
Enhancing Horticultural Crops through Genome Editing: Applications, Benefits, and Considerations
 , ,
, ,
Abstract
:1. Introduction to Genome Editing in Agriculture
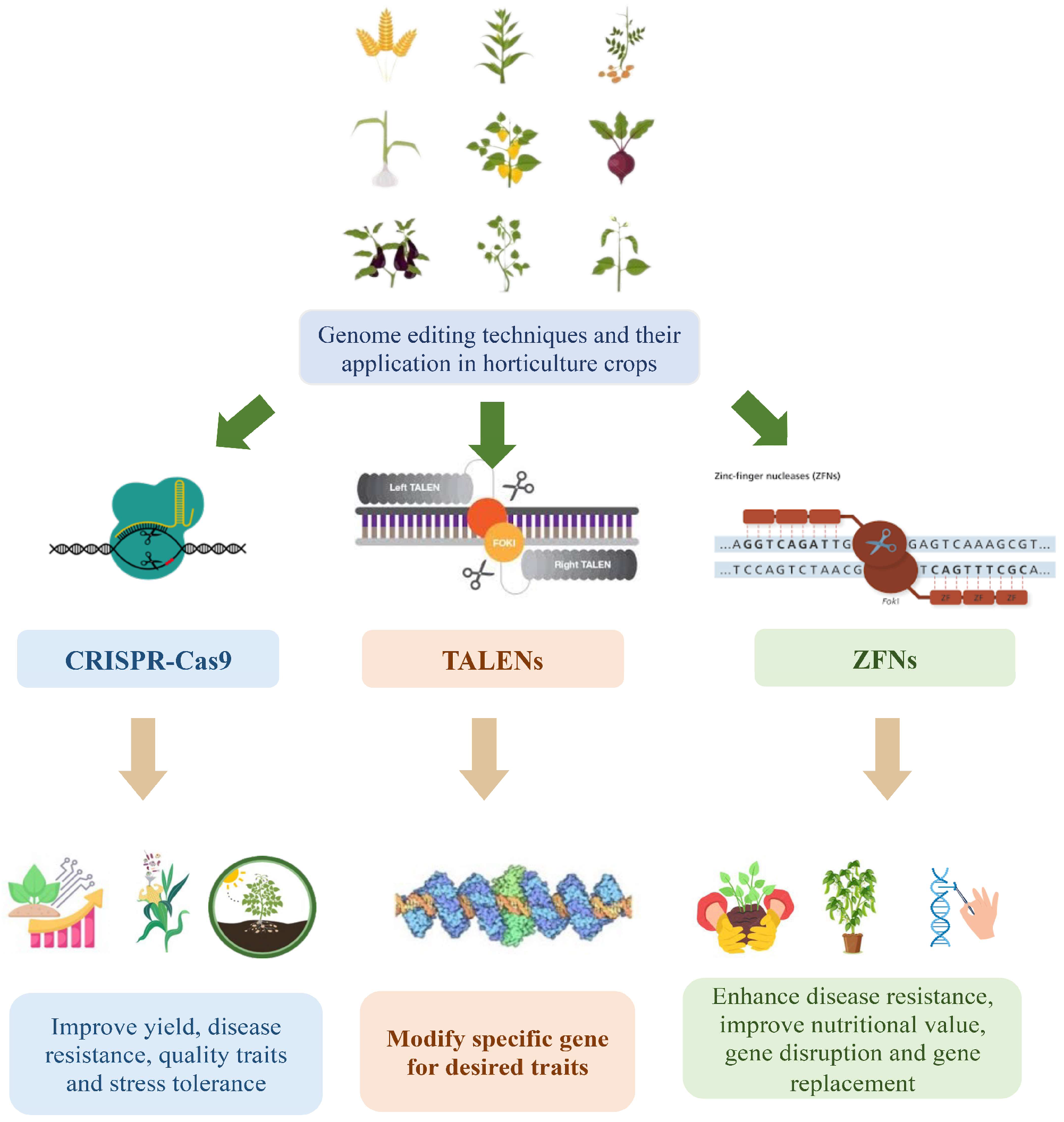
2. CRISPR-Cas9 in Horticultural Crops
2.1. Structural Components and Mechanisms of CRISPR-Cas9Subsection Applications of CRISPR-Cas9 in Horticultural Crops
2.2. Applications of CRISPR-Cas9 in Horticultural Crops
2.2.1. Trait Modification
2.2.2. Gene Knockout
2.2.3. Gene Activation/Suppression
2.2.4. Genome Engineering for Crop Domestication
2.2.5. Genome Editing for Quality Improvement
2.3. Potential Challenges Associated with CRISPR-Cas9 in Horticultural Crops
2.3.1. Off-Target Effects
2.3.2. Delivery and Transformation Efficiency
2.3.3. Off-Target Effects in Non-Coding Regions
2.3.4. Inheritance and Segregation of CRISPR-Edited Traits
2.4. Advancements in CRISPR-Cas9 to Overcome Initial Limitations
3. TALENs (Transcription Activator-like Effector Nucleases)
3.1. Applications of TALENs in Horticultural Crops
3.1.1. Gene Knockout
3.1.2. Gene Editing
3.2. Limitations of TALENs in Horticultural Crops
3.2.1. Design Complexity
3.2.2. Delivery and Transformation Efficiency
3.3. Advancements in TALENs to Overcome Limitations in Horticultural Crops
4. ZFNs (Zinc Finger Nucleases)
4.1. ZFNs and Their Applications
4.2. Challenges Associated with ZFNs in Horticultural Crops
4.3. Advancements in ZFNs Overcoming Challenges in Horticultural Crop Genome Editing
5. Regulatory and Ethical Considerations
5.1. Regulatory Frameworks for Genome-Edited Crops
5.2. Safety Assessment and Risk Analysis
5.3. Public Perception and Ethical Considerations
5.4. International Harmonization and Collaboration
6. Future Prospects of Genome-Edited Crops in Horticulture
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Okunlola, A.I.; Adepoju, A.O.; Akinpetide, E.O. The significant role of horticulture in environmental aesthetics and management. Int. J. Hortic. 2016, 6, 17. [Google Scholar] [CrossRef]
- Salgotra, R.K.; Chauhan, B.S. Genetic Diversity, Conservation, and Utilization of Plant Genetic Resources. Genes 2023, 14, 174. [Google Scholar] [CrossRef]
- Borlaug, N.E. Contributions of Conventional Plant-Breeding to Food-Production. Science 1983, 219, 689–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, H.C.; Crouch, J.H.; Sharma, K.K.; Seetharama, N.; Hash, C.T. Applications of biotechnology for crop improvement: Prospects and constraints. Plant Sci. 2002, 163, 381–395. [Google Scholar] [CrossRef] [Green Version]
- Beaver, J.S.; Osorno, J.M. Achievements and limitations of contemporary common bean breeding using conventional and molecular approaches. Euphytica 2009, 168, 145–175. [Google Scholar] [CrossRef]
- Xiong, J.S.; Ding, J.; Li, Y. Genome-editing technologies and their potential application in horticultural crop breeding. Hortic. Res. 2015, 2, 15019. [Google Scholar] [CrossRef] [Green Version]
- Doudna, J.A.; Charpentier, E. The new frontier of genome engineering with CRISPR-Cas9. Science 2014, 346, 1258096. [Google Scholar] [CrossRef]
- Thurtle-Schmidt, D.M.; Lo, T.W. Molecular biology at the cutting edge: A review on CRISPR/CAS9 gene editing for undergraduates. Biochem. Mol. Biol. Edu. 2018, 46, 195–205. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.M.; Hua, K.; Lang, Z.B. Genome editing for horticultural crop improvement. Hortic. Res. 2019, 6, 113. [Google Scholar] [CrossRef] [Green Version]
- Erpen-Dalla Corte, L.; Mahmoud, L.M.; Moraes, T.S.; Mou, Z.L.; Grosser, J.W.; Dutt, M. Development of Improved Fruit, Vegetable, and Ornamental Crops Using the CRISPR/Cas9 Genome Editing Technique. Plants 2019, 8, 601. [Google Scholar] [CrossRef] [Green Version]
- Zhang, D.Q.; Zhang, Z.Y.; Unver, T.; Zhang, B.H. CRISPR/Cas: A powerful tool for gene function study and crop improvement. J. Adv. Res. 2021, 29, 207–221. [Google Scholar] [CrossRef] [PubMed]
- Rani, R.; Yadav, P.; Barbadikar, K.M.; Baliyan, N.; Malhotra, E.V.; Singh, B.K.; Kumar, A.; Singh, D. CRISPR/Cas9: A promising way to exploit genetic variation in plants. Biotechnol. Lett. 2016, 38, 1991–2006. [Google Scholar] [CrossRef] [PubMed]
- Gaj, T.; Gersbach, C.A.; Barbas, C.F. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering. Trends Biotechnol. 2013, 31, 397–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, N.; Zhao, H.M. Transcription activator-like effector nucleases (TALENs): A highly efficient and versatile tool for genome editing. Biotechnol. Bioeng. 2013, 110, 1811–1821. [Google Scholar] [CrossRef]
- Joung, J.K.; Sander, J.D. INNOVATION TALENs: A widely applicable technology for targeted genome editing. Nat. Rev. Mol. Cell Bio. 2013, 14, 49–55. [Google Scholar] [CrossRef] [Green Version]
- Li, T.; Huang, S.; Jiang, W.Z.; Wright, D.; Spalding, M.H.; Weeks, D.P.; Yang, B. TAL nucleases (TALNs): Hybrid proteins composed of TAL effectors and FokI DNA-cleavage domain. Nucleic Acids Res. 2011, 39, 359–372. [Google Scholar] [CrossRef] [Green Version]
- Pattanayak, V.; Ramirez, C.L.; Joung, J.K.; Liu, D.R. Revealing off-target cleavage specificities of zinc-finger nucleases by in vitro selection. Nat. Methods 2011, 8, 765–770. [Google Scholar] [CrossRef] [Green Version]
- Osakabe, Y.; Osakabe, K. Genome Editing with Engineered Nucleases in Plants. Plant Cell Physiol. 2015, 56, 389–400. [Google Scholar] [CrossRef] [Green Version]
- Bhagwat, A.C.; Patil, A.M.; Saroj, S.D. CRISPR/Cas 9-Based Editing in the Production of Bioactive Molecules. Mol. Biotechnol. 2022, 64, 245–251. [Google Scholar] [CrossRef]
- Khanzadi, M.N.; Khan, A.A. CRISPR/Cas9: Nature’s gift to prokaryotes and an auspicious tool in genome editing. J. Basic Microb. 2020, 60, 91–102. [Google Scholar] [CrossRef]
- Noman, A.; Aqeel, M.; He, S.L. CRISPR-Cas9: Tool for Qualitative and Quantitative Plant Genome Editing. Front. Plant Sci. 2016, 7, 1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, M.J.; Wang, L.Q. CRISPR/Cas9 technology for improving agronomic traits and future prospective in agriculture. Planta 2021, 254, 68. [Google Scholar] [CrossRef]
- Rasheed, A.; Barqawi, A.A.; Mahmood, A.; Nawaz, M.; Shah, A.N.; Bay, D.H.; Alahdal, M.A.; Hassan, M.U.; Qari, S.H. CRISPR/Cas9 is a powerful tool for precise genome editing of legume crops: A review. Mol. Biol. Rep. 2022, 49, 5595–5609. [Google Scholar] [CrossRef] [PubMed]
- Urnov, F.D.; Rebar, E.J.; Holmes, M.C.; Zhang, H.S.; Gregory, P.D. Genome editing with engineered zinc finger nucleases. Nat. Rev. Genet. 2010, 11, 636–646. [Google Scholar] [CrossRef] [PubMed]
- Wai, A.H.; Naing, A.H.; Lee, D.J.; Kim, C.K.; Chung, M.Y. Molecular genetic approaches for enhancing stress tolerance and fruit quality of tomato. Plant Biotechnol. Rep. 2020, 14, 515–537. [Google Scholar] [CrossRef]
- Gonzales, L.R.; Shi, L.; Bergonzi, S.B.; Oortwijn, M.; Franco-Zorrilla, J.M.; Solano-Tavira, R.; Visser, R.G.F.; Abelenda, J.A.; Bachem, C.W.B. Potato CYCLING DOF FACTOR 1 and its lncRNA counterpart StFLORE link tuber development and drought response. Plant J. 2021, 105, 855–869. [Google Scholar] [CrossRef]
- Henry, R.J.; Furtado, A.; Rangan, P. Wheat seed transcriptome reveals genes controlling key traits for human preference and crop adaptation. Curr. Opin. Plant Biol. 2018, 45, 231–236. [Google Scholar] [CrossRef] [Green Version]
- Yang, W.; Ren, J.; Liu, W.; Liu, D.; Xie, K.; Zhang, F.; Wang, P.; Guo, W.; Wu, X. An efficient transient gene expression system for protein subcellular localization assay and genome editing in citrus protoplasts. Hortic. Plant J. 2023, 9, 425–436. [Google Scholar] [CrossRef]
- Martin-Pizarro, C.; Trivino, J.C.; Pose, D. Functional analysis of the TM6 MADS-box gene in the octoploid strawberry by CRISPR/Cas9-directed mutagenesis. J. Exp. Bot. 2019, 70, 885–895. [Google Scholar] [CrossRef] [Green Version]
- Capriotti, L.; Baraldi, E.; Mezzetti, B.; Limera, C.; Sabbadini, S. Biotechnological Approaches: Gene Overexpression, Gene Silencing, and Genome Editing to Control Fungal and Oomycete Diseases in Grapevine. Int. J. Mol. Sci. 2020, 21, 5701. [Google Scholar] [CrossRef]
- Afrin, K.S.; Rahim, M.A.; Jung, H.J.; Park, J.I.; Kim, H.T.; Nou, I.S. Development of Molecular Marker through Genome Realignment for Specific Detection of Xanthomonas campestris pv. campestris Race 5, a Pathogen of Black Rot Disease. J. Microbiol. Biotechnol. 2019, 29, 785–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waltz, E. CRISPR-edited crops free to enter market, skip regulation. Nat. Biotechnol. 2016, 34, 582. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.H.; Sheng, O.; Deng, G.M.; He, W.D.; Dong, T.; Yang, Q.S.; Dou, T.X.; Li, C.Y.; Gao, H.J.; Liu, S.W.; et al. CRISPR/Cas9-mediated genome editing of MaACO1 (aminocyclopropane-1-carboxylate oxidase 1) promotes the shelf life of banana fruit. Plant Biotechnol. J. 2021, 19, 654–656. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Deng, Y.J.; Liu, J.X.; Duan, A.Q.; Liu, H.; Xiong, A.S. DcCCD4 catalyzes the degradation of alpha-carotene and beta-carotene to affect carotenoid accumulation and taproot color in carrot. Plant J. 2021, 108, 1116–1130. [Google Scholar] [CrossRef] [PubMed]
- Abdullah; Faraji, S.; Mehmood, F.; Malik, H.M.T.; Ahmed, I.; Heidari, P.; Poczai, P. The GASA Gene Family in Cacao (Theobroma cacao, Malvaceae): Genome Wide Identification and Expression Analysis. Agronomy 2021, 11, 1425. [Google Scholar] [CrossRef]
- Nonaka, S.; Ito, M.; Ezura, H. Targeted modification of CmACO1 by CRISPR/Cas9 extends the shelf-life of Cucumis melo var. reticulatus melon. Front. Genome Ed. 2023, 5, 1176125. [Google Scholar] [CrossRef]
- Mishra, R.; Mohanty, J.N.; Mahanty, B.; Joshi, R.K. A single transcript CRISPR/Cas9 mediated mutagenesis of CaERF28 confers anthracnose resistance in chilli pepper (Capsicum annuum L.). Planta 2021, 254, 5. [Google Scholar] [CrossRef]
- Wang, C.P.; Li, Y.; Wang, N.; Yu, Q.; Li, Y.H.; Gao, J.P.; Zhou, X.F.; Ma, N. An efficient CRISPR/Cas9 platform for targeted genome editing in rose (Rosa hybrida). J. Integr. Plant Biol. 2023, 65, 895–899. [Google Scholar] [CrossRef]
- Pechar, G.S.; Donaire, L.; Gosalvez, B.; Garcia-Almodovar, C.; Sanchez-Pina, M.A.; Truniger, V.; Aranda, M.A. Editing melon eIF4E associates with virus resistance and male sterility. Plant Biotechnol. J. 2022, 20, 2006–2022. [Google Scholar] [CrossRef]
- Wang, L.; Chen, L.; Li, R.; Zhao, R.R.; Yang, M.J.; Sheng, J.P.; Shen, L. Reduced Drought Tolerance by CRISPR/Cas9-Mediated SlMAPK3 Mutagenesis in Tomato Plants. J. Agr. Food Chem. 2017, 65, 8674–8682. [Google Scholar] [CrossRef]
- Okuzaki, A.; Ogawa, T.; Koizuka, C.; Kaneko, K.; Inaba, M.; Imamura, J.; Koizuka, N. CRISPR/Cas9-mediated genome editing of the fatty acid desaturase 2 gene in Brassica napus. Plant Physiol. Biochem. 2018, 131, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Herath, D.; Voogd, C.; Mayo-Smith, M.; Yang, B.; Allan, A.C.; Putterill, J.; Varkonyi-Gasic, E. CRISPR-Cas9-mediated mutagenesis of kiwifruit BFT genes results in an evergrowing but not early flowering phenotype. Plant Biotechnol. J. 2022, 20, 2064–2076. [Google Scholar] [CrossRef]
- Shu, P.; Li, Z.Y.; Min, D.D.; Zhang, X.H.; Ai, W.; Li, J.Z.; Zhou, J.X.; Li, Z.L.; Li, F.J.; Li, X.A. CRISPR/Cas9-Mediated SlMYC2 Mutagenesis Adverse to Tomato Plant Growth and MeJA-Induced Fruit Resistance to Botrytis cinerea. J. Agr. Food Chem. 2020, 68, 5529–5538. [Google Scholar] [CrossRef]
- Ma, J.; Sun, S.; Whelan, J.; Shou, H.X. CRISPR/Cas9-Mediated Knockout of GmFATB1 Significantly Reduced the Amount of Saturated Fatty Acids in Soybean Seeds. Int. J. Mol. Sci. 2021, 22, 3877. [Google Scholar] [CrossRef]
- Wang, R. What Makes ‘Hayward’ Kiwifruit Store so Well? The Biological Basis for the Postharvest Behaviour of ‘Hayward’ Kiwifruit. Ph.D. Thesis, The University of Auckland, Auckland, New Zealand, 2021. [Google Scholar]
- Wang, H.X.; Wu, Y.L.; Zhang, Y.D.; Yang, J.; Fan, W.J.; Zhang, H.; Zhao, S.S.; Yuan, L.; Zhang, P. CRISPR/Cas9-Based Mutagenesis of Starch Biosynthetic Genes in Sweet Potato (Ipomoea Batatas) for the Improvement of Starch Quality. Int. J. Mol. Sci. 2019, 20, 4702. [Google Scholar] [CrossRef] [Green Version]
- Brewer, S.E.; Chambers, A.H. CRISPR/Cas9-mediated genome editing of phytoene desaturase in Carica papaya L. J. Hortic. Sci. Biotechnol. 2022, 97, 580–592. [Google Scholar] [CrossRef]
- Maioli, A.; Gianoglio, S.; Moglia, A.; Acquadro, A.; Valentino, D.; Milani, A.M.; Prohens, J.; Orzaez, D.; Granell, A.; Lanteri, S.; et al. Simultaneous CRISPR/Cas9 Editing of Three PPO Genes Reduces Fruit Flesh Browning in Solanum melongena L. Front. Plant Sci. 2020, 11, 607161. [Google Scholar] [CrossRef]
- Gomez, M.A.; Lin, Z.D.; Moll, T.; Chauhan, R.D.; Hayden, L.; Renninger, K.; Beyene, G.; Taylor, N.J.; Carrington, J.C.; Staskawicz, B.J.; et al. Simultaneous CRISPR/Cas9-mediated editing of cassava eIF4E isoforms nCBP-1 and nCBP-2 reduces cassava brown streak disease symptom severity and incidence. Plant Biotechnol. J. 2019, 17, 421–434. [Google Scholar] [CrossRef] [Green Version]
- Krishna, H.; Alizadeh, M.; Singh, D.; Singh, U.; Chauhan, N.; Eftekhari, M.; Sadh, R.K. Somaclonal variations and their applications in horticultural crops improvement. 3 Biotech 2016, 6, 54. [Google Scholar] [CrossRef] [Green Version]
- Søren, K.; Toni, W.; Christoph, D.; Hanne, C.T.; Magnus, R.; Morten Egevang, J.; Qiongxian, L.; Cynthia, V.; Emiko, M.; Jeppe Thulin, Ø.; et al. FIND-IT: Ultrafast mining of genome diversity. bioRxiv 2021. [Google Scholar] [CrossRef]
- Wagh, S.G.; Pohare, M.B. Current and Future Prospects of Plant Breeding with CRISPR/Cas. Current. J. Appl. Sci. Technol. 2019, 38, 1–17. [Google Scholar] [CrossRef]
- Zhu, L.H.J.; Holmes, B.R.; Aronin, N.; Brodsky, M.H. CRISPRseek: A Bioconductor Package to Identify Target-Specific Guide RNAs for CRISPR-Cas9 Genome-Editing Systems. PLoS ONE 2014, 9, e108424. [Google Scholar] [CrossRef] [Green Version]
- Jinek, M.; Jiang, F.G.; Taylor, D.W.; Sternberg, S.H.; Kaya, E.; Ma, E.B.; Anders, C.; Hauer, M.; Zhou, K.H.; Lin, S.; et al. Structures of Cas9 Endonucleases Reveal RNA-Mediated Conformational Activation. Science 2014, 343, 1247997. [Google Scholar] [CrossRef] [Green Version]
- Anders, C.; Niewoehner, O.; Duerst, A.; Jinek, M. Structural basis of PAM-dependent target DNA recognition by the Cas9 endonuclease. Nature 2014, 513, 569–573. [Google Scholar] [CrossRef] [Green Version]
- Gasiunas, G.; Siksnys, V. RNA-dependent DNA endonuclease Cas9 of the CRISPR system: Holy Grail of genome editing? Trends Microbiol. 2013, 21, 562–567. [Google Scholar] [CrossRef]
- Jiang, F.G.; Doudna, J.A. CRISPR-Cas9 Structures and Mechanisms. Annu. Rev. Biophys. 2017, 46, 505–529. [Google Scholar] [CrossRef] [Green Version]
- Al Abdallah, Q.; Ge, W.B.; Fortwendel, J.R. A Simple and Universal System for Gene Manipulation in Aspergillus fumigatus: In Vitro-Assembled Cas9-Guide RNA Ribonucleoproteins Coupled with Microhomology Repair Templates. Msphere 2017, 2, e00446-17. [Google Scholar] [CrossRef] [Green Version]
- Lemos, B.R.; Kaplan, A.C.; Bae, J.E.; Ferrazzoli, A.E.; Kuo, J.; Anand, R.P.; Waterman, D.P.; Haber, J.E. CRISPR/Cas9 cleavages in budding yeast reveal templated insertions and strand-specific insertion/deletion profiles. Proc. Natl. Acad. Sci. USA 2018, 115, E2040–E2047. [Google Scholar] [CrossRef] [Green Version]
- Song, F.; Stieger, K. Optimizing the DNA Donor Template for Homology-Directed Repair of Double-Strand Breaks. Mol. Ther. Nucleic Acids 2017, 7, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Cox, D.B.T.; Platt, R.J.; Zhang, F. Therapeutic genome editing: Prospects and challenges. Nat. Med. 2015, 21, 121–131. [Google Scholar] [CrossRef] [Green Version]
- Danner, E.; Bashir, S.; Yumlu, S.; Wurst, W.; Wefers, B.; Kuhn, R. Control of gene editing by manipulation of DNA repair mechanisms. Mamm. Genome 2017, 28, 262–274. [Google Scholar] [CrossRef] [Green Version]
- Boubakri, H. Recent progress in CRISPR/Cas9-based genome editing for enhancing plant disease resistance. Gene 2023, 866, 147334. [Google Scholar] [CrossRef]
- Wan, D.Y.; Guo, Y.; Cheng, Y.; Hu, Y.; Xiao, S.Y.; Wang, Y.J.; Wen, Y.Q. CRISPR/Cas9-mediated mutagenesis of VvMLO3 results in enhanced resistance to powdery mildew in grapevine (Vitis vinifera). Hortic. Res. 2020, 7, 116. [Google Scholar] [CrossRef]
- Atarashi, H.; Jayasinghe, W.H.; Kwon, J.; Kim, H.; Taninaka, Y.; Igarashi, M.; Ito, K.; Yamada, T.; Masuta, C.; Nakahara, K.S. Artificially Edited Alleles of the Eukaryotic Translation Initiation Factor 4E1 Gene Differentially Reduce Susceptibility to Cucumber Mosaic Virus and Potato Virus Y in Tomato. Front. Microbiol. 2020, 11, 564310. [Google Scholar] [CrossRef]
- Peng, A.; Chen, S.; Lei, T.; Xu, L.; He, Y.; Wu, L.; Yao, L.; Zou, X. Engineering canker-resistant plants through CRISPR/Cas9-targeted editing of the susceptibility gene CsLOB1 promoter in citrus. Plant Biotechnol. J. 2017, 15, 1509–1519. [Google Scholar] [CrossRef] [Green Version]
- Soyk, S.; Lemmon, Z.H.; Oved, M.; Fisher, J.; Liberatore, K.L.; Park, S.J.; Goren, A.; Jiang, K.; Ramos, A.; van der Knaap, E.; et al. Bypassing Negative Epistasis on Yield in Tomato Imposed by a Domestication Gene. Cell 2017, 169, 1142–1155.e12. [Google Scholar] [CrossRef]
- Zhang, B.; Xu, X.J.; Huang, R.W.; Yang, S.; Li, M.Y.; Guo, Y.L. CRISPR/Cas9-mediated targeted mutation reveals a role for AN4 rather than DPL in regulating venation formation in the corolla tube of Petunia hybrida. Hortic. Res. 2021, 8, 116. [Google Scholar] [CrossRef]
- Lopez-Casado, G.; Sanchez-Raya, C.; Ric-Varas, P.D.; Paniagua, C.; Blanco-Portales, R.; Munoz-Blanco, J.; Pose, S.; Matas, A.J.; Mercado, J.A. CRISPR/Cas9 editing of the polygalacturonase FaPG1 gene improves strawberry fruit firmness. Hortic. Res. 2023, 10, uhad011. [Google Scholar] [CrossRef]
- Li, X.D.; Wang, Y.N.; Chen, S.; Tian, H.Q.; Fu, D.Q.; Zhu, B.Z.; Luo, Y.B.; Zhu, H.L. Lycopene Is Enriched in Tomato Fruit by CRISPR/Cas9-Mediated Multiplex Genome Editing. Front. Plant Sci. 2018, 9, 559. [Google Scholar] [CrossRef]
- Beracochea, V.; Stritzler, M.; Radonic, L.; Bottero, E.; Jozefkowicz, C.; Darqui, F.; Ayub, N.; Bilbao, M.L.; Soto, G. CRISPR/Cas9-mediated knockout of SPL13 radically increases lettuce yield. Plant Cell Rep. 2023, 42, 645–647. [Google Scholar] [CrossRef]
- Nitarska, D.; Boehm, R.; Debener, T.; Lucaciu, R.C.; Halbwirth, H. First genome edited poinsettias: Targeted mutagenesis of flavonoid 3′-hydroxylase using CRISPR/Cas9 results in a colour shift. Plant Cell Tissue Organ Cult. 2021, 147, 49–60. [Google Scholar] [CrossRef]
- Huang, W.; Hu, N.; Xiao, Z.N.; Qiu, Y.P.; Yang, Y.; Yang, J.; Mao, X.; Wang, Y.C.; Li, Z.G.; Guo, H.W. A molecular framework of ethylene-mediated fruit growth and ripening processes in tomato. Plant Cell 2022, 34, 3280–3300. [Google Scholar] [CrossRef]
- Huynh, T.T.H.; Nguyen, T.L.; Luu, H.L.; Nguyen, H.H.; Le, H.D.; Bui, M.M.; Pham, T.H.; Doan, T.B.T.; Le, T.T.H.; Ha, H.H.; et al. Isolation and Characterization of a Dreb Homolog Gene from a Local Drought-Tolerant Maize Cultivar. Acta Biol. Cracoviensia Bot. 2019, 61, 13–24. [Google Scholar] [CrossRef]
- Wang, Y.P.; Wang, J.F.; Guo, S.G.; Tian, S.W.; Zhang, J.; Ren, Y.; Li, M.Y.; Gong, G.Y.; Zhang, H.Y.; Xu, Y. CRISPR/Cas9-mediated mutagenesis of ClBG1 decreased seed size and promoted seed germination in watermelon. Hortic. Res. 2021, 8, 70. [Google Scholar] [CrossRef]
- Kishchenko, O.; Zhou, Y.Z.; Jatayev, S.; Shavrukov, Y.; Borisjuk, N. Gene editing applications to modulate crop flowering time and seed dormancy. Abiotech 2020, 1, 233–245. [Google Scholar] [CrossRef]
- Lv, S.W.; Wu, W.G.; Wang, M.H.; Meyer, R.S.; Ndjiondjop, M.N.; Tan, L.B.; Zhou, H.Y.; Zhang, J.W.; Fu, Y.C.; Cai, H.W.; et al. Genetic control of seed shattering during African rice domestication. Nat. Plants 2018, 4, 331–337. [Google Scholar] [CrossRef]
- Li, T.; Xu, Y.X.; Zhang, L.C.; Ji, Y.L.; Tan, D.M.; Yuan, H.; Wang, A.D. The Jasmonate-Activated Transcription Factor MdMYC2 Regulates ETHYLENE RESPONSE FACTOR and Ethylene Biosynthetic Genes to Promote Ethylene Biosynthesis during Apple Fruit Ripening. Plant Cell 2017, 29, 1316–1334. [Google Scholar] [CrossRef] [Green Version]
- Jo, H.; Woo, C.; Norah, N.; Song, J.T.; Lee, J.D. Novel Allele of FAD2-1A from an EMS-Induced Mutant Soybean Line (PE529) Produces Elevated Levels of Oleic Acid in Soybean Oil. Agronomy 2022, 12, 2115. [Google Scholar] [CrossRef]
- Fu, M.X.; Chen, L.; Cai, Y.P.; Su, Q.; Chen, Y.Y.; Hou, W.S. CRISPR/Cas9-Mediated Mutagenesis of GmFAD2-1A and/or GmFAD2-1B to Create High-Oleic-Acid Soybean. Agronomy 2022, 12, 3218. [Google Scholar] [CrossRef]
- Mellidou, I.; Koukounaras, A.; Kostas, S.; Patelou, E.; Kanellis, A.K. Regulation of Vitamin C Accumulation for Improved Tomato Fruit Quality and Alleviation of Abiotic Stress. Genes 2021, 12, 694. [Google Scholar] [CrossRef]
- Liu, J.H.; Liu, M.T.; Wang, J.Y.; Zhang, J.; Miao, H.X.; Wang, Z.; Jia, C.H.; Zhang, J.B.; Xu, B.Y.; Jin, Z.Q. Transcription factor MaMADS36 plays a central role in regulating banana fruit ripening. J. Exp. Bot. 2021, 72, 7078–7091. [Google Scholar] [CrossRef]
- Manghwar, H.; Li, B.; Ding, X.; Hussain, A.; Lindsey, K.; Zhang, X.L.; Jin, S.X. CRISPR/Cas Systems in Genome Editing: Methodologies and Tools for sgRNA Design, Off-Target Evaluation, and Strategies to Mitigate Off-Target Effects. Adv. Sci. 2020, 7, 1902312. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Wolt, J.D. Risk associated with off-target plant genome editing and methods for its limitation. Emerg. Top. Life Sci. 2017, 1, 231–240. [Google Scholar] [CrossRef] [Green Version]
- Peng, R.X.; Lin, G.G.; Li, J.M. Potential pitfalls of CRISPR/Cas9-mediated genome editing. Febs. J. 2016, 283, 1218–1231. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Bae, S.; Park, J.; Kim, E.; Kim, S.; Yu, H.R.; Hwang, J.; Kim, J.I.; Kim, J.S. Digenome-seq: Genome-wide profiling of CRISPR-Cas9 off-target effects in human cells. Nat. Methods 2015, 12, 237–243. [Google Scholar] [CrossRef]
- Vakulskas, C.A.; Dever, D.P.; Rettig, G.R.; Turk, R.; Jacobi, A.M.; Collingwood, M.A.; Bode, N.M.; McNeill, M.S.; Yan, S.Q.; Camarena, J.; et al. A high-fidelity Cas9 mutant delivered as a ribonucleoprotein complex enables efficient gene editing in human hematopoietic stem and progenitor cells. Nat. Med. 2018, 24, 1216–1224. [Google Scholar] [CrossRef] [Green Version]
- Anjanappa, R.B.; Gruissem, W. Current progress and challenges in crop genetic transformation. J. Plant Physiol. 2021, 261, 153411. [Google Scholar] [CrossRef]
- Gonzalez, M.N.; Massa, G.A.; Andersson, M.; Oneto, C.A.D.; Turesson, H.; Storani, L.; Olsson, N.; Falt, A.S.; Hofvander, P.; Feingold, S.E. Comparative potato genome editing: Agrobacterium tumefaciens-mediated transformation and protoplasts transfection delivery of CRISPR/Cas9 components directed to StPPO2 gene. Plant Cell Tissue Organ Cult. 2021, 145, 291–305. [Google Scholar] [CrossRef]
- Zlobin, N.E.; Lebedeva, M.V.; Taranov, V.V. CRISPR/Cas9 genome editing through in planta transformation. Crit. Rev. Biotechnol. 2020, 40, 153–168. [Google Scholar] [CrossRef]
- Bhowmik, P.; Konkin, D.; Polowick, P.; Hodgins, C.L.; Subedi, M.; Xiang, D.; Yu, B.; Patterson, N.; Rajagopalan, N.; Babic, V.; et al. CRISPR/Cas9 gene editing in legume crops: Opportunities and challenges. Legume Sci. 2021, 3, e96. [Google Scholar] [CrossRef]
- Cardi, T.; D’Agostino, N.; Tripodi, P. Genetic Transformation and Genomic Resources for Next-Generation Precise Genome Engineering in Vegetable Crops. Front. Plant Sci. 2017, 8, 241. [Google Scholar] [CrossRef] [Green Version]
- Kausch, A.P.; Nelson-Vasilchik, K.; Hague, J.; Mookkan, M.; Quemada, H.; Dellaporta, S.; Fragoso, C.; Zhang, Z.Y.J. Edit at will: Genotype independent plant transformation in the era of advanced genomics and genome editing. Plant Sci. 2019, 281, 186–205. [Google Scholar] [CrossRef]
- Lee, K.S.; Wang, K. Strategies for genotype-flexible plant transformation. Curr. Opin. Biotechnol. 2023, 79, 102848. [Google Scholar] [CrossRef]
- Son, S.; Park, S.R. Challenges Facing CRISPR/Cas9-Based Genome Editing in Plants. Front. Plant Sci. 2022, 13, 902413. [Google Scholar] [CrossRef]
- Jackson, M.A.; Anderson, D.J.; Birch, R.G. Comparison of Agrobacterium and particle bombardment using whole plasmid or minimal cassette for production of high-expressing, low-copy transgenic plants. Transgenic Res. 2013, 22, 143–151. [Google Scholar] [CrossRef]
- Vats, S.; Kumawat, S.; Kumar, V.; Patil, G.B.; Joshi, T.; Sonah, H.; Sharma, T.R.; Deshmukh, R. Genome Editing in Plants: Exploration of Technological Advancements and Challenges. Cells 2019, 8, 1386. [Google Scholar] [CrossRef] [Green Version]
- Basak, J.; Nithin, C. Targeting Non-Coding RNAs in Plants with the CRISPR-Cas Technology is a Challenge yet Worth Accepting. Front. Plant Sci. 2015, 6, 1001. [Google Scholar] [CrossRef] [Green Version]
- Qi, L.S.; Larson, M.H.; Gilbert, L.A.; Doudna, J.A.; Weissman, J.S.; Arkin, A.P.; Lim, W.A. Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression. Cell 2013, 152, 1173–1183. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.J.; Lim, D.A. Modulating the expression of long non-coding RNAs for functional studies. EMBO Rep. 2018, 19, e46955. [Google Scholar] [CrossRef]
- Tycko, J.; Wainberg, M.; Marinov, G.K.; Ursu, O.; Hess, G.T.; Ego, B.K.; Aradhana; Li, A.; Truong, A.; Trevino, A.E.; et al. Mitigation of off-target toxicity in CRISPR-Cas9 screens for essential non-coding elements. Nat. Commun. 2019, 10, 4063. [Google Scholar] [CrossRef] [Green Version]
- Zischewski, J.; Fischer, R.; Bortesi, L. Detection of on-target and off-target mutations generated by CRISPR/Cas9 and other sequence-specific nucleases. Biotechnol. Adv. 2017, 35, 95–104. [Google Scholar] [CrossRef]
- Hoijer, I.; Johansson, J.; Gudmundsson, S.; Chin, C.S.; Bunikis, I.; Haggqvist, S.; Emmanouilidou, A.; Wilbe, M.; den Hoed, M.; Bondeson, M.L.; et al. Amplification-free long-read sequencing reveals unforeseen CRISPR-Cas9 off-target activity. Genome Biol. 2020, 21, 290. [Google Scholar] [CrossRef]
- Teotia, S.; Singh, D.; Tang, X.Q.; Tang, G.L. Essential RNA-Based Technologies and Their Applications in Plant Functional Genomics. Trends Biotechnol. 2016, 34, 106–123. [Google Scholar] [CrossRef]
- Durr, J.; Papareddy, R.; Nakajima, K.; Gutierrez-Marcos, J. Highly efficient heritable targeted deletions of gene clusters and non-coding regulatory regions in Arabidopsis using CRISPR/Cas9. Sci. Rep. 2018, 8, 4443. [Google Scholar] [CrossRef]
- Waheed, S.; Zeng, L.H. The Critical Role of miRNAs in Regulation of Flowering Time and Flower Development. Genes 2020, 11, 319. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.Y.; Shin, C. Regulatory non-coding RNAs in plants: Potential gene resources for the improvement of agricultural traits. Plant Biotechnol. Rep. 2016, 10, 35–47. [Google Scholar] [CrossRef]
- Han, H.A.; Pang, J.K.S.; Soh, B.S. Mitigating off-target effects in CRISPR/Cas9-mediated in vivo gene editing. J. Mol. Med. 2020, 98, 615–632. [Google Scholar] [CrossRef] [Green Version]
- Salika, R.; Riffat, J. Abiotic stress responses in maize: A review. Acta Physiol. Plant. 2021, 43, 130. [Google Scholar] [CrossRef]
- Mao, Y.F.; Botella, J.R.; Liu, Y.G.; Zhu, J.K. Gene editing in plants: Progress and challenges. Natl. Sci. Rev. 2019, 6, 421–437. [Google Scholar] [CrossRef]
- Xu, R.F.; Li, H.; Qin, R.Y.; Li, J.; Qiu, C.H.; Yang, Y.C.; Ma, H.; Li, L.; Wei, P.C.; Yang, J.B. Generation of inheritable and “transgene clean” targeted genome-modified rice in later generations using the CRISPR/Cas9 system. Sci. Rep. 2015, 5, 11491. [Google Scholar] [CrossRef] [Green Version]
- Cermak, T.; Baltes, N.J.; Cegan, R.; Zhang, Y.; Voytas, D.F. High-frequency, precise modification of the tomato genome. Genome Biol. 2015, 16, 232. [Google Scholar] [CrossRef] [Green Version]
- Zhu, H.C.; Li, C.; Gao, C.X. Applications of CRISPR-Cas in agriculture and plant biotechnology. Nat. Rev. Mol. Cell Biol. 2020, 21, 782. [Google Scholar] [CrossRef]
- Mahfouz, M.M.; Piatek, A.; Stewart, C.N. Genome engineering via TALENs and CRISPR/Cas9 systems: Challenges and perspectives. Plant Biotechnol. J. 2014, 12, 1006–1014. [Google Scholar] [CrossRef]
- Eki, R.; She, J.; Parlak, M.; Benamar, M.; Du, K.P.; Kumar, P.; Abbas, T. A robust CRISPR-Cas9-based fluorescent reporter assay for the detection and quantification of DNA double-strand break repair. Nucleic Acids Res. 2020, 48, e126. [Google Scholar] [CrossRef]
- Bradford, K.J.; Van Deynze, A.; Gutterson, N.; Parrott, W.; Strauss, S.H. Regulating transgenic crops sensibly: Lessons from plant breeding, biotechnology and genomics. Nat. Biotechnol. 2005, 23, 439–444. [Google Scholar] [CrossRef]
- Alonge, M.; Wang, X.G.; Benoit, M.; Soyk, S.; Pereira, L.; Zhang, L.; Suresh, H.; Ramakrishnan, S.; Maumus, F.; Ciren, D.; et al. Major Impacts of Widespread Structural Variation on Gene Expression and Crop Improvement in Tomato. Cell 2020, 182, 145–161. [Google Scholar] [CrossRef]
- Zhou, H.B.; Liu, B.; Weeks, D.P.; Spalding, M.H.; Yang, B. Large chromosomal deletions and heritable small genetic changes induced by CRISPR/Cas9 in rice. Nucleic Acids Res. 2014, 42, 10903–10914. [Google Scholar] [CrossRef]
- Lorenzo, C.D.; Debray, K.; Herwegh, D.; Develtere, W.; Impens, L.; Schaumont, D.; Vandeputte, W.; Aesaert, S.; Coussens, G.; De Boe, Y.; et al. BREEDIT: A multiplex genome editing strategy to improve complex quantitative traits in maize. Plant Cell 2023, 35, 1160. [Google Scholar] [CrossRef]
- Modrzejewski, D.; Hartung, F.; Sprink, T.; Krause, D.; Kohl, C.; Schiemann, J.; Wilhelm, R. What is the available evidence for the application of genome editing as a new tool for plant trait modification and the potential occurrence of associated off-target effects: A systematic map protocol. Environ. Evid. 2018, 7, 18. [Google Scholar] [CrossRef]
- Zhang, D.; Zhang, H.; Li, T.; Chen, K.; Qiu, J.-L.; Gao, C. Perfectly matched 20-nucleotide guide RNA sequences enable robust genome editing using high-fidelity SpCas9 nucleases. Genome Biol. 2017, 18, 191. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.; Shang, P.; Mohanraju, P.; Geijsen, N. Prime editing: Advances and therapeutic applications. Trends Biotechnol. 2023, 41, 1000–1012. [Google Scholar] [CrossRef] [PubMed]
- Rasheed, A.; Gill, R.A.; Hassan, M.U.; Mahmood, A.; Qari, S.; Zaman, Q.U.; Ilyas, M.; Aamer, M.; Batool, M.; Li, H. A critical review: Recent advancements in the use of CRISPR/Cas9 technology to enhance crops and alleviate global food crises. Curr. Issues Mol. Biol. 2021, 43, 1950–1976. [Google Scholar] [CrossRef] [PubMed]
- Wada, N.; Osakabe, K.; Osakabe, Y. Expanding the plant genome editing toolbox with recently developed CRISPR–Cas systems. Plant Physiol. 2022, 188, 1825–1837. [Google Scholar] [CrossRef] [PubMed]
- Slesarenko, Y.S.; Lavrov, A.V.; Smirnikhina, S.A. Off-target effects of base editors: What we know and how we can reduce it. Curr. Genet. 2022, 68, 39–48. [Google Scholar] [CrossRef] [PubMed]
- Ghogare, R.; Ludwig, Y.; Bueno, G.M.; Slamet-Loedin, I.H.; Dhingra, A. Genome editing reagent delivery in plants. Transgenic Res. 2021, 30, 321–335. [Google Scholar] [CrossRef]
- Wijerathna-Yapa, A.; Ramtekey, V.; Ranawaka, B.; Basnet, B.R. Applications of in vitro tissue culture technologies in breeding and genetic improvement of wheat. Plants 2022, 11, 2273. [Google Scholar] [CrossRef]
- Lee, J.; Chung, J.H.; Kim, H.M.; Kim, D.W.; Kim, H. Designed nucleases for targeted genome editing. Plant Biotechnol. J. 2016, 14, 448–462. [Google Scholar] [CrossRef]
- Sanjana, N.E.; Cong, L.; Zhou, Y.; Cunniff, M.M.; Feng, G.P.; Zhang, F. A transcription activator-like effector toolbox for genome engineering. Nat. Protoc. 2012, 7, 171–192. [Google Scholar] [CrossRef] [Green Version]
- Morbitzer, R.; Romer, P.; Boch, J.; Lahaye, T. Regulation of selected genome loci using de novo-engineered transcription activator-like effector (TALE)-type transcription factors. Proc. Natl. Acad. Sci. USA 2010, 107, 21617–21622. [Google Scholar] [CrossRef]
- Miller, J.C.; Tan, S.Y.; Qiao, G.J.; Barlow, K.A.; Wang, J.B.; Xia, D.F.; Meng, X.D.; Paschon, D.E.; Leung, E.; Hinkley, S.J.; et al. A TALE nuclease architecture for efficient genome editing. Nat. Biotechnol. 2011, 29, 143–148. [Google Scholar] [CrossRef]
- Christian, M.L.; Demorest, Z.L.; Starker, C.G.; Osborn, M.J.; Nyquist, M.D.; Zhang, Y.; Carlson, D.F.; Bradley, P.; Bogdanove, A.J.; Voytas, D.F. Targeting G with TAL Effectors: A Comparison of Activities of TALENs Constructed with NN and NK Repeat Variable Di-Residues. PLoS ONE 2012, 7, e45383. [Google Scholar] [CrossRef] [Green Version]
- Xiao, A.; Wu, Y.D.; Yang, Z.P.; Hu, Y.Y.; Wang, W.Y.; Zhang, Y.T.; Kong, L.; Gao, G.; Zhu, Z.Y.; Lin, S.; et al. EENdb: A database and knowledge base of ZFNs and TALENs for endonuclease engineering. Nucleic Acids Res. 2013, 41, D415–D422. [Google Scholar] [CrossRef] [Green Version]
- Cermak, T.; Doyle, E.L.; Christian, M.; Wang, L.; Zhang, Y.; Schmidt, C.; Baller, J.A.; Somia, N.V.; Bogdanove, A.J.; Voytas, D.F. Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting. Nucleic Acids Res. 2011, 39, 7879. [Google Scholar] [CrossRef] [Green Version]
- Puchta, H.; Fauser, F. Synthetic nucleases for genome engineering in plants: Prospects for a bright future. Plant J. 2014, 78, 727–741. [Google Scholar] [CrossRef] [PubMed]
- Mackon, E.; Jeazet Dongho Epse Mackon, G.C.; Guo, Y.; Ma, Y.; Yao, Y.; Liu, P. Development and application of CRISPR/Cas9 to improve anthocyanin pigmentation in plants: Opportunities and perspectives. Plant Sci. 2023, 333, 111746. [Google Scholar] [CrossRef] [PubMed]
- Jia, H.; Orbovic, V.; Jones, J.B.; Wang, N. Modification of the PthA4 effector binding elements in Type I CsLOB1 promoter using Cas9/sgRNA to produce transgenic Duncan grapefruit alleviating XccΔpthA4:dCsLOB1.3 infection. Plant Biotechnol. J. 2016, 14, 1291–1301. [Google Scholar] [CrossRef]
- Jaganathan, D.; Ramasamy, K.; Sellamuthu, G.; Jayabalan, S.; Venkataraman, G. CRISPR for Crop Improvement: An Update Review. Front. Plant Sci. 2018, 9, 985. [Google Scholar] [CrossRef]
- Holkers, M.; Maggio, I.; Henriques, S.F.D.; Janssen, J.M.; Cathomen, T.; Goncalves, M.A.F.V. Adenoviral vector DNA for accurate genome editing with engineered nucleases. Nat. Methods 2014, 11, 1051–1057. [Google Scholar] [CrossRef]
- Tiwari, J.K.; Challam, C.; Chakrabarti, S.K.; Feingold, S.E. Climate-Smart Potato: An Integrated Breeding, Genomics, and Phenomics Approach. In Genomic Designing of Climate-Smart Vegetable Crops; Kole, C., Ed.; Springer International Publishing: Cham, Switzerland, 2020; pp. 1–46. [Google Scholar]
- Wei, C.; Liu, J.; Yu, Z.; Zhang, B.; Gao, G.; Jiao, R. TALEN or Cas9—Rapid, efficient and specific choices for genome modifications. J. Genet. Genom. 2013, 40, 281–289. [Google Scholar] [CrossRef]
- Doyle, E.L.; Stoddard, B.L.; Voytas, D.F.; Bogdanove, A.J. TAL effectors: Highly adaptable phytobacterial virulence factors and readily engineered DNA-targeting proteins. Trends Cell Biol. 2013, 23, 390–398. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.M.; Cradick, T.J.; Fine, E.J.; Bao, G. Nuclease Target Site Selection for Maximizing On-target Activity and Minimizing Off-target Effects in Genome Editing. Mol. Ther. 2016, 24, 475–487. [Google Scholar] [CrossRef] [Green Version]
- Wright, D.A.; Li, T.; Yang, B.; Spalding, M.H. TALEN-mediated genome editing: Prospects and perspectives. Biochem. J. 2014, 462, 15–24. [Google Scholar] [CrossRef]
- Li, H.Y.; Yang, Y.; Hong, W.Q.; Huang, M.Y.; Wu, M.; Zhao, X. Applications of genome editing technology in the targeted therapy of human diseases: Mechanisms, advances and prospects. Signal Transduct. Target. Ther. 2020, 5, 1. [Google Scholar] [CrossRef]
- Castro, N.G.; Bjelic, J.; Malhotra, G.; Huang, C.; Alsaffar, S.H. Comparison of the Feasibility, Efficiency, and Safety of Genome Editing Technologies. Int. J. Mol. Sci. 2021, 22, 10355. [Google Scholar] [CrossRef]
- Zhu, C.F.; Bortesi, L.; Baysal, C.; Twyman, R.M.; Fischer, R.; Capell, T.; Schillberg, S.; Christou, P. Characteristics of Genome Editing Mutations in Cereal Crops. Trends Plant Sci. 2017, 22, 38–52. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhang, F.; Li, X.H.; Baller, J.A.; Qi, Y.P.; Starker, C.G.; Bogdanove, A.J.; Voytas, D.F. Transcription Activator-Like Effector Nucleases Enable Efficient Plant Genome Engineering. Plant Physiol. 2013, 161, 20–27. [Google Scholar] [CrossRef] [Green Version]
- Malzahn, A.; Lowder, L.; Qi, Y.P. Plant genome editing with TALEN and CRISPR. Cell Biosci. 2017, 7, 21. [Google Scholar] [CrossRef] [Green Version]
- Ansari, W.A.; Chandanshive, S.U.; Bhatt, V.; Nadaf, A.B.; Vats, S.; Katara, J.L.; Sonah, H.; Deshmukh, R. Genome Editing in Cereals: Approaches, Applications and Challenges. Int. J. Mol. Sci. 2020, 21, 4040. [Google Scholar] [CrossRef]
- Voytas, D.F.; Gao, C.X. Precision Genome Engineering and Agriculture: Opportunities and Regulatory Challenges. PLoS Biol. 2014, 12, e1001877. [Google Scholar] [CrossRef]
- Yau, Y.Y.; Stewart, C.N. Less is more: Strategies to remove marker genes from transgenic plants. BMC Biotechnol. 2013, 13, 36. [Google Scholar] [CrossRef] [Green Version]
- Yee, J.K. Off-target effects of engineered nucleases. FEBS J. 2016, 283, 3239–3248. [Google Scholar] [CrossRef] [Green Version]
- Gurushidze, M.; Hensel, G.; Hiekel, S.; Schedel, S.; Valkov, V.; Kumlehn, J. True-Breeding Targeted Gene Knock-Out in Barley Using Designer TALE-Nuclease in Haploid Cells. PLoS ONE 2014, 9, e92046. [Google Scholar] [CrossRef] [Green Version]
- Kopischke, S.; Schussler, E.; Althoff, F.; Zachgo, S. TALEN-mediated genome-editing approaches in the liverwort Marchantia polymorpha yield high efficiencies for targeted mutagenesis. Plant Methods 2017, 13, 20. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Prasad, M. Advances in Agrobacterium tumefaciens-mediated genetic transformation of graminaceous crops. Protoplasma 2016, 253, 691–707. [Google Scholar] [CrossRef]
- Čermák, T.; Curtin, S.J.; Gil-Humanes, J.; Čegan, R.; Kono, T.J.; Konečná, E.; Belanto, J.J.; Starker, C.G.; Mathre, J.W.; Greenstein, R.L. A multipurpose toolkit to enable advanced genome engineering in plants. Plant Cell 2017, 29, 1196–1217. [Google Scholar]
- Yan, Y.; Zhu, X.; Yu, Y.; Li, C.; Zhang, Z.; Wang, F. Nanotechnology strategies for plant genetic engineering. Adv. Mater. 2022, 34, 2106945. [Google Scholar]
- Govindan, G.; Ramalingam, S. Programmable Site-Specific Nucleases for Targeted Genome Engineering in Higher Eukaryotes. J. Cell Physiol. 2016, 231, 2380–2392. [Google Scholar] [CrossRef]
- Durai, S.; Mani, M.; Kandavelou, K.; Wu, J.; Porteus, M.H.; Chandrasegaran, S. Zinc finger nucleases: Custom-designed molecular scissors for genome engineering of plant and mammalian cells. Nucleic Acids Res. 2005, 33, 5978–5990. [Google Scholar] [CrossRef]
- Mandell, J.G.; Barbas, C.F. Zinc finger tools: Custom DNA-binding domains for transcription factors and nucleases. Nucleic Acids Res. 2006, 34, W516–W523. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.M.T. Multiple modes of RNA recognition by zinc finger proteins. Curr. Opin. Struct. Biol. 2005, 15, 367–373. [Google Scholar] [CrossRef] [Green Version]
- Cathomen, T.; Joung, J.K. Zinc-finger nucleases: The next generation emerges. Mol. Ther. 2008, 16, 1200–1207. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, J.S. A guide to genome engineering with programmable nucleases. Nat. Rev. Genet. 2014, 15, 321–334. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, A.; Plaisier, C.L.; Carroll, D.; Drews, G.N. Targeted mutagenesis using zinc-finger nucleases in Arabidopsis. Proc. Natl. Acad. Sci. USA 2005, 102, 2232–2237. [Google Scholar] [CrossRef]
- Maeder, M.L.; Thibodeau-Beganny, S.; Osiak, A.; Wright, D.A.; Anthony, R.M.; Eichtinger, M.; Jiang, T.; Foley, J.E.; Winfrey, R.J.; Townsend, J.A.; et al. Rapid “Open-Source” engineering of customized zinc-finger nucleases for highly efficient gene modification. Mol. Cell 2008, 31, 294–301. [Google Scholar] [CrossRef] [Green Version]
- Marton, I.; Zuker, A.; Shklarman, E.; Zeevi, V.; Tovkach, A.; Roffe, S.; Ovadis, M.; Tzfira, T.; Vainstein, A. Nontransgenic Genome Modification in Plant Cells. Plant Physiol. 2010, 154, 1079–1087. [Google Scholar] [CrossRef] [Green Version]
- Shukla, V.K.; Doyon, Y.; Miller, J.C.; DeKelver, R.C.; Moehle, E.A.; Worden, S.E.; Mitchell, J.C.; Arnold, N.L.; Gopalan, S.; Meng, X.D.; et al. Precise genome modification in the crop species Zea mays using zinc-finger nucleases. Nature 2009, 459, 437–441. [Google Scholar] [CrossRef] [PubMed]
- Townsend, J.A.; Wright, D.A.; Winfrey, R.J.; Fu, F.L.; Maeder, M.L.; Joung, J.K.; Voytas, D.F. High-frequency modification of plant genes using engineered zinc-finger nucleases. Nature. 2009, 459, 442–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tovkach, A.; Zeevi, V.; Tzfira, T. A toolbox and procedural notes for characterizing novel zinc finger nucleases for genome editing in plant cells. Plant J. 2009, 57, 747–757. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Maeder, M.L.; Unger-Wallace, E.; Hoshaw, J.P.; Reyon, D.; Christian, M.; Li, X.H.; Pierick, C.J.; Dobbs, D.; Peterson, T.; et al. High frequency targeted mutagenesis in Arabidopsis thaliana using zinc finger nucleases. Proc. Natl. Acad. Sci. USA 2010, 107, 12028–12033. [Google Scholar] [CrossRef]
- De Pater, S.; Neuteboom, L.W.; Pinas, J.E.; Hooykaas, P.J.; van der Zaal, B.J. ZFN-induced mutagenesis and gene-targeting in Arabidopsis through Agrobacterium-mediated floral dip transformation. Plant Biotechnol. J. 2009, 7, 821–835. [Google Scholar] [CrossRef]
- Wu, J.; Kandavelou, K.; Chandrasegaran, S. Custom-designed zinc finger nucleases: What is next? Cell. Mol. Life Sci. 2007, 64, 2933–2944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhang, J.S.; Lang, Z.B.; Botella, J.R.; Zhu, J.K. Genome Editing-Principles and Applications for Functional Genomics Research and Crop Improvement. Crit. Rev. Plant Sci. 2017, 36, 291–309. [Google Scholar] [CrossRef]
- Liu, M.J.; Zhao, J.; Cai, Q.L.; Liu, G.C.; Wang, J.R.; Zhao, Z.H.; Liu, P.; Dai, L.; Yan, G.J.; Wang, W.J.; et al. The complex jujube genome provides insights into fruit tree biology. Nat. Commun. 2014, 5, 5315. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.H.; Tee, L.Y.; Wang, X.G.; Huang, Q.S.; Yang, S.H. Off-target Effects in CRISPR/Cas9-mediated Genome Engineering. Mol. Ther. Nucleic Acids 2015, 4, e264. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.M.; Yao, Y.F.; Zhang, Y.C.; Fan, G.F. CRISPR system: Discovery, development and off-target detection. Cell. Signal. 2020, 70, 109577. [Google Scholar] [CrossRef]
- Tzfira, T.; Weinthal, D.; Marton, I.; Zeevi, V.; Zuker, A.; Vainstein, A. Genome modifications in plant cells by custom-made restriction enzymes. Plant Biotechnol. J. 2012, 10, 373–389. [Google Scholar] [CrossRef] [PubMed]
- Welker, C.M.; Balasubramanian, V.K.; Petti, C.; Rai, K.M.; DeBolt, S.; Mendu, V. Engineering Plant Biomass Lignin Content and Composition for Biofuels and Bioproducts. Energies 2015, 8, 7654–7676. [Google Scholar] [CrossRef] [Green Version]
- Zaidi, S.S.E.A.; Mansoor, S. Viral Vectors for Plant Genome Engineering. Front. Plant Sci. 2017, 8, 539. [Google Scholar] [CrossRef] [Green Version]
- Chandrasegaran, S.; Carroll, D. Origins of programmable nucleases for genome engineering. J. Mol. Biol. 2016, 428, 963–989. [Google Scholar]
- Qi, Y. High efficient genome modification by designed zinc finger nuclease. In Advances in New Technology for Targeted Modification of Plant Genomes; Springer: New York, NY, USA, 2015; pp. 39–53. [Google Scholar]
- Osborn, M.J.; DeFeo, A.P.; Blazar, B.R.; Tolar, J. Synthetic zinc finger nuclease design and rapid assembly. Hum. Gene Ther. 2011, 22, 1155–1165. [Google Scholar]
- Yasafova, D.; Gankin, D.; Sharapov, Y.; Bright, C.; Driss, F.; Campi, F. panCRISPR Toolbox—A Deep Learning Approach to Improve CRISPR/Cas Experiments. 2022. Available online: https://www.mdsi.tum.de/fileadmin/w00cet/di-lab/pdf/Helmholtz_AI_WS2021_Final_Report.pdf (accessed on 22 February 2022).
- Matchett-Oates, L. Production of Novel Medicinal Cannabis Using Genome Editing Technology. Ph.D. Thesis, La Trobe University, Bundoora, Australia, 2022. [Google Scholar]
- Gao, C. Genome engineering for crop improvement and future agriculture. Cell 2021, 184, 1621–1635. [Google Scholar] [CrossRef]
- Wolt, J.D.; Wang, K.; Yang, B. The regulatory status of genome-edited crops. Plant Biotechnol. J. 2016, 14, 510–518. [Google Scholar] [CrossRef] [Green Version]
- Menz, J.; Modrzejewski, D.; Hartung, F.; Wilhelm, R.; Sprink, T. Genome edited crops touch the market: A view on the global development and regulatory environment. Front. Plant Sci. 2020, 11, 586027. [Google Scholar] [CrossRef]
- Bridgers, M. Genetically modified organisms and the precautionary principle: How the GMO dispute before the world trade organization could decide the fate of international GMO regulation. Temp. Envtl. L. Tech. J. 2003, 22, 171. [Google Scholar]
- Hundleby, P.; Harwood, W. Regulatory Constraints and Differences of Genome-Edited Crops Around the Globe. In Genome Editing: Current Technology Advances and Applications for Crop Improvement; Springer: Berlin/Heidelberg, Germany, 2022; pp. 319–341. [Google Scholar]
- Idris, S.H.; Mat Jalaluddin, N.S.; Chang, L.W. Ethical and legal implications of gene editing in plant breeding: A systematic literature review. J. Zhejiang Univ. Sci. B 2023, 1–13. [Google Scholar] [CrossRef]
- Wolt, J.D.; Wolf, C. Policy and governance perspectives for regulation of genome edited crops in the United States. Front. Plant Sci. 2018, 9, 1606. [Google Scholar] [CrossRef] [Green Version]
- Hartung, F.; Schiemann, J. Precise plant breeding using new genome editing techniques: Opportunities, safety and regulation in the EU. Plant J. 2014, 78, 742–752. [Google Scholar] [CrossRef]
- Eckerstorfer, M.F.; Engelhard, M.; Heissenberger, A.; Simon, S.; Teichmann, H. Plants developed by new genetic modification techniques—Comparison of existing regulatory frameworks in the EU and non-EU countries. Front. Bioeng. Biotechnol. 2019, 7, 26. [Google Scholar] [CrossRef] [Green Version]
- Lassoued, R.; Phillips, P.W.; Macall, D.M.; Hesseln, H.; Smyth, S.J. Expert opinions on the regulation of plant genome editing. Plant Biotechnol. J. 2021, 19, 1104–1109. [Google Scholar] [CrossRef]
- Turnbull, C.; Lillemo, M.; Hvoslef-Eide, T.A. Global regulation of genetically modified crops amid the gene edited crop boom—A review. Front. Plant Sci. 2021, 12, 630396. [Google Scholar] [CrossRef]
- Levidow, L.; Carr, S. GM Food on Trial: Testing European Democracy; Routledge: Oxfordshire, UK, 2009. [Google Scholar]
- Dederer, H.-G. Options for the Regulation of Genome Edited Plants–Framing the Issues. In Genome Editing in Agriculture; Nomos: Baden, Germany, 2019; pp. 75–122. [Google Scholar]
- Kawall, K.; Cotter, J.; Then, C. Broadening the GMO risk assessment in the EU for genome editing technologies in agriculture. Environ. Sci. Eur. 2020, 32, 106. [Google Scholar] [CrossRef]
- Entine, J.; Felipe, M.S.S.; Groenewald, J.-H.; Kershen, D.L.; Lema, M.; McHughen, A.; Nepomuceno, A.L.; Ohsawa, R.; Ordonio, R.L.; Parrott, W.A. Regulatory approaches for genome edited agricultural plants in select countries and jurisdictions around the world. Transgenic Res. 2021, 30, 551–584. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Araki, M. Consumer acceptance of food crops developed by genome editing. Plant Cell Rep. 2016, 35, 1507–1518. [Google Scholar] [CrossRef] [PubMed]
- Bartkowski, B.; Theesfeld, I.; Pirscher, F.; Timaeus, J. Snipping around for food: Economic, ethical and policy implications of CRISPR/Cas genome editing. Geoforum 2018, 96, 172–180. [Google Scholar]
- Bäckstrand, K. Civic science for sustainability: Reframing the role of experts, policy-makers and citizens in environmental governance. Glob. Environ. Politics 2003, 3, 24–41. [Google Scholar] [CrossRef]
- Sendhil, R.; Nyika, J.; Yadav, S.; Mackolil, J.; Prashat, G.; Workie, E.; Ragupathy, R.; Ragupathy, R.; Ramasundaram, P. Consumer Perception and Preference towards Genetically Modified (GM) Foods: Bibliometric Evidence and Policy Imperatives. agriRxiv 2021, 2021, 20210260196. [Google Scholar] [CrossRef]
- National Academies of Sciences, Engineering, and Medicine. The Current Biotechnology Regulatory System. In Preparing for Future Products of Biotechnology; National Academies Press (US): Washington, DC, USA, 2017. [Google Scholar]
- Ahmad, A.; Munawar, N.; Khan, Z.; Qusmani, A.T.; Khan, S.H.; Jamil, A.; Ashraf, S.; Ghouri, M.Z.; Aslam, S.; Mubarik, M.S. An outlook on global regulatory landscape for genome-edited crops. Int. J. Mol. Sci. 2021, 22, 11753. [Google Scholar]
- Hilder, V.A.; Boulter, D. Genetic engineering of crop plants for insect resistance—A critical review. Crop Prot. 1999, 18, 177–191. [Google Scholar] [CrossRef]
- Van Esse, H.P.; Reuber, T.L.; van der Does, D. Genetic modification to improve disease resistance in crops. New Phytol. 2020, 225, 70–86. [Google Scholar] [CrossRef] [Green Version]
- Malhotra, S.K. Horticultural crops and climate change: A review. Indian J. Agr. Sci. 2017, 87, 12–22. [Google Scholar] [CrossRef]
- Reguera, M.; Peleg, Z.; Blumwald, E. Targeting, metabolic pathways for genetic engineering abiotic stress-tolerance in crops. BBA-Gene Regul. Mech. 2012, 1819, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Zsogon, A.; Peres, L.E.P.; Xiao, Y.J.; Yan, J.B.; Fernie, A.R. Enhancing crop diversity for food security in the face of climate uncertainty. Plant J. 2022, 109, 402–414. [Google Scholar] [CrossRef] [PubMed]
- DellaPenna, D. Nutritional genomics: Manipulating plant micronutrients to improve human health. Science 1999, 285, 375–379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Palve, A.; Joshi, C.; Srivastava, R.K. Crop biofortification for iron (Fe), zinc (Zn) and vitamin A with transgenic approaches. Heliyon 2019, 5, e01914. [Google Scholar] [CrossRef] [Green Version]
- Lemmon, Z.H.; Reem, N.T.; Dalrymple, J.; Soyk, S.; Swartwood, K.E.; Rodriguez-Leal, D.; Van Eck, J.; Lippman, Z.B. Rapid improvement of domestication traits in an orphan crop by genome editing. Nat. Plants 2018, 4, 766–770. [Google Scholar] [CrossRef]
- Wang, T.; Zhang, H.Y.; Zhu, H.L. CRISPR technology is revolutionizing the improvement of tomato and other fruit crops. Hortic. Res. 2019, 6, 77. [Google Scholar] [CrossRef] [Green Version]
- Tiwari, M.; Trivedi, P.K.; Pandey, A. Emerging tools and paradigm shift of gene editing in cereals, fruits, and horticultural crops for enhancing nutritional value and food security. Food Energy Secur. 2021, 10, e258. [Google Scholar] [CrossRef]
- French, E.; Kaplan, I.; Lyer-Pascuzzi, A.; Nakatsu, C.H.; Enders, L. Emerging strategies for precision microbiome management in diverse agroecosystems. Nat. Plants 2021, 7, 256–267. [Google Scholar] [CrossRef]
- Zha, F.C.; Rao, J.J.; Chen, B.C. Modification of pulse proteins for improved functionality and flavor profile: A comprehensive review. Compr. Rev. Food Sci. F 2021, 20, 3036–3060. [Google Scholar] [CrossRef]
- Lowry, G.V.; Avellan, A.; Gilbertson, L.M. Opportunities and challenges for nanotechnology in the agri-tech revolution. Nat. Nanotechnol. 2019, 14, 517–522. [Google Scholar] [CrossRef]


{kind=link}
| Genome Editing Technique | Description | Applications in Horticultural Crops | References |
|---|---|---|---|
| CRISPR-Cas9 | A versatile and widely used technique for precise DNA editing | Improve yield, disease resistance, quality traits, and stress tolerance | [19] |
| TALEN | Transcription activator-like effector nuclease for targeted editing | Modify specific genes for desired traits | [14] |
| Zinc Finger Nuclease (ZFNs) | Engineered DNA-binding proteins for targeted gene editing | Enhance disease resistance, improve nutritional value, gene disruption, and gene replacement | [20] |
| Crops | Modified Gene(s) | Trait/Function | Reference |
|---|---|---|---|
| Tomato | E8, Phytoene desaturase (PDS), SlDELLA | Enhanced fruit ripening, delayed fruit senescence, reduced plant height | [25] |
| Potato | StCDF1 | Increased tuberization and yield | [26] |
| Wheat | TaGW2, Puroindoline genes | Enhanced thousand grain weight, improved grain quality | [27] |
| Citrus | CsPDS | Improved disease resistance, reduced ethylene production | [28] |
| Strawberry | FaTM6 | Petal and stamen development | [29] |
| Grape | VvWRKY52, VvWRKY2 | Enhanced disease resistance, improved abiotic stress tolerance | [30] |
| Brassica oleracea | XccR5-89.2 | Improved resistance to blackleg disease | [31] |
| Mushroom (Agaricus bisporus) | Polyphenol oxidase (PPO) genes | Reduced browning and improved shelf life | [32] |
| Banana | MaACO1 | Promotes the shelf life of banana | [33] |
| Carrot | DcCCD4 | Different colored taproots in carrots | [34] |
| Strawberry | FaGAST1 | Increased fruit size | [35] |
| Cucumis melo | CmACO1 | Extends the shelf-life | [36] |
| Capsicum annuum | CaERF28 | Anthracnose resistance | [37] |
| Rose | RhEIN2 | Ethylene insensitivity in rose | [38] |
| Melon | eIF4E | Virus resistance and male sterility | [39] |
| Tomato | SlMAPK3 | Reduced drought tolerance | [40] |
| Brassica napus | FAD2 | Catalyzes the desaturation of oleic acid | [41] |
| Kiwifruit | AcBFT | Reduce plant dormancy | [42] |
| Tomato | SlMYC2 | Fruit Resistance to Botrytis cinerea | [43] |
| Soybean | GmFATB1 | Reduce saturated fatty acids | [44] |
| Kiwi fruit | AcCBF3 | Dwarf plants and enhanced freezing tolerance | [45] |
| Sweet Potato | IbGBSSI and IbSBEII | Improvement of starch quality | [46] |
| Papaya | phytoene desaturase (CpPDS) | Inducing a visually scorable albino phenotype | [47] |
| Eggplant | SmelPPO4, SmelPPO5, and SmelPPO6 | Reduces fruit flesh browning | [48] |
| Cassava | eIF4E | Reduces cassava brown streak disease symptom | [49] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Daniel, M.A.; Sebastin, R.; Yu, J.-K.; Soosaimanickam, M.P.; Chung, J.W. Enhancing Horticultural Crops through Genome Editing: Applications, Benefits, and Considerations. Horticulturae 2023, 9, 884. https://doi.org/10.3390/horticulturae9080884
Daniel MA, Sebastin R, Yu J-K, Soosaimanickam MP, Chung JW. Enhancing Horticultural Crops through Genome Editing: Applications, Benefits, and Considerations. Horticulturae. 2023; 9(8):884. https://doi.org/10.3390/horticulturae9080884
Chicago/Turabian StyleDaniel, Melvin A., Raveendar Sebastin, Ju-Kyung Yu, Maria Packiam Soosaimanickam, and Jong Wook Chung. 2023. "Enhancing Horticultural Crops through Genome Editing: Applications, Benefits, and Considerations" Horticulturae 9, no. 8: 884. https://doi.org/10.3390/horticulturae9080884