biological Control of Meloidogyne Incognita on Tomato. Nematropica 2007, 91, 267–276. [Google Scholar]
- O’callaghan, M. Microbial inoculation of seed for improved crop performance: Issues and opportunities. Appl. Microbiol. Biotechnol. 2016, 100, 5729–5746. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; van der Putten, W.H. Going back to the roots: The microbial ecology of the rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef]
- Jaffee, B.A.; Muldoon, A.E. Susceptibility of root-knot and cyst nematodes to the nematode-trapping fungi Monacrosporium ellipsosporum and M. cionopagum. Soil Biol. Biochem. 1995, 27, 1083–1090. [Google Scholar] [CrossRef]
- Aboul-Eid, H.Z.; Noweer, E.M.A.; Ashour, N.E.; Ameen, H.H. Evaluation of a nematode bio-product Dbx-20% against root-knot nematode Meloidogyne incognita affecting grapevine under field conditions. Commun. Agric. Appl. Biol. Sci. 2006, 71, 659–668. [Google Scholar]
- Song, Z.Y.; Shen, L.; Zhong, Q.; Yin, Y.P.; Wang, Z.K. Liquid culture production of microsclerotia of Purpureocillium lilacinum for use as bionematicide. Nematology 2016, 18, 719–726. [Google Scholar] [CrossRef]
- Nagachandrabose, S. Liquid bioformulations for the management of root-knot nematode, Meloidogyne hapla that infects carrot. Crop Prot. 2018, 114, 155–161. [Google Scholar] [CrossRef]
- Rao, M.S.; Kamalnath, M.; Umamaheswari, R.; Rajinikanth, R.; Prabu, P.; Priti, K.; Grace, G.N.; Chaya, M.K.; Gopalakrishnan, C. Bacillus subtilis IIHR BS-2 enriched vermicompost controls root knot nematode and soft rot disease complex in carrot. Sci. Hortic. 2017, 218, 56–62. [Google Scholar] [CrossRef]
- Sowmya, D.S.; Rao, M.S.; Kumar, R.M.; Gavaskar, J.; Priti, K. Bio-Management of Meloidogyne Incognita and Erwinia Ca-rotovora in Carrot (Daucus Carota L.) Using Pseudomonas Putida and Paecilomyces Lilacinus. Nematol. Mediterr. 2012, 40, 189–194. [Google Scholar]
- Guru Prasad, G.R.; Ravichandra, N.G.; Narasimhamurthy, T.N.; Punith Kumar, C.H.; Yadahalli, P. Management of Meloidogyne Inognita Infecting Carrot by Using Bioagents. JBiopest 2014, 7, 144–150. [Google Scholar]
- Giannakou, I.O.; Karpouzas, D.G.; Prophetou-Athanasiadou, D. A novel non-chemical nematicide for the control of root-knot nematodes. Appl. Soil Ecol. 2004, 26, 69–79. [Google Scholar] [CrossRef]
- Stirling, G.R.; Smith, L.J.; Licastro, K.A.; Eden, L.M. Control of Root-knot Nematode with Formulations of the Nematode-Trapping FungusArthrobotrys dactyloides. Biol. Control. 1998, 11, 224–230. [Google Scholar] [CrossRef]
- Siddiqui, Z.A.; Futai, K. Biocontrol of Meloidogyne incognita on tomato using antagonistic fungi, plant-growth-promoting rhizobacteria and cattle manure. Pest Manag. Sci. 2009, 65, 943–948. [Google Scholar] [CrossRef] [PubMed]
- Saumell, C.A.; Padilha, T.; Santos, C.d.P. Nematophagous fungi in sheep faeces in Minas Gerais, Brazil. Mycol. Res. 2000, 104, 1005–1008. [Google Scholar] [CrossRef]
- Abawi, G.S.; Widmer, T.L. Impact of soil health management practices on soilborne pathogens, nematodes and root diseases of vegetable crops. Appl. Soil Ecol. 2000, 15, 37–47. [Google Scholar] [CrossRef]
- Wang, L.; Wang, Y.; Xiu, W.; Tan, B.; Li, G.; Zhao, J.; Yang, D.; Zhang, G.; Zhang, Y. Responses of Soil Microbial and Nematode Communities to Various Cover Crop Patterns in a Tea Garden of China. Int. J. Environ. Res. Public Health 2022, 19, 2695. [Google Scholar] [CrossRef]
- Pulavarty, A.; Horgan, K.; Kakouli-Duarte, T. Effect of an Alltech soil health product on entomopathogenic nematodes, root-knot nematodes and on the growth of tomato plants in the greenhouse. J. Nematol. 2020, 52, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valdes, Y.; Viaene, N.; Moens, M. Effects of yellow mustard amendments on the soil nematode community in a potato field with focus on Globodera rostochiensis. Appl. Soil Ecol. 2012, 59, 39–47. [Google Scholar] [CrossRef]
- Fourie, H.; Ahuja, P.; Lammers, J.; Daneel, M. Brassicacea-based management strategies as an alternative to combat nematode pests: A synopsis. Crop Prot. 2016, 80, 21–41. [Google Scholar] [CrossRef]
- Oka, Y.; Shapira, N.; Fine, P. Control of root-knot nematodes in organic farming systems by organic amendments and soil solarization. Crop Prot. 2007, 26, 1556–1565. [Google Scholar] [CrossRef]
- Peruzzi, A.; Martelloni, L.; Frasconi, C.; Fontanelli, M.; Pirchio, M.; Raffaelli, M. Machines for non-chemical intra-row weed control in narrow and wide-row crops: A review. J. Agric. Eng. 2017, 48, 57–70. [Google Scholar] [CrossRef] [Green Version]
- Expósito, A.; Munera, M.; Giné, A.; López-Gómez, M.; Cáceres, A.; Picó, B.; Gisbert, C.; Medina, V.; Sorribas, F.J. Cucumis metuliferusis resistant to root-knot nematodeMi1.2gene (a)virulent isolates and a promising melon rootstock. Plant Pathol. 2018, 67, 1161–1167. [Google Scholar] [CrossRef] [Green Version]
- Ntalli, N.G.; Caboni, P. Botanical Nematicides: A Review. J. Agric. Food Chem. 2012, 60, 9929–9940. [Google Scholar] [CrossRef]
- Ntalli, N.; Adamski, Z.; Doula, M.; Monokrousos, N. Nematicidal Amendments and Soil Remediation. Plants 2020, 9, 429. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Z.; Liu, M.; Jiang, L.; Chen, X.; Griffiths, B.S.; Li, H.; Hu, F. Vermicompost increases defense against root-knot nematode (Meloidogyne incognita) in tomato plants. Appl. Soil Ecol. 2016, 105, 177–186. [Google Scholar] [CrossRef]
- Akhtar, M.; Malik, A. Roles of organic soil amendments and soil organisms in the biological control of plant-parasitic nematodes: A review. Bioresour. Technol. 2000, 74, 35–47. [Google Scholar] [CrossRef]
- Deguine, J.-P.; Penvern, S. Agroecological Crop Protection in Organic Farming: Relevance and Limits. In Organic Farming, Prototype for Sustainable Agricultures: Prototype for Sustainable Agricultures; Bellon, S., Penvern, S., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 107–130. ISBN 978-94-007-7927-3. [Google Scholar]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furmanczyk, E.M.; Malusà, E. Control of Nematodes in Organic Horticulture Exploiting the Multifunctional Capacity of Microorganisms. Horticulturae 2023, 9, 920. https://doi.org/10.3390/horticulturae9080920
Furmanczyk EM, Malusà E. Control of Nematodes in Organic Horticulture Exploiting the Multifunctional Capacity of Microorganisms. Horticulturae. 2023; 9(8):920. https://doi.org/10.3390/horticulturae9080920
Chicago/Turabian StyleFurmanczyk, Ewa M., and Eligio Malusà. 2023. "Control of Nematodes in Organic Horticulture Exploiting the Multifunctional Capacity of Microorganisms" Horticulturae 9, no. 8: 920. https://doi.org/10.3390/horticulturae9080920