
Hydroponic Production of Selenium-Enriched Baby Leaves of Swiss Chard (Beta vulgaris var. cicla) and Its Wild Ancestor Sea Beet (Beta vulgaris ssp. maritima)

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material, Growth Conditions, Treatments, and Experimental Design
2.2. Determinations
2.3. Estimated Dietary Intake and Health Risk Assessment
2.4. Statistical Analysis
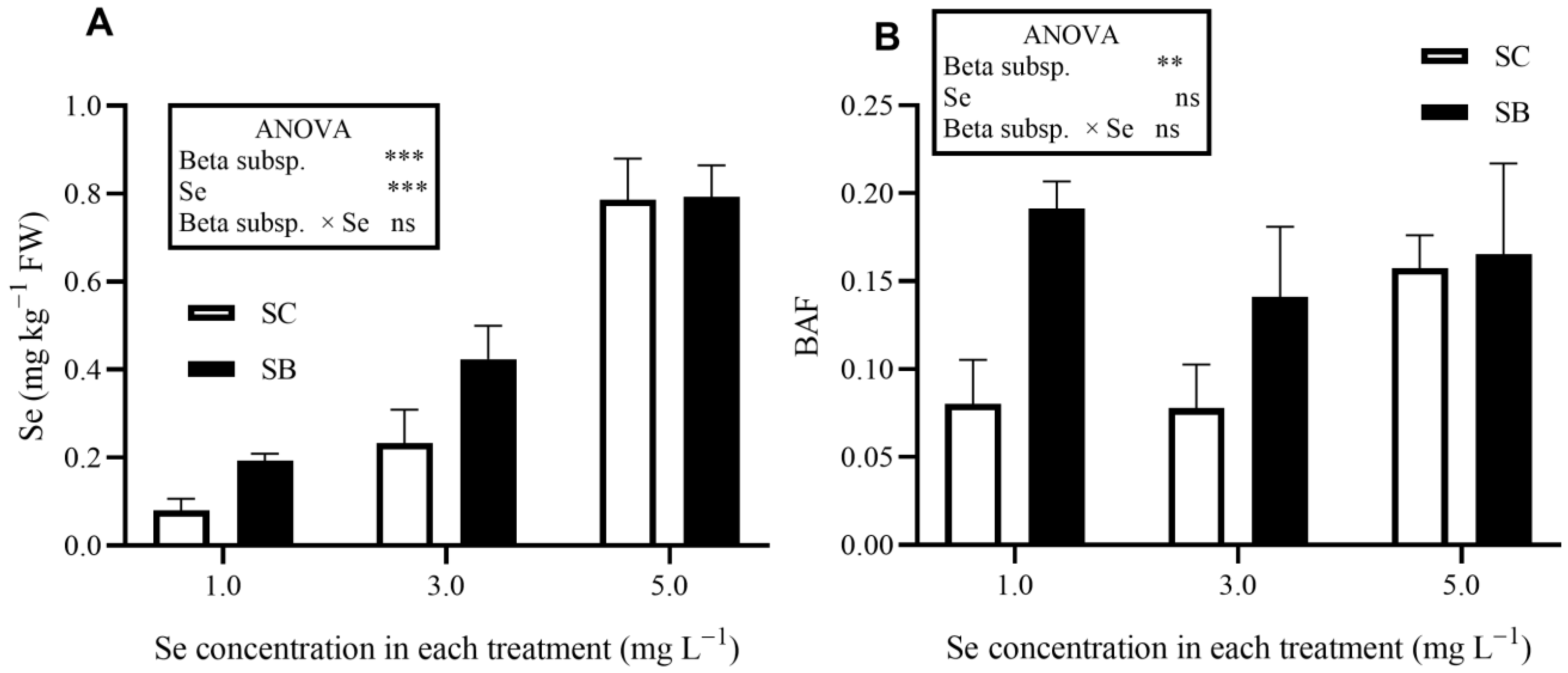
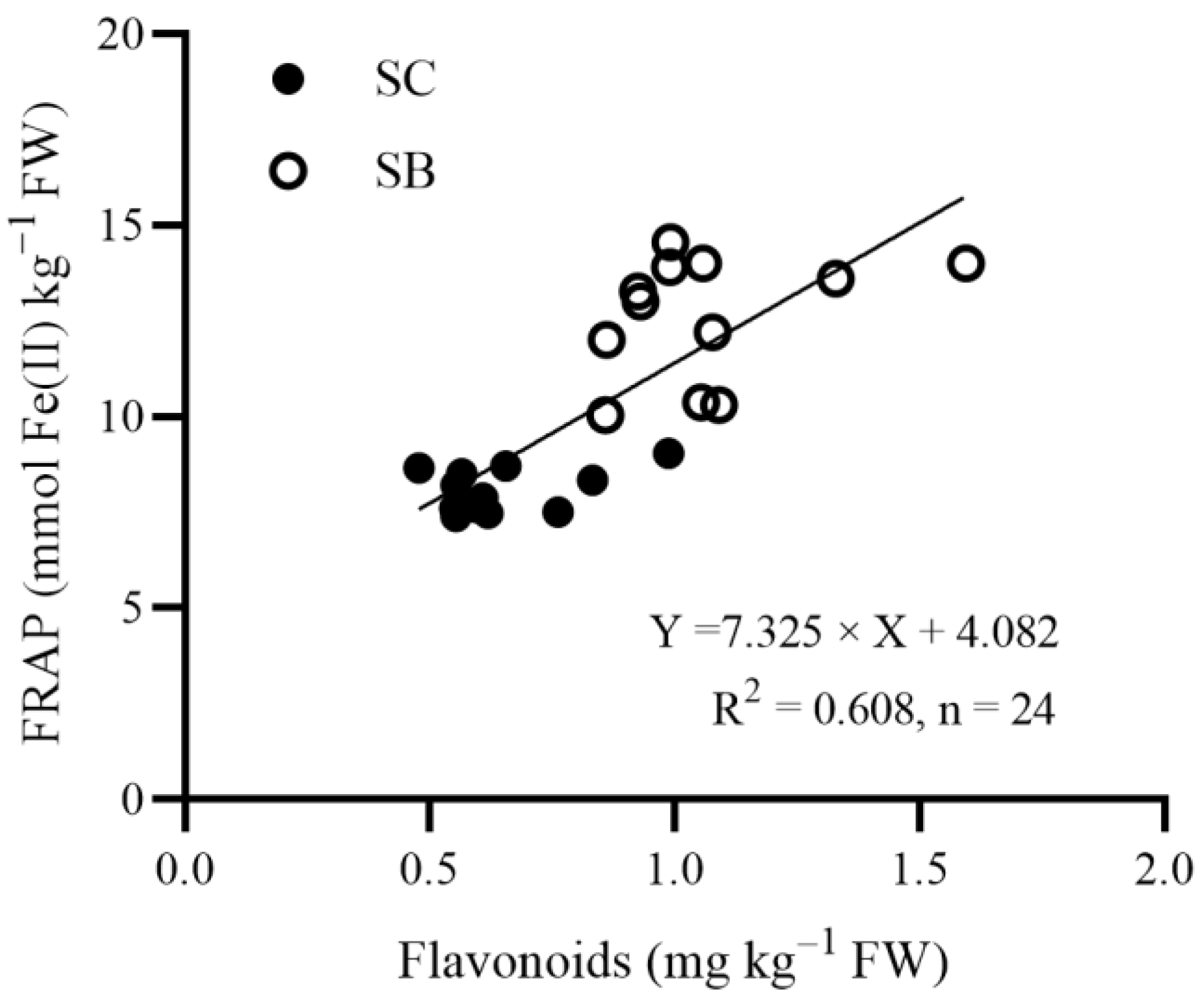
3. Results
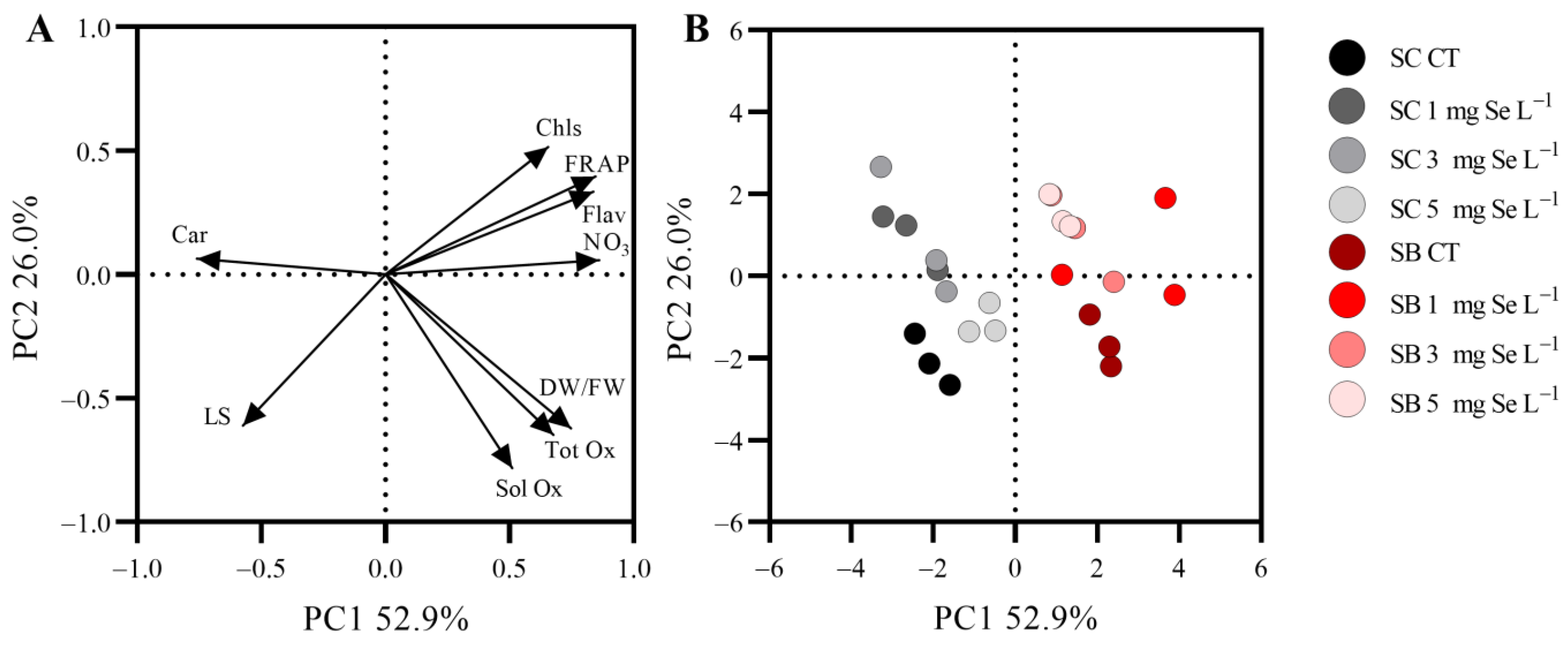
Principal Component Analysis (PCA)
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- White, P.J.; Broadley, M.R. Biofortification of Crops with Seven Mineral Elements Often Lacking in Human Diets—Iron, Zinc, Copper, Calcium, Magnesium, Selenium and Iodine. New Phytol. 2009, 182, 49–84. [Google Scholar] [CrossRef]
- Rayman, M.P. Selenium and Human Health. Lancet 2012, 379, 1256–1268. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority. Scientific Opinion on Dietary Reference Values for Selenium. EFSA J. 2014, 12, 3846. [Google Scholar] [CrossRef]
- European Food Safety Authority. Scientific Opinion on Dietary Reference Values for Iodine. EFSA J. 2014, 12, 3660. [Google Scholar] [CrossRef] [Green Version]
- Gonzali, S.; Kiferle, C.; Perata, P. Iodine Biofortification of Crops: Agronomic Biofortification, Metabolic Engineering and Iodine Bioavailability. Curr. Opin. Biotechnol. 2017, 44, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Consentino, B.B.; Ciriello, M.; Sabatino, L.; Vultaggio, L.; Baldassano, S.; Vasto, S.; Rouphael, Y.; La Bella, S.; De Pascale, S. Current Acquaintance on Agronomic Biofortification to Modulate the Yield and Functional Value of Vegetable Crops: A Review. Horticulturae 2023, 9, 219. [Google Scholar] [CrossRef]
- Izydorczyk, G.; Ligas, B.; Mikula, K.; Witek-Krowiak, A.; Moustakas, K.; Chojnacka, K. Biofortification of Edible Plants with Selenium and Iodine—A Systematic Literature Review. Sci. Total Environ. 2021, 754, 141983. [Google Scholar] [CrossRef]
- Puccinelli, M.; Carmassi, G.; Botrini, L.; Bindi, A.; Rossi, L.; Fierro-sañudo, J.F.; Pardossi, A.; Incrocci, L. Growth and Mineral Relations of Beta vulgaris var. cicla and Beta vulgaris ssp. maritima Cultivated Hydroponically with Diluted Seawater and Low Nitrogen Level in the Nutrient Solution. Horticulturae 2022, 8, 638. [Google Scholar] [CrossRef]
- Puccinelli, M.; Pezzarossa, B.; Rosellini, I.; Malorgio, F. Selenium Enrichment Enhances the Quality and Shelf Life of Basil Leaves. Plants 2020, 9, 801. [Google Scholar] [CrossRef]
- Hajiboland, R.; Amjad, L. Does Antioxidant Capacity of Leaves Play a Role in Growth Response to Selenium at Different Sulfur Nutritional Status? Plant Soil Environ. 2007, 53, 207–215. [Google Scholar] [CrossRef] [Green Version]
- Ramos, S.J.; Faquin, V.; Guilherme, L.R.G.; Castro, E.M.; Ávila, F.W.; Carvalho, G.S.; Bastos, C.E.A.; Oliveira, C. Selenium Biofortification and Antioxidant Activity in Lettuce Plants Fed with Selenate and Selenite. Plant Soil Environ. 2010, 56, 584–588. [Google Scholar] [CrossRef] [Green Version]
- Bian, Z.H.; Lei, B.; Cheng, R.F.; Wang, Y.; Li, T.; Yang, Q.C. Selenium Distribution and Nitrate Metabolism in Hydroponic Lettuce (Lactuca sativa L.): Effects of Selenium Forms and Light Spectra. J. Integr. Agric. 2020, 19, 133–144. [Google Scholar] [CrossRef]
- Hernández-Castro, E.; Trejo-Téllez, L.I.; Gómez-Merino, F.C.; Rodríguez-Mendoza, M.N.; Sánchez-García, P.; Robledo-Paz, A. Bioaccumulation of Iron, Selenium, Nitrate, and Proteins in Chard Shoots. J. Soil Sci. Plant Nutr. 2015, 15, 694–710. [Google Scholar] [CrossRef]
- Puccinelli, M.; Malorgio, F.; Maggini, R.; Rosellini, I.; Pezzarossa, B. Biofortification of Ocimum basilicum L. Plants with Selenium. Acta Hortic. 2019, 1242, 663–670. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Devi, D.D.; Shanker, A.K.; Sheeba, J.A.; Bangarusamy, U. Selenium—An Antioxidative Protectant in Soybean during Senescence. Plant Soil. 2005, 272, 77–86. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B. Enhanced Selenium Content in Sweet Basil (Ocimum basilicum L.) by Foliar Fertilization. Veg. Crops Res. Bull. 2008, 69, 63–72. [Google Scholar] [CrossRef]
- Schiavon, M.; Dall’Acqua, S.; Mietto, A.; Pilon-Smits, E.A.H.; Sambo, P.; Masi, A.; Malagoli, M. Selenium Fertilization Alters the Chemical Composition and Antioxidant Constituents of Tomato (Solanum lycopersicon L.). J. Agric. Food Chem. 2013, 61, 10542–10554. [Google Scholar] [CrossRef]
- Smoleń, S.; Baranski, R.; Ledwożyw-Smoleń, I.; Skoczylas, Ł.; Sady, W. Combined Biofortification of Carrot with Iodine and Selenium. Food Chem. 2019, 300, 125202. [Google Scholar] [CrossRef]
- Motti, R.; Bonanomi, G.; Lanzotti, V.; Sacchi, R. The Contribution of Wild Edible Plants to the Mediterranean Diet: An Ethnobotanical Case Study Along the Coast of Campania (Southern Italy). Econ. Bot. 2020, 74, 249–272. [Google Scholar] [CrossRef]
- Ceccanti, C.; Landi, M.; Guidi, L.; Pardossi, A.; Incrocci, L. Seasonal Fluctuations of Crop Yield, Total Phenolic Content and Antioxidant Activity in Fresh or Cooked Borage (Borago officinalis L.), Mallow (Malva sylvestris L.) and Buck’s-Horn Plantain (Plantago coronopus L.) Leaves. Horticulturae 2022, 8, 253. [Google Scholar] [CrossRef]
- Lombardi, T.; Bertacchi, A.; Pistelli, L.; Pardossi, A.; Pecchia, S.; Toffanin, A.; Sanmartin, C. Biological and Agronomic Traits of the Main Halophytes Widespread in the Mediterranean Region as Potential New Vegetable Crops. Horticulturae 2022, 8, 195. [Google Scholar] [CrossRef]
- Bouchmaa, N.; Ben Mrid, R.; Kabach, I.; Zouaoui, Z.; Karrouchi, K.; Chtibi, H.; Zyad, A.; Cacciola, F.; Nhiri, M. Beta Vulgaris Subsp. Maritima: A Valuable Food with High Added Health Benefits. Appl. Sci. 2022, 12, 1866. [Google Scholar] [CrossRef]
- Saini, R.K.; Ko, E.Y.; Keum, Y.-S. Minimally Processed Ready-to-Eat Baby-Leaf Vegetables: Production, Processing, Storage, Microbial Safety, and Nutritional Potential. Food Rev. Int. 2017, 33, 644–663. [Google Scholar] [CrossRef]
- Petroski, W.; Minich, D.M. Is There Such a Thing as “Anti-Nutrients”? A Narrative Review of Perceived Problematic Plant Compounds. Nutrients 2020, 12, 2929. [Google Scholar] [CrossRef]
- Kiani, A.; Sharafi, K.; Omer, A.K.; Matin, B.K.; Davoodi, R.; Mansouri, B.; Sharafi, H.; Soleimani, H.; Massahi, T.; Ahmadi, E. Accumulation and Human Health Risk Assessment of Nitrate in Vegetables Irrigated with Different Irrigation Water Sources- Transfer Evaluation of Nitrate from Soil to Vegetables. Environ. Res. 2022, 205, 112527. [Google Scholar] [CrossRef]
- Puccinelli, M.; Malorgio, F.; Incrocci, L.; Rosellini, I.; Pezzarossa, B. Effects of Individual and Simultaneous Selenium and Iodine Biofortification of Baby-Leaf Lettuce Plants Grown in Two Different Hydroponic Systems. Horticulturae 2021, 7, 590. [Google Scholar] [CrossRef]
- U.S. EPA. Method 3051A (SW-846): Microwave Assisted Acid Digestion of Sediments, Sludges, and Oils; U.S. EPA: San Francisco, CA, USA, 2007. [Google Scholar]
- Cataldo, D.A.; Maroon, M.; Schrader, L.E.; Youngs, V.L. Rapid Colorimetric Determination of Nitrate in Plant Tissue by Nitration of Salicylic Acid. Commun. Soil. Sci. Plant Anal. 1975, 6, 71–80. [Google Scholar] [CrossRef]
- Naik, V.V.; Mahavidyalaya, S.P.K.; Sindhudurg, D. Methodology in Determination of Oxalic Acid in Plnat Tissue: A Comparative Approach. J. Glob. Trends Pharm. Sci. 2014, 5, 1662–1672. [Google Scholar]
- Wellburn, A.R.; Lichtenthaler, H. Formulae and Program to Determine Total Carotenoids and Chlorophylls A and B of Leaf Extracts in Different Solvents. In Advances in Photosynthesis Research; Springer: Berlin/Heidelberg, Germany, 1984; pp. 9–12. [Google Scholar]
- Kim, D.O.; Chun, O.K.; Kim, Y.J.; Moon, H.Y.; Lee, C.Y. Quantification of Polyphenolics and Their Antioxidant Capacity in Fresh Plums. J. Agric. Food Chem. 2003, 51, 6509–6515. [Google Scholar] [CrossRef]
- Kang, H.M.; Saltveit, M.E. Antioxidant Capacity of Lettuce Leaf Tissue Increases after Wounding. J. Agric. Food Chem. 2002, 50, 7536–7541. [Google Scholar] [CrossRef]
- Benzie, I.F.F.; Strain, J. The Ferric Reducing Ability of Plasma (FRAP) as a Measure of “Antioxidant Power”: The FRAP Assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Food Safety Authority. Tolerable Upper Intake Level on Vitamins and Minerals. Sci. Comm. Food 2006, 33, 480. [Google Scholar]
- European Food Safety Authority. Nitrate in Vegetables—Scientific Opinion of the Panel on Contaminants in the Food Chain. EFSA J. 2008, 6, 689. [Google Scholar] [CrossRef]
- Libert, B.; Franceschi, V.R. Oxalate in Crop Plants. J. Agric. Food Chem. 1987, 35, 926–938. [Google Scholar] [CrossRef]
- Puccinelli, M.; Malorgio, F.; Rosellini, I.; Pezzarossa, B. Uptake and Partitioning of Selenium in Basil (Ocimum basilicum L.) Plants Grown in Hydroponics. Sci. Hortic. 2017, 225, 271–276. [Google Scholar] [CrossRef]
- Francini, A.; Quattrini, E.; Giuffrida, F.; Ferrante, A. Biofortification of Baby Leafy Vegetables Using Nutrient Solution Containing Selenium. J. Sci. Food Agric. 2023, 103, 5472–5480. [Google Scholar] [CrossRef]
- Duarte, B.; Feijão, E.; Pinto, M.V.; Matos, A.R.; Silva, A.; Figueiredo, A.; Fonseca, V.F.; Reis-Santos, P.; Caçador, I. Nutritional Valuation and Food Safety of Endemic Mediterranean Halophytes Species Cultivated in Abandoned Salt Pans under a Natural Irrigation Scheme. Estuar. Coast. Shelf Sci. 2022, 265, 107733. [Google Scholar] [CrossRef]
- Ferrarese, M.; Mahmoodi Sourestani, M.; Quattrini, E.; Schiavi, M.; Ferrante, A. Biofortification of Spinach Plants Applying Selenium in the Nutrient Solution of Floating System. Veg. Crops Res. Bull. 2012, 76, 127–136. [Google Scholar] [CrossRef]
- Wang, C.; Yue, L.; Cheng, B.; Chen, F.; Zhao, X.; Wang, Z.; Xing, B. Mechanisms of Growth-Promotion and Se-Enrichment in Brassica chinensis L. by Selenium Nanomaterials: Beneficial Rhizosphere Microorganisms, Nutrient Availability, and Photosynthesis. Environ. Sci. Nano 2022, 9, 302–312. [Google Scholar] [CrossRef]
- Hawrylak-Nowak, B.; Matraszek, R.; Pogorzelec, M. The Dual Effects of Two Inorganic Selenium Forms on the Growth, Selected Physiological Parameters and Macronutrients Accumulation in Cucumber Plants. Acta Physiol. Plant. 2015, 37, 41. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Grimm, B. Connecting Chlorophyll Metabolism with Accumulation of the Photosynthetic Apparatus. Trends Plant Sci. 2021, 26, 484–495. [Google Scholar] [CrossRef]
- Skorupa, M.; Gołȩbiewski, M.; Kurnik, K.; Niedojadło, J.; Kȩsy, J.; Klamkowski, K.; Wójcik, K.; Treder, W.; Tretyn, A.; Tyburski, J. Salt Stress vs. Salt Shock—The Case of Sugar Beet and Its Halophytic Ancestor. BMC Plant Biol. 2019, 19, 57. [Google Scholar] [CrossRef] [Green Version]
- Saeedi, M.; Soltani, F.; Babalar, M.; Izadpanah, F.; Wiesner-Reinhold, M.; Baldermann, S. Selenium Fortification Alters the Growth, Antioxidant Characteristics and Secondary Metabolite Profiles of Cauliflower (Brassica oleracea var. botrytis) Cultivars in Hydroponic Culture. Plants 2021, 10, 1537. [Google Scholar] [CrossRef]
- Shalaby, T.A.; Abd-Alkarim, E.; El-Aidy, F.; Hamed, E.S.; Sharaf-Eldin, M.; Taha, N.; El-Ramady, H.; Bayoumi, Y.; dos Reis, A.R. Nano-Selenium, Silicon and H2O2 Boost Growth and Productivity of Cucumber under Combined Salinity and Heat Stress. Ecotoxicol. Environ. Saf. 2021, 212, 111962. [Google Scholar] [CrossRef]
- Sabatino, L.; Ntatsi, G.; Iapichino, G.; D’anna, F.; De Pasqual, C. Effect of Selenium Enrichment and Type of Application on Yield, Functional Quality and Mineral Composition of Curly Endive Grown in a Hydroponic System. Agronomy 2019, 9, 207. [Google Scholar] [CrossRef] [Green Version]
- Mishra, V.K.; Gamage, T.V. Postharvest Physiology of Fruits and Vegetables. In Handbook of Food Preservation; CRC Press: Boca Raton, FL, USA, 2020; p. 20. ISBN 9780128132784. [Google Scholar]
- European Parliament and Council of the European Union. Commission Regulation (EU) No 1258/2011 of 2 December 2011 Amending Regulation (EC) No 1881/2006 as Regards Maximum Levels for Nitrates in Foodstuffs. Off. J. Eur. Union 2011, 320, 15–17. [Google Scholar]
- Colla, G.; Kim, H.J.; Kyriacou, M.C.; Rouphael, Y. Nitrate in Fruits and Vegetables. Sci. Hortic. 2018, 237, 221–238. [Google Scholar] [CrossRef]
- Golob, A.; Stibilj, V.; Nečemer, M.; Kump, P.; Kreft, I.; Hočevar, A.; Gaberščik, A.; Germ, M. Calcium Oxalate Druses Affect Leaf Optical Properties in Selenium-Treated Fagopyrum Tataricum. J. Photochem. Photobiol. B 2018, 180, 51–55. [Google Scholar] [CrossRef]
- Freidig, A.K.; Goldman, I.L. Variation in Oxalic Acid Content among Commercial Table Beet Cultivars and Related Crops. J. Am. Soc. Hortic. Sci. 2011, 136, 54–60. [Google Scholar] [CrossRef] [Green Version]
- Chang, D.C.; Park, C.S.; Kim, S.Y.; Lee, Y.B. Growth and Tuberization of Hydroponically Grown Potatoes. Potato Res. 2012, 55, 69–81. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Kopsell, D.E.; Curran-Celentano, J. Carotenoid Pigments in Kale Are Influenced by Nitrogen Concentration and Form. J. Sci. Food Agric. 2007, 87, 900–907. [Google Scholar] [CrossRef]
- Lebedev, V.G.; Lebedeva, T.N.; Vidyagina, E.O.; Sorokopudov, V.N.; Popova, A.A.; Shestibratov, K.A. Relationship between Phenolic Compounds and Antioxidant Activity in Berries and Leaves of Raspberry Genotypes and Their Genotyping by SSR Markers. Antioxidants 2022, 11, 1961. [Google Scholar] [CrossRef] [PubMed]
- Muflihah, Y.M.; Gollavelli, G.; Ling, Y.C. Correlation Study of Antioxidant Activity with Phenolic and Flavonoid Compounds in 12 Indonesian Indigenous Herbs. Antioxidants 2021, 10, 1530. [Google Scholar] [CrossRef]
- Aryal, S.; Baniya, M.K.; Danekhu, K.; Kunwar, P.; Gurung, R.; Koirala, N. Total Phenolic Content, Flavonoid Content and Antioxidant Potential of Wild Vegetables from Western Nepal. Plants 2019, 8, 96. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Pare, P.W.; Zhang, J.; Kang, T.; Zhang, Z.; Yang, D.; Wang, K.; Xing, H. Antioxidant Capacity Connection with Phenolic and Flavonoid Content in Chinese Medicinal Herbs. Rec. Nat. Prod. 2018, 12, 239–250. [Google Scholar] [CrossRef]
- Mezeyova, I.; Hegedusova, A.; Andrejiová, A.; Mezeyová, I.; Hegedűsová, A.; Hegedűs, O.; Golian, M. Phytomass and Content of Essential Oils in Ocimum basilicum after Foliar Treatment with Selenium. Agric. Food 2016, 4, 19–27. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Sowing Date | 29 September 2021 |
| Transplant date | 22 October 2021 |
| Start of treatment | 25 October 2021 |
| Harvest date | 7 November 2021 |
| Days of treatment | 8 (11) * |
| Mean air temperature (°C) | 20.5 ** |
| Mean daily solar radiation (MJ m−2 day−1) | 7.6 ** |
| Cumulative solar radiation (MJ m−2) | 91.0 ** |
| Beta Subsp. | Se Concentration | Yield | Leaf DM | DM/FM | LAI | LS |
|---|---|---|---|---|---|---|
| (mg L−1) | kg m−2 | g m−2 | % | kg m−2 | ||
| Swiss chard | 0 | 2.13 | 64.26 | 3.01 abc | 2.80 | 0.761 a |
| 1 | 2.68 | 50.57 | 1.89 c | 5.48 | 0.489 b | |
| 3 | 2.44 | 51.37 | 2.09 bc | 4.79 | 0.511 b | |
| 5 | 2.40 | 74.97 | 3.14 ab | 4.75 | 0.504 b | |
| Sea beet | 0 | 1.81 | 67.75 | 3.76 a | 3.65 | 0.495 b |
| 1 | 2.05 | 66.61 | 3.29 a | 4.95 | 0.413 c | |
| 3 | 1.66 | 51.87 | 3.06 abc | 4.46 | 0.373 c | |
| 5 | 1.90 | 56.20 | 2.94 abc | 4.62 | 0.411 c | |
| Mean effect | ||||||
| Swiss chard | 2.41 a | 60.29 | 2.53 b | 4.46 | 0.566 a | |
| Sea beet | 1.85 b | 60.61 | 3.26 a | 4.42 | 0.423 b | |
| 0 | 1.97 b | 66.00 | 3.38 a | 3.23 b | 0.628 a | |
| 1 | 2.36 a | 58.59 | 2.59 b | 5.22 a | 0.451 b | |
| 3 | 2.05 ab | 51.62 | 2.58 b | 4.62 a | 0.442 b | |
| 5 | 2.15 ab | 65.58 | 3.04 ab | 4.69 a | 0.457 b | |
| ANOVA | ||||||
| Beta subsp. | *** | ns | *** | ns | *** | |
| Se concentration | * | ns | * | *** | *** | |
| Beta subsp. × Se | ns | ns | * | ns | *** | |
| Beta Subsp. | Se Concentration | Nitrate | Total Oxalate | Soluble Oxalate | Chlorophylls | Carotenoids | Phenols | Flavonoids | FRAP Index |
|---|---|---|---|---|---|---|---|---|---|
| mg L−1 | mg kg−1 | mmol Fe(II)kg−1 | |||||||
| Swiss chard | 0 | 787.9 d | 7329.4 | 5972.5 ab | 748.0 | 114.9 | 1.200 | 0.584 | 7.70 c |
| 1 | 915.5 cd | 4950.6 | 3921.4 b | 802.3 | 137.1 | 1.070 | 0.645 | 7.46 c | |
| 3 | 1189.4 c | 4527.3 | 3991.5 ab | 786.2 | 137.5 | 1.073 | 0.650 | 8.35 c | |
| 5 | 1275.1 c | 7667.1 | 6615.3 ab | 815.5 | 160.6 | 1.041 | 0.706 | 8.80 bc | |
| Sea beet | 0 | 2446.8 a | 9043.6 | 6960.9 a | 771.0 | 30.1 | 1.208 | 1.000 | 10.24 b |
| 1 | 1900.4 b | 8278.5 | 6503.5 ab | 1101.7 | 44.1 | 1.106 | 1.261 | 13.20 a | |
| 3 | 1859.8 b | 7157.1 | 5096.2 ab | 963.0 | 76.7 | 1.185 | 1.000 | 13.26 a | |
| 5 | 1797.3 b | 6126.3 | 4397.2 ab | 988.0 | 91.4 | 1.486 | 0.990 | 13.72 a | |
| Mean effect | |||||||||
| Swiss chard | 1042.0 b | 6118.6 b | 5125.2 | 788.0 b | 137.5 a | 1.096 b | 0.646 | 8.08 b | |
| Sea beet | 2001.1 a | 7651.4 a | 5739.4 | 955.9 a | 60.6 b | 1.246 a | 1.063 | 12.61 a | |
| 0 | 1617.3 | 8186.5 | 6466.7 a | 759.5 b | 72.5 b | 1.204 | 0.792 | 9.0 b | |
| 1 | 1407.9 | 6614.6 | 5212.4 ab | 952.0 a | 90.6 ab | 1.088 | 0.953 | 10.3 a | |
| 3 | 1524.6 | 5842.2 | 4543.9 b | 874.6 ab | 107.1 ab | 1.129 | 0.825 | 10.8 a | |
| 5 | 1536.2 | 6896.7 | 5506.2 ab | 901.7 ab | 126.0 a | 1.264 | 0.848 | 11.3 a | |
| ANOVA | |||||||||
| Beta subsp. | *** | * | ns | *** | *** | *** | ns | *** | |
| Se concentration | ns | ns | * | * | ** | ns | ns | *** | |
| Beta subsp. × Se | *** | ns | ** | ns | ns | ns | ns | ** | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Puccinelli, M.; Rosellini, I.; Malorgio, F.; Pardossi, A.; Pezzarossa, B. Hydroponic Production of Selenium-Enriched Baby Leaves of Swiss Chard (Beta vulgaris var. cicla) and Its Wild Ancestor Sea Beet (Beta vulgaris ssp. maritima). Horticulturae 2023, 9, 909. https://doi.org/10.3390/horticulturae9080909
Puccinelli M, Rosellini I, Malorgio F, Pardossi A, Pezzarossa B. Hydroponic Production of Selenium-Enriched Baby Leaves of Swiss Chard (Beta vulgaris var. cicla) and Its Wild Ancestor Sea Beet (Beta vulgaris ssp. maritima). Horticulturae. 2023; 9(8):909. https://doi.org/10.3390/horticulturae9080909
Chicago/Turabian StylePuccinelli, Martina, Irene Rosellini, Fernando Malorgio, Alberto Pardossi, and Beatrice Pezzarossa. 2023. "Hydroponic Production of Selenium-Enriched Baby Leaves of Swiss Chard (Beta vulgaris var. cicla) and Its Wild Ancestor Sea Beet (Beta vulgaris ssp. maritima)" Horticulturae 9, no. 8: 909. https://doi.org/10.3390/horticulturae9080909