
In Vitro Growth and Regeneration of Brassica oleracea var. gongylodes: A Decade of Research

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Kohlrabi: Brassica’s Hidden Gem
2. In Vitro Growth and Regeneration as a Biotechnological Approach for Studying and Improvement of Kohlrabi
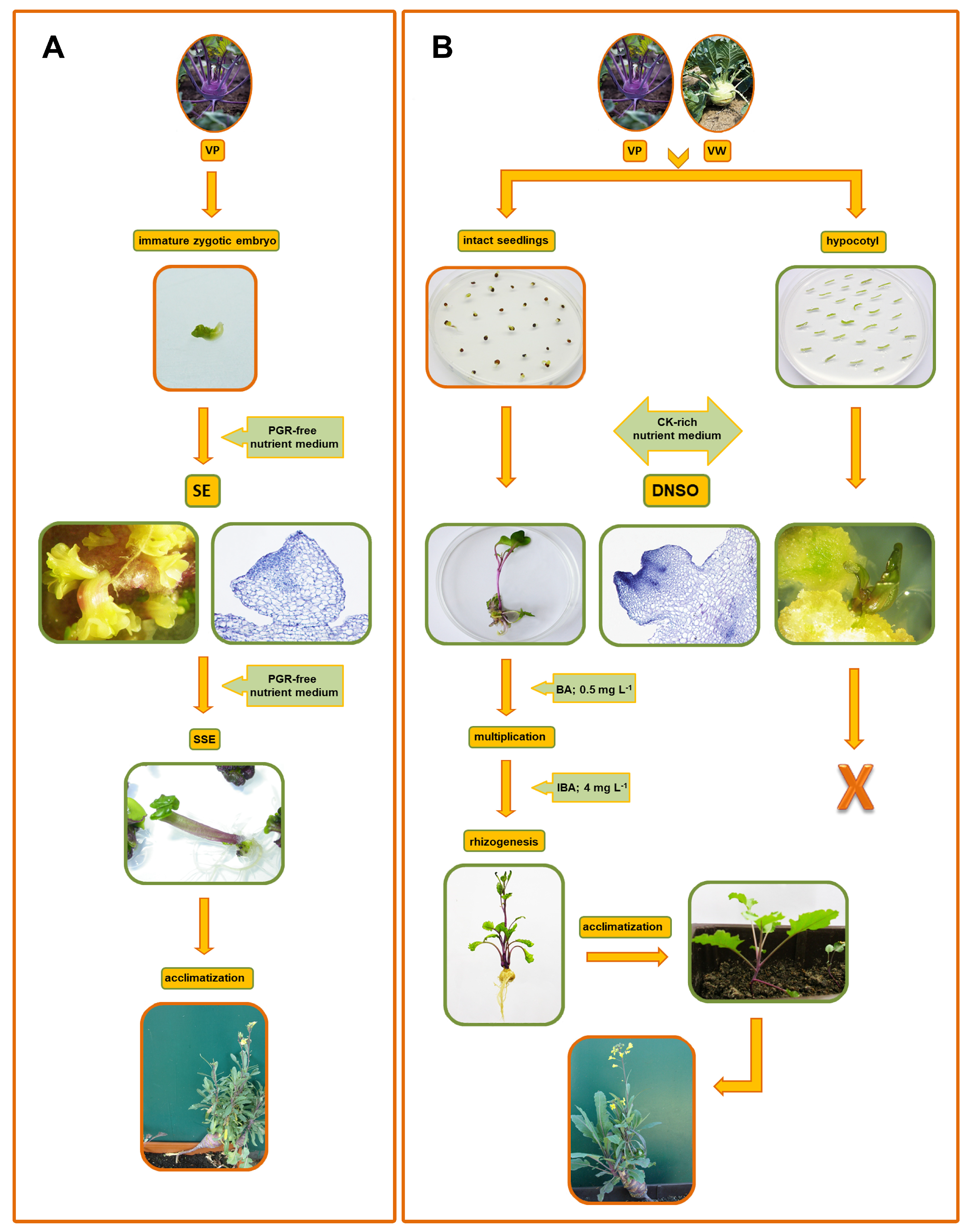
3. Direct Somatic Embryogenesis from Kohlrabi Immature Zygotic Embryos
4. One-Step De Novo Shoot Organogenesis in Kohlrabi
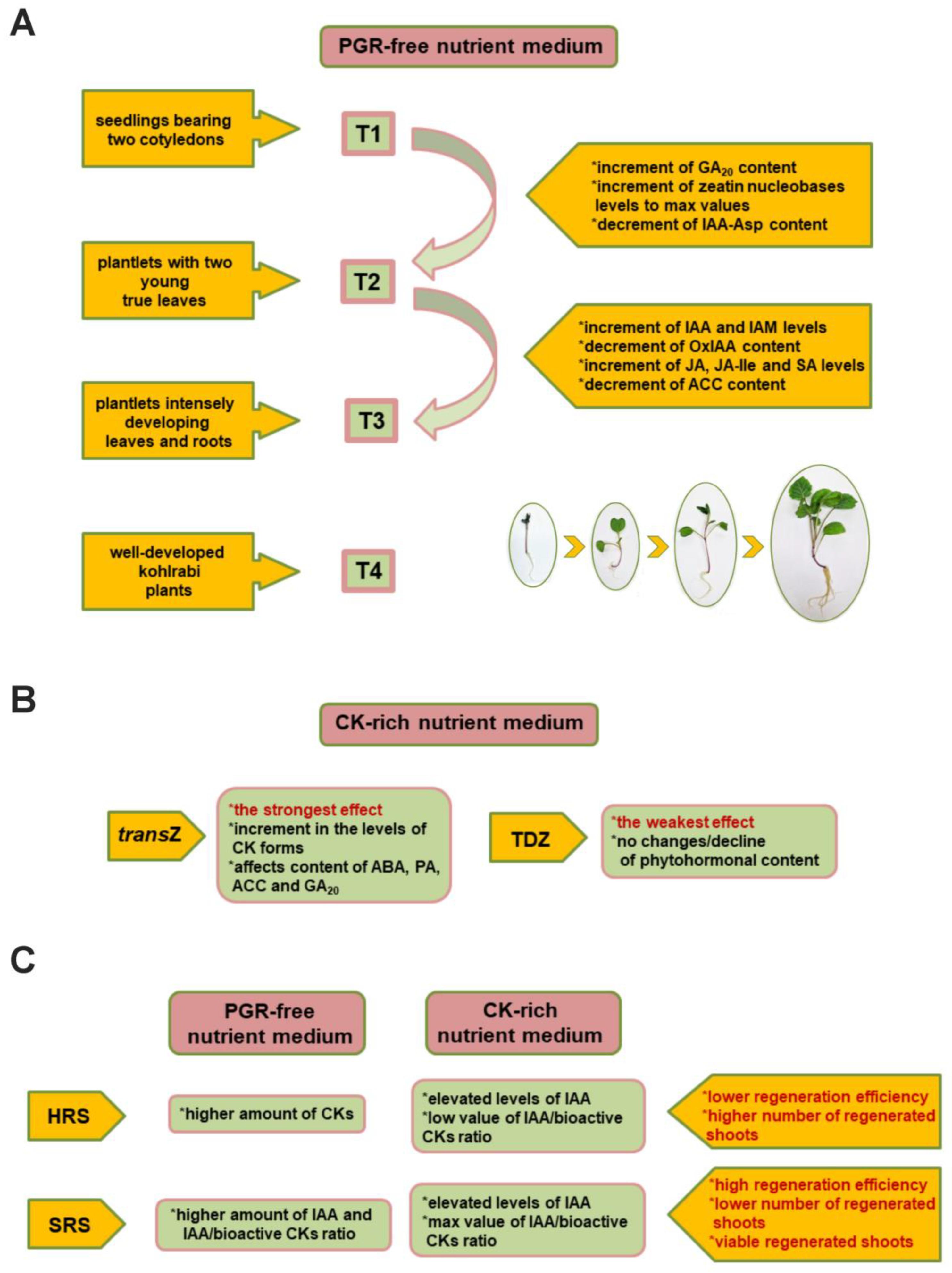
5. Hormonal Input during Growth and Regeneration of Kohlrabi In Vitro
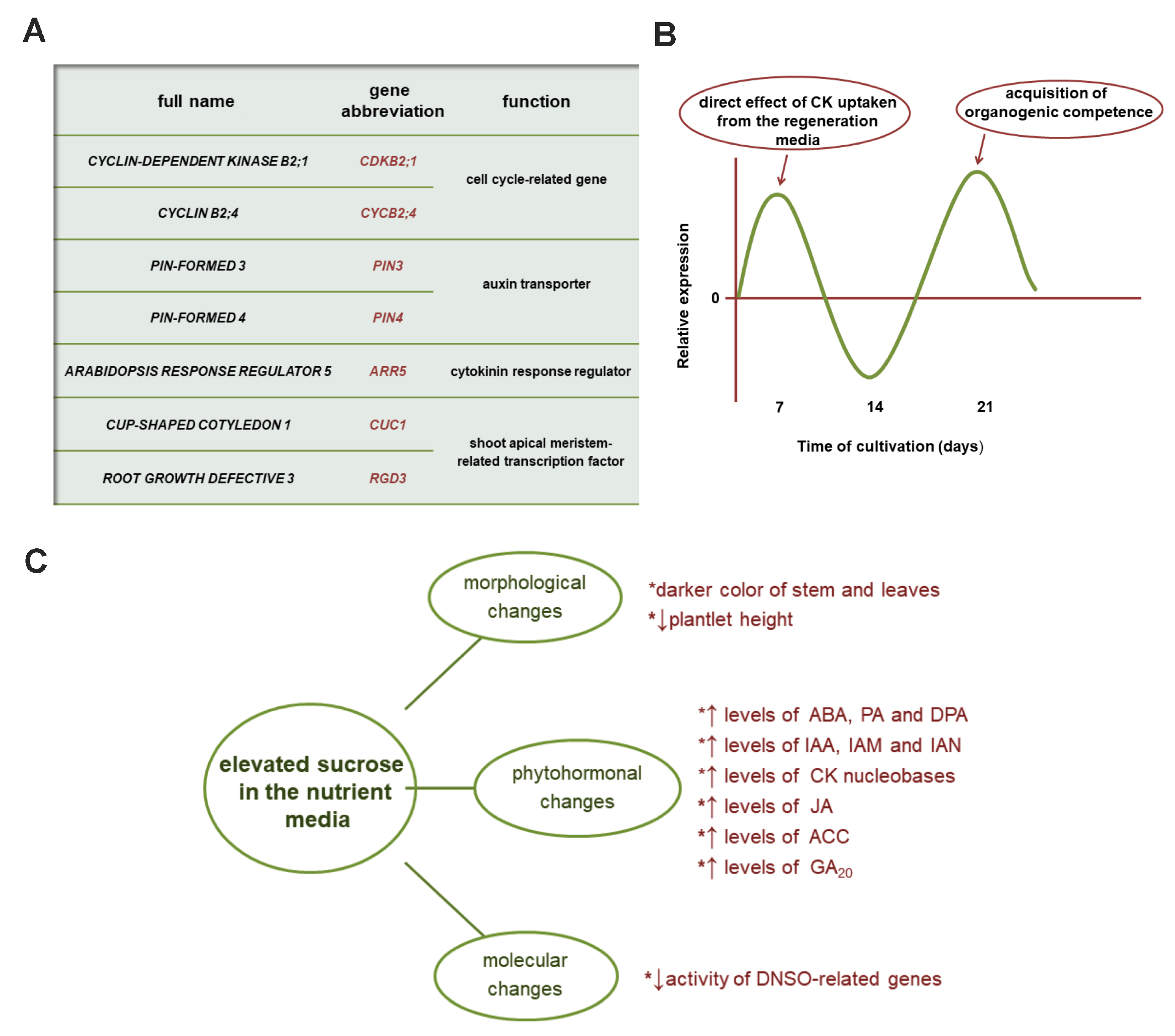
6. Genetic Insights into De Novo Shoot Organogenesis of Kohlrabi
7. Sucrose as an Important Component Affecting Multiple Aspects of Kohlrabi Growth and Regeneration In Vitro
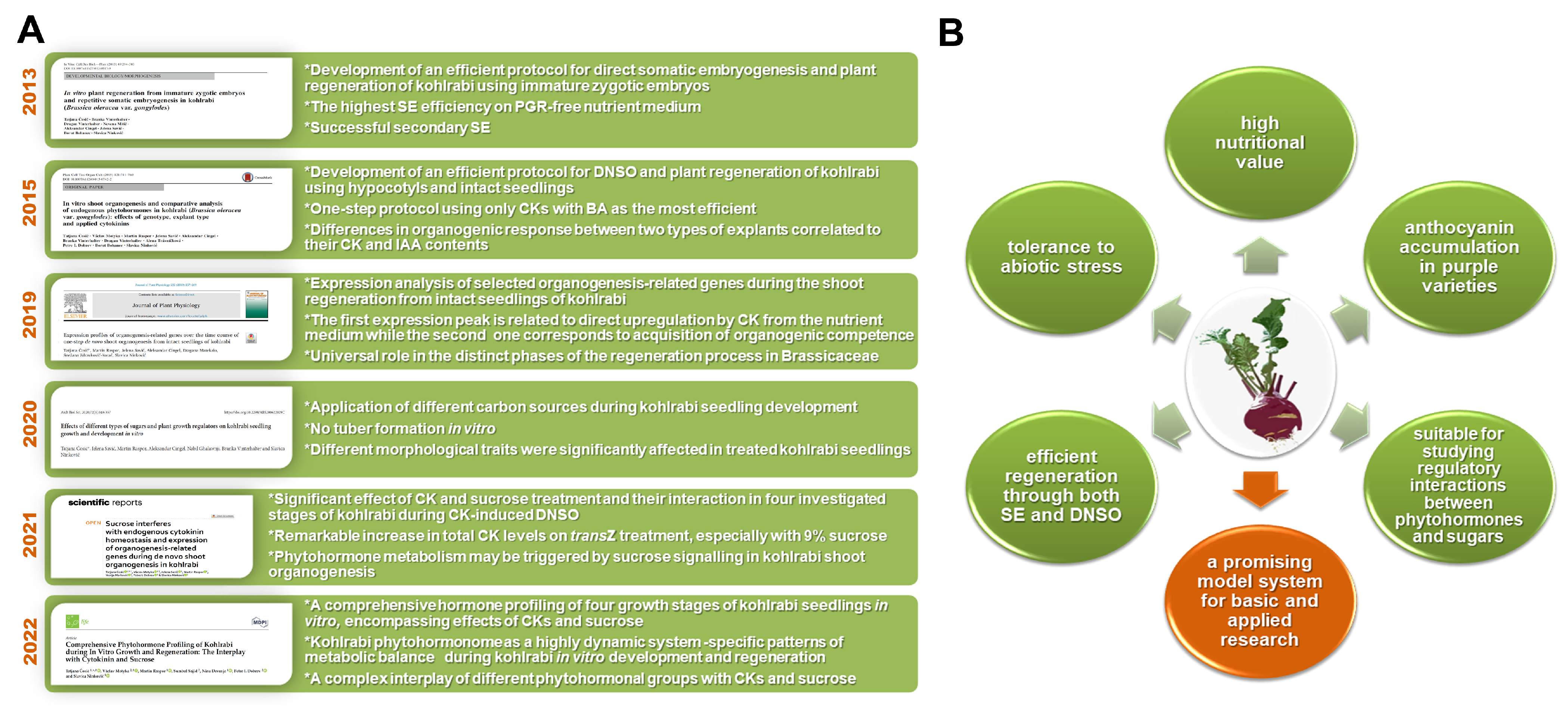
8. Conclusions and Further Perspectives
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Escalona, V.H.; Aguayo, E.; Artés, F. Extending the shelf life of kohlrabi stems by modified atmosphere packaging. J. Food Sci. 2007, 72, 308–313. [Google Scholar] [CrossRef]
- Hassan, E.A.; Hussein, A.; El-Awadi, M.E. Kohlrabi (Brassica caulorapa L): A potent anticancer vegetable processed in baked biscuits. Nat. Sci. 2011, 9, 149–157. Available online: http://www.sciencepub.net/nature/ns0908/27_6344ns0908_149_157.pdf (accessed on 25 April 2023).
- Paśko, P.; Galanty, A.; Tyszka-Czochara, M.; Żmudzki, P.; Zagrodzki, P.; Gdula-Argasińska, J.; Prochownik, E.; Gorinstein, S. Health Promoting vs. Anti-nutritive Aspects of Kohlrabi Sprouts, a Promising Candidate for Novel Functional Food. Plant Foods Hum. Nutr. 2021, 76, 76–82. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.H.; Ryu, D.K.; Park, S.; Ahn, K.G.; Lim, Y.P.; An, G. Composition analysis between kohlrabi (Brassica oleracea var. gongylodes) and Radish (Raphanus sativus). Kor. J. Hort. Sci. Technol. 2010, 28, 469–475. Available online: http://www.koreascience.or.kr/article/JAKO201018651618927.page (accessed on 25 April 2023).
- Selman, I.W.; Kulasegaram, S. Development of the stem tuber in kohlrabi. J. Exp. Bot. 1967, 18, 471–490. [Google Scholar] [CrossRef]
- Uddin, J.; Solaiman, A.H.M.; Hasanuzzaman, M. Plant characters and yield of kohlrabi (Brassica oleracea var. gongylodes) as affected by different organic manures. J. Hort. Sci. Ornam. Plants 2009, 1, 1–4. Available online: https://idosi.org/jhsop/jhsop1(1)09.htm (accessed on 25 April 2023).
- Abd, A.M.; Alalawy, H.H.; Hathal, A.A. Response yield of four cultivar kohlrabi (Brassica oleracea var. caulorapa L.) to plant density and foliar nutrition of seaweed. Plant Arch. 2020, 20, 4069–4076. Available online: http://www.plantarchives.org/SPL%20ISSUE%2020-2/670__4069-4076_.pdf (accessed on 25 April 2023).
- Yi, G.E.; Robin, A.H.K.; Yang, K.; Park, J.I.; Kang, J.G.; Yang, T.J.; Nou, I.S. Identification and expression analysis of glucosinolate biosynthetic genes and estimation of glucosinolate contents in edible organs of Brassica oleracea subspecies. Molecules 2015, 20, 13089–13111. [Google Scholar] [CrossRef] [Green Version]
- Iori, R.; Bernardi, R.; Gueyrard, D.; Rollin, P.; Palmieri, S. Formation of glucoraphanin by chemoselective oxidation of natural glucoerucin: A chemoenzymatic route to sulforaphane. Bioorg. Med. Chem. Lett. 1999, 9, 1047–1048. [Google Scholar] [CrossRef]
- Fahey, J.W.; Zalcmann, A.T.; Talalay, P. The chemical diversity and distribution of glucosinolates and isothiocyanates among plants. Phytochemistry 2001, 56, 5–51. [Google Scholar] [CrossRef]
- Shetty, P.; Banu, U.; Kedilaya, T.; Patheja, M. Characterization of peroxidase from Brassica oleracea gongylodes gives a lead for use of bromocresol purple as a novel substrate for peroxidase assay. J. Biochem. Technol. 2012, 4, 502–507. Available online: https://jbiochemtech.com/article/characterization-of-peroxidase-from-brassica-oleracea-gongylodes-gives-a-lead-for-use-of-bromocresol-purple-as-a-substrate-for-peroxidase-assay (accessed on 25 April 2023).
- Sathasivam, R.; Kim, M.C.; Yeo, H.J.; Nguyen, B.V.; Sohn, S.I.; Park, S.U.; Kim, J. Accumulation of Phenolic Compounds and Glucosinolates in Sprouts of Pale Green and Purple Kohlrabi (Brassica oleracea var. gongylodes) under Light and Dark Conditions. Agronomy 2021, 11, 1939. [Google Scholar] [CrossRef]
- Ben Sassi, A.; Ascrizzi, R.; Chiboub, W.; Mhamed, A.C.; ElAyeb, A.; Skhiri, F.; Saidani, M.T.; Mastouri, M.; Flamini, G. Volatiles, phenolic compounds, antioxidant and antibacterial properties of kohlrabi leaves. Nat. Prod. Res. 2022, 36, 3143–3148. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Hu, Z.; Zhu, M.; Zhu, Z.; Wang, Z.; Tian, S.; Chen, G. Antocyanin accumulation and molecular analysis of correlated genes in purple kohlrabi (Brassica oleracea var. gongylodes L.). J. Agric. Food Chem. 2015, 16, 4160–4169. [Google Scholar] [CrossRef]
- Rahim, M.A.; Robin, A.H.K.; Natarajan, S.; Jung, H.J.; Lee, J.; Kim, H.R.; Kim, H.T.; Park, J.I.; Nou, I.S. Identification and characterization of anthocyanin biosynthesis-related genes in kohlrabi. Appl. Biochem. Biotechnol. 2018, 184, 1120–1141. [Google Scholar] [CrossRef] [PubMed]
- Golob, A.; Novak, T.; Kacjan Maršić, N.; Šircelj, H.; Stibilj, V.; Jerše, A.; Kroflič, A.; Germ, M. Biofortification with selenium and iodine changes morphological properties of Brassica oleracea L. var. gongylodes and increases their contents in tubers. Plant Physiol. Biochem. 2020, 150, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Lošák, T.; Válka, T.; Elbl, J.; Kintl, A.; Keutgen, A.; Keutgen, N.; Demková, L.; Árvay, J.; Varga, L.; Hnátková, H.; et al. Fertilization with Magnesium- and Sulfur-Supplemented Digestate Increases the Yield and Quality of Kohlrabi. Sustainability 2020, 12, 5733. [Google Scholar] [CrossRef]
- Bhandari, S.; Bhandari, A.; Shrestha, J. Effect of different doses of triacontanol on growth and yield of kohlrabi (Brassica oleracea L. var. gongylodes). Heliyon 2021, 7, e08242. [Google Scholar] [CrossRef]
- Hearn, D.J.; O’Brien, P.; Poulsen, T.M. Comparative transcriptomics reveals shared gene expression changes during independent evolutionary origins of stem and hypocotyl/root tubers in Brassica (Brassicaceae). PLoS ONE 2018, 13, e0197166. [Google Scholar] [CrossRef] [Green Version]
- Nuruzzaman, M.; Sato, M.; Okamoto, S.; Hoque, M.; Shea, D.J.; Fujimoto, R.; Shimizu, M.; Fukai, E.; Okazaki, K. Comparative transcriptome analysis during tuberous stem formation in Kohlrabi (B. oleracea var. gongylodes) at early growth periods (seedling stages). Physiol. Plant. 2022, 174, e13770. [Google Scholar] [CrossRef]
- Nuruzzaman, M.; Kojima, M.; Sato, M.; Takebayashi, Y.; Hoque, M.; Okamoto, S.; Shea, D.J.; Fujimoto, R.; Fukai, E.; Sakakibara, H.; et al. Comparative anatomical and hormonal analyses between kohlrabi and broccoli seedlings: Relevance to kohlrabi stem tuber intiation. Sci. Hortic. 2023, 316, 112002. [Google Scholar] [CrossRef]
- Grosch, R.; Schneider, J.H.M.; Kofoet, A. Characterisation of Rhizoctonia solani anastomosis groups causing bottom rot in field-grown lettuce in Germany. Eur. J. Plant Pathol. 2004, 110, 53–62. [Google Scholar] [CrossRef]
- Ciaghi, S.; Schwelm, A.; Neuhauser, S. Transcriptomic response in symptomless roots of clubroot infected kohlrabi (Brassica oleracea var. gongylodes) mirrors resistant plants. BMC Plant Biol. 2019, 19, 288. [Google Scholar] [CrossRef] [Green Version]
- Neelakandan, A.K.; Wang, K. Recent progress in the understanding of tissue culture-induced genome level changes in plants and potential applications. Plant Cell Rep. 2012, 31, 597–620. [Google Scholar] [CrossRef]
- Fehér, A. Callus, dedifferentiation, totipotency, somatic embryogenesis: What these terms mean in the era of molecular plant biology? Front. Plant Sci. 2019, 10, 536. [Google Scholar] [CrossRef] [Green Version]
- Su, Y.H.; Tang, L.P.; Zhao, X.Y.; Zhang, X.S. Plant cell totipotency: Insights into cellular reprogramming. J. Integr. Plant Biol. 2021, 63, 228–243. [Google Scholar] [CrossRef] [PubMed]
- Long, Y.; Yang, Y.; Pan, G.; Shen, Y. New Insights Into Tissue Culture Plant-Regeneration Mechanisms. Front. Plant Sci. 2022, 13, 926752. [Google Scholar] [CrossRef] [PubMed]
- Gaj, M.D. Factors influencing somatic embryogenesis induction and plant regeneration with particular reference to Arabidopsis thaliana (L.) Heynh. Plant Growth Regul. 2004, 43, 27–47. [Google Scholar] [CrossRef]
- Ikeuchi, M.; Shibata, M.; Rymen, B.; Iwase, A.; Bågman, A.M.; Watt, L.; Coleman, D.; Favero, D.S.; Takahashi, T.; Ahnert, S.E.; et al. A gene regulatory network for cellular reprogramming in plant regeneration. Plant Cell Physiol. 2018, 59, 770–782. [Google Scholar] [CrossRef] [PubMed]
- Raspor, M.; Motyka, V.; Kaleri, A.R.; Ninković, S.; Tubić, L.; Cingel, A.; Ćosić, T. Integrating the roles for cytokinin and auxin in de novo shoot organogenesis: From hormone uptake to signaling outputs. Int. J. Mol. Sci. 2021, 22, 8554. [Google Scholar] [CrossRef]
- Su, Y.H.; Liu, Y.B.; Zhang, X.S. Auxin-cytokinin interaction regulates meristem development. Mol. Plant 2011, 4, 616–625. [Google Scholar] [CrossRef]
- Cardoza, V.; Stewart, C.N. Brassica biotechnology: Progress in cellular and molecular biology. Vitr. Cell. Dev. Biol. -Plant 2004, 40, 542–551. [Google Scholar] [CrossRef]
- Vinterhalter, D.; Sretenović-Rajičić, T.; Vinterhalter, B.; Ninković, S. Genetic transformation of Brassica oleracea vegetables. Transgenic Plant J. 2007, 1, 340–355. Available online: http://www.globalsciencebooks.info/Online/GSBOnline/OnlineTPJ_1_2.html (accessed on 25 April 2023).
- Mitra, M.; Gantait, S. Tissue Culture-Mediated Biotechnological Advancements in Genus Brassica. In Brassica Improvement; Wani, S.H., Thakur, A.K., Jeshima Khan, Y., Eds.; Springer: Cham, Switzerland, 2020; pp. 85–107. ISBN 978-3-030-34694-2. [Google Scholar] [CrossRef]
- Koh, W.L.; Loh, C.S. Direct somatic embryogenesis, plant regeneration and in vitro flowering in rapid-cycling Brassica napus. Plant Cell Rep. 2000, 19, 1177–1183. [Google Scholar] [CrossRef]
- Zeynali, M.; Maleki Zanjani, B.; Amiri, M.E.; Noruzian, M.; Aghajari, S.M. Influence of genotype and plant growth regulator on somatic embryogenesis in rapeseed (Brassica napus L.). Afr. J. Biotechnol. 2010, 9, 4050–4055. Available online: https://www.ajol.info/index.php/ajb/article/view/82564 (accessed on 21 April 2023).
- Yang, J.L.; Seong, E.S.; Kim, M.J.; Ghimire, B.K.; Kang, W.H.; Yu, C.Y.; Li, C.H. Direct somatic embryogenesis from pericycle cells of broccoli (Brassica oleracea L. var. italica) root explants. Plant Cell Tiss. Organ Cult. 2010, 100, 49–58. [Google Scholar] [CrossRef]
- Banjac, N.; Vinterhalter, B.; Krstić-Milošević, D.; Milojević, J.; Tubić, L.; Ghalawenji, N.; Zdravković-Korać, S. Somatic embryogenesis and shoot organogenesis from the hypocotyl slices and free radical scavenging activity of regenerants of collard greens (Brassica oleracea L. var. acephala). Plant Cell Tiss. Organ Cult. 2019, 137, 613–626. [Google Scholar] [CrossRef]
- Al-Hardan, G.M.; AL-Shamari, M.; Hanshil, M.A. Impact of culture medium and 2,4-D on direct somatic embryogenesis in red cabbage (Brassica oleracea var. capitata forma rubra). Plant Arch. 2020, 20, 3022–3026. Available online: http://www.plantarchives.org/SPL%20ISSUE%2020-2/501__3022-3026_.pdf (accessed on 25 April 2023).
- Pavlović, S.; Savić, J.; Milojević, J.; Vinterhalter, B.; Girek, Z.; Adžić, S.; Zečević, B.; Banjac, N. Introduction of the Nicotiana protein kinase (NPK1) gene by combining Agrobacterium-mediated transformation and recurrent somatic embryogenesis to enhance salt tolerance in cauliflower. Plant Cell Tiss. Organ Cult. 2020, 143, 635–651. [Google Scholar] [CrossRef]
- Choi, P.S.; Soh, W.Y.; Liu, J.R. Somatic embryogenesis and plant regeneration in cotyledonary explant cultures of Chinese cabbage. Plant Cell Tiss. Organ Cult. 1996, 44, 253–256. [Google Scholar] [CrossRef]
- Barro, F.; Martín, A. Response of different genotypes of Brassica carinata to microspore culture. Plant Breed. 1999, 118, 79–81. [Google Scholar] [CrossRef]
- Shah, R.U.; Munir, I. Efficient in vitro plant regeneration through somatic embryogenesis from callus induction method for Brassica carinata. Sarhad J. Agric. 2019, 35, 314–319. Available online: https://researcherslinks.com/current-issues/Efficient-in-Vitro-Plant-Regeneration-Through-Somatic-Embryogenesis-from-Callus-Induction-Method-for-Brassica-carinata/14/1/2076 (accessed on 25 April 2023). [CrossRef]
- Piovan, A.; Caniato, R.; Filippini, R. Somatic embryogenesis and glucosinolate/myrosinase system in vulnerable Brassica repanda subsp. glabrescens (Poldini) Gómez-Campo. Sci. Hortic. 2014, 172, 317–324. [Google Scholar] [CrossRef]
- Faisal, M.; Abdel-Salam, E.M.; Alatar, A.A.; Qahtan, A.A. Induction of somatic embryogenesis in Brassica juncea L. and analysis of regenerants using ISSR-PCR and flow cytometer. Saudi J. Biol. Sci. 2021, 28, 1147–1153. [Google Scholar] [CrossRef]
- Bhalla, P.L.; Singh, M.B. Agrobacterium-mediated transformation of Brassica napus and Brassica oleracea. Nat. Protoc. 2008, 3, 181–189. [Google Scholar] [CrossRef]
- Ghnaya, A.B.; Charles, G.; Branchard, M. Rapid shoot regeneration from thin cell layer explants excised from petioles and hypocotyls in four cultivars of Brassica napus L. Plant Cell Tiss. Organ Cult. 2008, 92, 25–30. [Google Scholar] [CrossRef]
- Pavlović, S.; Vinterhalter, B.; Mitić, N.; Adžić, S.; Pavlović, N.; Zdravković, M.; Vinterhalter, D. In vitro shoot regeneration from seedling explants in Brassica vegetables: Red cabbage, broccoli, Savoy cabbage and cauliflower. Arch. Biol. Sci. 2010, 62, 337–345. [Google Scholar] [CrossRef]
- Glendening, T.M.; Sjolund, R. In vitro propagation of kohlrabi from leaf explants. HortScience 1988, 23, 772. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Khan, M.; Guo, B.; Bokhari, S.A.; Khan, M.A. Efficient regeneration and antioxidative enzyme activities in Brassica rapa var. turnip. Plant Cell Tiss. Organ Cult. 2011, 105, 337–344. [Google Scholar] [CrossRef]
- Lillo, C.; Shahin, E.A. Rapid regeneration of plants from hypocotyl protoplasts and root segments of cabbage. HortScience 1986, 21, 315–317. [Google Scholar] [CrossRef]
- Cogbill, S.; Faulcon, T.; Jones, G.; McDaniel, M.; Harmon, G.; Blackmon, R.; Young, M. Adventitious shoot regeneration from cotyledonary explants of rapid-cycling fast plants of Brassica rapa L. Plant Cell Tiss. Organ Cult. 2010, 101, 127–133. [Google Scholar] [CrossRef]
- Sharma, S.; Gambhir, G.; Srivastava, D.K. High Frequency Organogenesis in Cotyledon and Hypocotyl Explants of Cabbage (Brassica oleracea L. var. capitata). Natl. Acad. Sci. Lett. 2014, 37, 5–12. [Google Scholar] [CrossRef]
- Hu, Q.; Anderson, S.B.; Hansen, L.N. Plant regeneration capacity of mesophyll protoplasts from Brassica napus and related species. Plant Cell Tiss. Organ Cult. 1999, 59, 189–196. [Google Scholar] [CrossRef]
- Kumar, P.; Srivastava, D.K. High frequency organogenesis in hypocotyl, cotyledon, leaf and petiole explants of broccoli (Brassica oleracea L. var. italica), an important vegetable crop. Physiol. Mol. Biol. Plants 2015, 21, 279–285. [Google Scholar] [CrossRef] [Green Version]
- Bhalla, P.L.; de Weerd, N. In vitro propagation of cauliflower, Brassica oleracea var. botrytis for hybrid seed production. Plant Cell Tiss. Organ Cult. 1999, 56, 89–95. [Google Scholar] [CrossRef]
- Werbrouck, S.P.O.; Strnad, M.; Van Onckelen, H.A.; Debergh, P.C. Meta-topolin, an alternative to benzyladenine in tissue culture? Physiol. Plant. 1996, 98, 291–297. [Google Scholar] [CrossRef]
- Klíma, M.; Vyvadilová, M.; Kučera, V. Production and utilization of doubled haploids in Brassica oleracea vegetables. Hortic. Sci. 2004, 31, 119–123. [Google Scholar] [CrossRef] [Green Version]
- Ćosić, T.; Vinterhalter, B.; Vinterhalter, D.; Mitić, N.; Cingel, A.; Savić, J.; Bohanec, B.; Ninković, S. In vitro plant regeneration from immature zygotic embryos and repetitive somatic embryogenesis in kohlrabi (Brassica oleracea var. gongylodes). Vitr. Cell. Dev. Biol. Plant 2013, 49, 294–303. [Google Scholar] [CrossRef]
- Ćosić, T.; Motyka, V.; Raspor, M.; Savić, J.; Cingel, A.; Vinterhalter, B.; Vinterhalter, D.; Trávníčková, A.; Dobrev, P.I.; Bohanec, B.; et al. In vitro shoot organogenesis and comparative analysis of endogenous phytohormones in kohlrabi (Brassica oleracea var. gongylodes): Effects of genotype, explant type and applied cytokinins. Plant Cell Tiss. Organ Cult. 2015, 121, 741–760. [Google Scholar] [CrossRef]
- Ćosić, T.; Raspor, M.; Savić, J.; Cingel, A.; Matekalo, D.; Zdravković-Korać, S.; Ninković, S. Expression profiles of organogenesis-related genes over the time course of one-step de novo shoot organogenesis from intact seedlings of kohlrabi. J. Plant Physiol. 2019, 232, 257–269. [Google Scholar] [CrossRef]
- Ćosić, T.; Savić, J.; Raspor, M.; Cingel, A.; Ghalawnji, N.; Vinterhalter, B.; Ninković, S. Effects of different types of sugars and plant growth regulators on kohlrabi seedling growth and development in vitro. Arch. Biol. Sci. 2020, 72, 349–357. [Google Scholar] [CrossRef]
- Ćosić, T.; Motyka, V.; Savić, J.; Raspor, M.; Marković, M.; Dobrev, P.I.; Ninković, S. Sucrose interferes with endogenous cytokinin homeostasis and expression of organogenesis-related genes during de novo shoot organogenesis in kohlrabi. Sci. Rep. 2021, 11, 6494. [Google Scholar] [CrossRef]
- Ćosić, T.; Motyka, V.; Raspor, M.; Sajid, S.; Devrnja, N.; Dobrev, P.I.; Ninković, S. Comprehensive Phytohormone Profiling of Kohlrabi during in vitro Growth and Regeneration: The Interplay with Cytokinin and Sucrose. Life 2022, 12, 1585. [Google Scholar] [CrossRef]
- Gaj, M.D. Direct somatic embryogenesis as a rapid and efficient system for in vitro regeneration of Arabidopsis thaliana. Plant Cell Tiss. Organ Cult. 2001, 64, 39–46. [Google Scholar] [CrossRef]
- Salaün, C.; Lepiniec, L.; Dubreucq, B. Genetic and molecular control of somatic embryogenesis. Plants 2021, 10, 1467. [Google Scholar] [CrossRef] [PubMed]
- Elhiti, M.; Stasolla, C. Transduction of Signals during Somatic Embryogenesis. Plants 2022, 11, 178. [Google Scholar] [CrossRef]
- Fehér, A. Somatic embryogenesis—Stress-induced remodeling of plant cell fate. Biochim. Biophys. Acta Gene Regul. Mech. 2015, 1849, 385–402. [Google Scholar] [CrossRef]
- Garin, E.; Isabel, N.; Plourde, A. Screening of large numbers of seed families of Pinus strobus L. for somatic embryogenesis from immature and mature zygotic embryos. Plant Cell Rep. 1998, 18, 37–43. [Google Scholar] [CrossRef]
- Choi, Y.E.; Yang, D.C.; Park, J.C.; Soh, W.Y.; Choi, K.T. Regenerative ability of somatic single and multiple embryos from cotyledons of Korean ginseng on hormone-free medium. Plant Cell Rep. 1998, 17, 544–551. [Google Scholar] [CrossRef]
- Pillon, E.; Terzi, M.; Baldan, B.; Mariani, P.; Lo Schiavo, F. A protocol for obtaining embryogenic cell lines from Arabidopsis. Plant J. 1996, 9, 573–577. [Google Scholar] [CrossRef] [PubMed]
- Mordhorst, A.P.; Voerman, K.J.; Hartog, M.V.; Meijer, E.A.; van Went, J.; Koornneef, M.; De Vries, S.C. Somatic embryogenesis in Arabidopsis thaliana is facilitated by mutations in genes repressing meristematic cell divisions. Genetics 1998, 149, 549–563. [Google Scholar] [CrossRef]
- Raemakers, C.J.J.M.; Jacobsen, E.; Visser, R.G.F. Secondary somatic embryogenesis and applications in plant breeding. Euphytica 1995, 81, 93–107. [Google Scholar] [CrossRef]
- von Arnold, S.; Clapham, D.; Egertsdotter, U.; Mo, L.H. Somatic embryogenesis in conifers—A case study of induction and development of somatic embryos in Picea abies. Plant Growth Regul. 1996, 20, 3–9. [Google Scholar] [CrossRef]
- Burbulis, N.; Kupriene, R. Induction of somatic embryos on in vitro cultured zygotic embryos of spring Brassica napus. Acta Univ. Latv. Biol. 2005, 691, 137–143. Available online: https://eeb.lu.lv/EEB/2005/Burbulis2.shtml (accessed on 21 April 2023).
- Rai, M.K.; Akhtar, N.; Jaiswal, V.S. Somatic embryogenesis and plant regeneration in Psidium guajava L. cv. Banarasi local. Sci. Hortic. 2007, 113, 129–133. [Google Scholar] [CrossRef]
- Kim, S.W.; Oh, S.C.; Liu, J.R. Control of direct and indirect somatic embryogenesis by exogenous growth regulators in immature zygotic embryo cultures of rose. Plant Cell Tiss. Organ Cult. 2003, 74, 61–66. [Google Scholar] [CrossRef]
- Konar, R.N.; Thomas, E.; Street, H.E. Origin and structure of embryoids arising from epidermal cells of the stem of Ranunculus sceleratus L. J. Cell Sci. 1972, 11, 77–93. [Google Scholar] [CrossRef]
- Sugimoto, K.; Jiao, Y.; Meyerowitz, E.M. Arabidopsis regeneration from multiple tissues occurs via a root development pathway. Dev. Cell 2010, 18, 463–471. [Google Scholar] [CrossRef] [Green Version]
- Fehér, A.; Pasternak, T.P.; Dudits, D. Transition of somatic plant cells to an embryogenic state. Plant Cell Tiss. Organ Cult. 2003, 74, 201–228. [Google Scholar] [CrossRef]
- Méndez-Hernández, H.A.; Ledezma-Rodríguez, M.; Avilez-Montalvo, R.N.; Juárez-Gómez, Y.L.; Skeete, A.; Avilez-Montalvo, J.; De-la-Peña, C.; Loyola-Vargas, V.M. Signaling Overview of Plant Somatic Embryogenesis. Front. Plant Sci. 2019, 10, 77. [Google Scholar] [CrossRef] [Green Version]
- Simonović, A.D.; Trifunović-Momčilov, M.M.; Filipović, B.K.; Marković, M.P.; Bogdanović, M.D.; Subotić, A.R. Somatic Embryogenesis in Centaurium erythraea Rafn—Current Status and Perspectives: A Review. Plants 2021, 10, 70. [Google Scholar] [CrossRef] [PubMed]
- Choi, P.S.; Min, S.R.; Ahn, M.Y.; Soh, W.Y.; Liu, J.R. Somatic embryogenesis and plant regeneration in immature zygotic embryo, ovule, and anther filament cultures of Chinese cabbage. Sci. Hortic. 1998, 72, 151–155. [Google Scholar] [CrossRef]
- Rodriguez, A.P.M.; Wetzstein, H.Y. The effect of auxin type and concentration on pecan (Carya illinoinensis) somatic embryo morphology and subsequent conversion into plants. Plant Cell Rep. 1994, 13, 607–611. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, A.P.M.; Wetzstein, H.Y. A morphological and histological comparison of the initiation and development of pecan (Carya illinoinensis) somatic embryogenic cultures induced with naphthaleneacetic acid or 2,4-dichlorophenoxyacetic acid. Protoplasma 1998, 204, 71–83. [Google Scholar] [CrossRef]
- Choi, Y.E.; Kim, H.S.; Soh, W.Y.; Yang, D.C. Developmental and structural aspects of somatic embryos formed on medium containing 2,3,5-triiodobenzoic acid. Plant Cell Rep. 1997, 16, 738–744. [Google Scholar] [CrossRef] [PubMed]
- von Arnold, S.; Sabala, I.; Bozhkov, P.; Dyachok, J.; Filonova, L. Developmental pathways of somatic embryogenesis. Plant Cell Tiss. Organ Cult. 2002, 69, 233–249. [Google Scholar] [CrossRef]
- Jiménez, V.M. Involvement of plant hormones and plant growth regulators on in vitro somatic embryogenesis. Plant Growth Regul. 2005, 47, 91–110. [Google Scholar] [CrossRef]
- Ninković, S.; Miljuš-Djukić, J.; Nešković, M. Genetic transformation of alfalfa somatic embryos and their clonal propagation through repetitive somatic embryogenesis. Plant Cell Tiss. Organ Cult. 1995, 42, 255–260. [Google Scholar] [CrossRef]
- Uzelac, B.; Ninković, S.; Smigocki, A.; Budimir, S. Origin and development of secondary somatic embryos in transformed embryogenic cultures of Medicago sativa. Biol. Plant. 2007, 51, 1–6. [Google Scholar] [CrossRef]
- Bogdanović, M.D.; Ćuković, K.B.; Subotić, A.R.; Dragićević, M.B.; Simonović, A.D.; Filipović, B.K.; Todorović, S.I. Secondary Somatic Embryogenesis in Centaurium erythraea Rafn. Plants 2021, 10, 199. [Google Scholar] [CrossRef]
- Agarwal, S.; Kanwar, K.; Sharma, D.R. Factors affecting secondary somatic embryogenesis and embryo maturation in Morus alba L. Sci. Hortic. 2004, 102, 359–368. [Google Scholar] [CrossRef]
- Pavlović, S.; Vinterhalter, B.; Zdravković-Korać, S.; Vinterhalter, D.; Zdravković, J.; Cvikić, D.; Mitić, N. Recurrent somatic embryogenesis and plant regeneration from immature zygotic embryos of cabbage (Brassica oleracea var. capitata) and cauliflower (Brassica oleracea var. botrytis). Plant Cell Tiss. Organ Cult. 2013, 113, 397–406. [Google Scholar] [CrossRef]
- Vasic, D.; Alibert, G.; Skoric, D. Protocols for efficient repetitive and secondary somatic embryogenesis in Helianthus maximiliani (Schrader). Plant Cell Rep. 2001, 20, 121–125. [Google Scholar] [CrossRef]
- Thorpe, T.A. Somatic embryogenesis: Morphogenesis, physiology, biochemistry and molecular biology. Kor. J. Plant Tiss. Cult. 2000, 27, 245–258. [Google Scholar]
- Martinelli, L.; Candioli, E.; Costa, D.; Poletti, V.; Rascio, N. Morphogenic competence of Vitis rupestris S. secondary somatic embryos with a long culture history. Plant Cell Rep. 2001, 20, 279–284. [Google Scholar] [CrossRef]
- Haccius, B. Question of unicellular origin of non-zigotic embryos in callus cultures. Phytomorphology 1978, 28, 74–81. [Google Scholar]
- Toonen, M.A.J.; Hendriks, T.; Schmidt, E.D.L.; Verhoeven, H.A.; van Kammen, A.; de Vries, S.C. Description of somatic-embryo-forming single cells in carrot suspension cultures employing video cell tracking. Planta 1994, 194, 565–572. [Google Scholar] [CrossRef]
- Chan, J.L.; Saénz, L.; Talavera, C.; Hornung, R.; Robert, M.; Oropeza, C. Regeneration of coconut (Cocos nucifera L.) from plumule explants through somatic embryogenesis. Plant Cell Rep. 1998, 17, 515–521. [Google Scholar] [CrossRef]
- Kim, Y.W.; Youn, Y.; Noh, E.R.; Kim, J.C. Somatic embryogenesis and plant regeneration from immature embryos of five families of Quercus acutissima. Plant Cell Rep. 1997, 16, 869–873. [Google Scholar] [CrossRef]
- Yang, D.C.; Choi, Y.E. Production of transgenic plants via Agrobacterium rhizogenes-mediated transformation of Panax ginseng. Plant Cell Rep. 2000, 19, 491–496. [Google Scholar] [CrossRef]
- Ahmed, R.; Gupta, D.S.; De, D.N. Somatic embryogenesis and plant regeneration from leaf derived callus of winged bean [Psophocarpus tetragonolobus (L.) DC.]. Plant Cell Rep. 1996, 15, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Sarasan, V.; Roberts, A.V.; Rout, G.R. Methyl laurate and 6-benzyladenine promote the germination of somatic embryos of a hybrid rose. Plant Cell Rep. 2001, 20, 183–186. [Google Scholar] [CrossRef]
- Skoog, F.; Miller, C.O. Chemical regulation of growth and organ formation in plant tissues cultured in vitro. Symp. Soc. Exp. Biol. 1957, 11, 118–130. [Google Scholar]
- Christianson, M.L.; Warnick, D.A. Temporal requirement for phytohormone balance in the control of organogenesis in vitro. Dev. Biol. 1985, 112, 494–497. [Google Scholar] [CrossRef]
- Zhao, X.Y.; Su, Y.H.; Cheng, Z.J.; Zhang, X.S. Cell fate switch during in vitro plant organogenesis. J. Integr. Plant Biol. 2008, 50, 816–824. [Google Scholar] [CrossRef]
- Oinam, G.; Yeung, E.; Kurepin, L.; Haslam, T.; Lopez-Villalobos, A. Adventitious root formation in ornamental plants: I. General overview and recent successes. Propag. Ornam. Plants 2011, 11, 78–90. Available online: http://www.journal-pop.org/2011_11_2_78-90.html (accessed on 25 April 2023).
- Cary, A.J.; Che, P.; Howell, S.H. Developmental events and shoot apical meristem gene expression patterns during shoot development in Arabidopsis thaliana. Plant J. 2002, 32, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Che, P.; Gingerich, D.J.; Lall, S.; Howell, S.H. Global and hormone-induced gene expression changes during shoot development in Arabidopsis. Plant Cell 2002, 14, 2771–2785. [Google Scholar] [CrossRef]
- Valvekens, D.; Van Montagu, M.; Van Lijsebettens, M. Agrobacterium tumefaciens-mediated transformation of Arabidopsis thaliana root explants by using kanamycin selection. Proc. Natl. Acad. Sci. USA 1988, 85, 5536–5540. [Google Scholar] [CrossRef] [Green Version]
- Ćosić, T.; Raspor, M. The role of auxin and cytokinin signaling components in de novo shoot organogenesis. In Auxins, Cytokinins and Gibberellins Signaling in Plants; Aftab, T., Ed.; Springer: Cham, Switzerland, 2022; pp. 47–75. ISBN 978-3-031-05427-3. [Google Scholar] [CrossRef]
- Alvarez, J.M.; Bueno, N.; Cuesta, C.; Feito, I.; Ordás, R.J. Hormonal and gene dynamics in de novo shoot meristem formation during adventitious caulogenesis in cotyledons of Pinus pinea. Plant Cell Rep. 2020, 39, 527–541. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Lee, J.; Jie, E.Y.; Choi, S.H.; Jiang, L.; Ahn, W.S.; Kim, C.Y.; Kim, S.W. Temporal and spatial expression analysis of shoot-regeneration regulatory genes during the adventitious shoot formation in hypocotyl and cotyledon explants of tomato (cv. Micro-Tom). Int. J. Mol. Sci. 2020, 21, 5309. [Google Scholar] [CrossRef]
- Subrahmanyeswari, T.; Gantait, S.; Sarkar, S.; Bhattacharyya, S. Accelerated mono-phasic in vitro mass production of banana propagules and their morpho-cyto-genetic stability assessment. South Afr. J. Bot. 2022, 146, 794–806. [Google Scholar] [CrossRef]
- Sharma, K.K.; Thorpe, T.A. In vitro regeneration of shoot buds and plantlets from seedling root segments of Brassica napus L. Plant Cell Tiss. Organ Cult. 1989, 18, 129–141. [Google Scholar] [CrossRef]
- Mollika, S.R.; Sarker, R.H.; Hoque, M.I. In vitro plant regeneration in Brassica spp. Plant Tissue Cult. Biotech. 2011, 21, 127–134. [Google Scholar] [CrossRef]
- Ravanfar, S.A.; Aziz, M.A.; Kadir, M.A.; Rashid, A.A.; Haddadi, F. In vitro adventitious shoot regeneration and acclimatisation of Brassica oleracea subsp. italica cv. Green Marvel. Afr. J. Biotechnol. 2011, 10, 5614–5619. Available online: https://www.ajol.info/index.php/ajb/article/view/94349 (accessed on 30 March 2023).
- Ravanfar, S.A.; Aziz, M.A.; Rashid, A.A.; Salim, S. In vitro adventitious shoot regeneration from cotyledon explant of Brassica oleracea subsp. italica and Brassica oleracea subsp. capitata using TDZ and NAA. Pak. J. Bot. 2014, 46, 329–335. Available online: http://www.pakbs.org/pjbot/archives2.php?vol=46&iss=1&yea=2014 (accessed on 25 April 2023).
- Burnett, L.; Arnoldo, M.; Yarrow, S.; Huang, B. Enhancement of shoot regeneration from cotyledon explants of Brassica rapa ssp. oleifera through pretreatment with auxin and cytokinin and use of ethylene inhibitors. Plant Cell Tiss. Organ Cult. 1994, 37, 253–256. [Google Scholar] [CrossRef]
- Teo, W.; Lakshmanan, P.; Kumar, P.; Goh, C.J.; Swarup, S. Direct shoot formation and plant regeneration from cotyledon explants of rapid-cycling Brassica rapa. Vitr. Cell. Dev. Biol. Plant 1997, 33, 288–292. [Google Scholar] [CrossRef]
- Ono, Y.; Takahata, Y.; Kaizuma, N. Effect of genotype on shoot regeneration from cotyledonary explants of rapeseed (Brassica napus L.). Plant Cell Rep. 1994, 14, 13–17. [Google Scholar] [CrossRef]
- Sharma, K.K.; Bhojwani, S.S.; Thorpe, T.A. Factors affecting high frequency differentiation of shoots and roots from cotyledon explants of Brassica juncea (L.) Czern. Plant Sci. 1990, 66, 247–253. [Google Scholar] [CrossRef] [Green Version]
- Fuller, M.P.; Strullu, D.G.; Schlesser, A. The regeneration of shoots from seedlings explants of cauliflower. Crucif. Newslett. 1994, 16, 53–54. Available online: https://www.brassica.info/info/publications/cruciferae-newsletter.php (accessed on 25 April 2023).
- Kamal, G.B.; Illich, K.G.; Asadollah, A. Effects of genotype, explant type and nutrient medium components on canola (Brassica napus L.) shoot in vitro organogenesis. Afr. J. Biotechnol. 2007, 6, 861–867. Available online: https://www.ajol.info/index.php/ajb/article/view/56918 (accessed on 21 April 2023).
- Jin, R.G.; Liu, Y.B.; Tabashnik, B.E.; Borthakur, D. Development of transgenic cabbage (Brassica oleracea var. capitata) for insect resistance by Agrobacterium tumefaciens-mediated transformation. Vitr. Cell. Dev. Biol. Plant 2000, 36, 231–237. [Google Scholar] [CrossRef]
- Khan, M.R.; Rashid, H.; Ansar, M.; Chaudry, Z. High frequency shoot regeneration and Agrobacterium-mediated DNA transfer in Canola (Brassica napus). Plant Cell Tiss. Organ Cult. 2003, 75, 223–231. [Google Scholar] [CrossRef]
- Bedir, H.; Ari, E.; Vural, G.E.; Seguí-Simarro, J.M. Effect of the genotype, explant source and culture medium in somatic embryogenesis and organogenesis in Vaccaria hispanica (Mill.) Rauschert. Plant Cell Tiss. Organ Cult. 2022, 150, 329–343. [Google Scholar] [CrossRef]
- Yang, H.; Yuan, H.; Du, C.; Liang, L.; Chen, M.; Zou, L. Development of a Highly Efficient Shoot Organogenesis System for an Ornamental Aeschynanthus pulcher (Blume) G. Don Using Leaves as Explants. Plants 2022, 11, 2456. [Google Scholar] [CrossRef]
- Guo, B.; Abbasi, B.H.; Zeb, A.; Xu, L.L.; Wei, Y.H. Thidiazuron: A multi-dimensional plant growth regulator. Afr. J. Biotechnol. 2011, 10, 8984–9000. [Google Scholar] [CrossRef] [Green Version]
- Motyka, V.; Kamínek, M. Characterization of cytokinin oxidase from tobacco and poplar callus cultures. In Physiology and Biochemistry of Cytokinins in Plants; Kamínek, M., Mok, D.W.S., Zažímalová, E., Eds.; SPB Academic Publishing: The Hague, The Netherlands, 1992; pp. 33–39. ISBN 978-9051030662. [Google Scholar]
- Motyka, V.; Kamínek, M. Cytokinin oxidase from auxin- and cytokinin-dependent callus cultures of tobacco (Nicotiana tabacum L.). J. Plant Growth Regul. 1994, 13, 1–9. [Google Scholar] [CrossRef]
- Nisler, J.; Kopečný, D.; Končitíková, R.; Zatloukal, M.; Bazgier, V.; Berka, K.; Zalabák, D.; Briozzo, P.; Strnad, M.; Spíchal, L. Novel thidiazuron-derived inhibitors of cytokinin oxidase/dehydrogenase. Plant Mol. Biol. 2016, 92, 235–248. [Google Scholar] [CrossRef]
- Magyar-Tábori, K.; Dobránszki, J.; Teixeira da Silva, J.A.; Bulley, S.M.; Hudák, I. The role of cytokinins in shoot organogenesis in apple. Plant Cell Tiss. Organ Cult. 2010, 101, 251–267. [Google Scholar] [CrossRef]
- Dewir, Y.H.; Nurmansyah; Naidoo, Y.; Teixeira da Silva, J.A. Thidiazuron-induced abnormalities in plant tissue cultures. Plant Cell Rep. 2018, 37, 1451–1470. [Google Scholar] [CrossRef]
- Coleman, G.D.; Ernst, S.G. In vitro shoot regeneration of Populus deltoides: Effect of cytokinin and genotype. Plant Cell Rep. 1989, 8, 459–462. [Google Scholar] [CrossRef] [PubMed]
- Souza, B.M.; Kraus, J.E.; Endres, L.; Mercier, H. Relationships between endogenous hormonal levels and axillary bud development of Ananas comosus nodal segments. Plant Physiol. Biochem. 2003, 41, 733–739. [Google Scholar] [CrossRef]
- Suri, S.S.; Saini, A.R.K.; Ramawat, K.G. High frequency regeneration and Agrobacterium tumefaciens-mediated transformation of broccoli (Brassica oleracea var. italica). Eur. J. Hortic. Sci. 2005, 70, 71–78. Available online: https://www.pubhort.org/ejhs/2005/29079.htm (accessed on 30 March 2023).
- Nikolić, R.; Mitić, N.; Miletić, R.; Nešković, M. Effects of cytokinins on in vitro seed germination and early seedling morphogenesis in Lotus corniculatus L. J. Plant Growth Regul. 2006, 25, 187–194. [Google Scholar] [CrossRef]
- Werner, T.; Motyka, V.; Strnad, M.; Schmülling, T. Regulation of plant growth by cytokinin. Proc. Natl. Acad. Sci. USA 2001, 98, 10487–10492. [Google Scholar] [CrossRef] [Green Version]
- Werner, T.; Motyka, V.; Laucou, V.; Smets, R.; Van Onckelen, H.; Schmülling, T. Cytokinin-deficient transgenic Arabidopsis plants show multiple developmental alterations indicating opposite functions of cytokinins in the regulation of shoot and root meristem activity. Plant Cell 2003, 15, 2532–2550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyawaki, K.; Tarkowski, P.; Matsumoto-Kitano, M.; Kato, T.; Sato, S.; Tarkowska, D.; Tabata, S.; Sandberg, G.; Kakimoto, T. Roles of Arabidopsis ATP/ADP isopentenyltransferases and tRNA isopentenyltransferases in cytokinin biosynthesis. Proc. Natl. Acad. Sci. USA 2006, 103, 16598–16603. [Google Scholar] [CrossRef] [Green Version]
- Schmitz, R.Y.; Skoog, F.; Playtis, A.J.; Leonard, N.J. Cytokinins: Synthesis and biological activity of geometric and position isomers of zeatin. Plant Physiol. 1972, 50, 702–705. [Google Scholar] [CrossRef]
- Kamínek, M.; Pačes, V.; Corse, J.; Challice, J.S. Effect of stereospecific hydroxylation of N6-(Δ2-isopentenyl)adenosine on cytokinin activity. Planta 1979, 145, 239–243. [Google Scholar] [CrossRef]
- Gajdošová, S.; Spíchal, L.; Kamínek, M.; Hoyerová, K.; Novák, O.; Dobrev, P.I.; Galuszka, P.; Klíma, P.; Gaudinová, A.; Žižková, E.; et al. Distribution, biological activities, metabolism, and the conceivable function of cis-zeatin-type cytokinins in plants. J. Exp. Bot. 2011, 62, 2827–2840. [Google Scholar] [CrossRef] [Green Version]
- Kudo, T.; Makita, N.; Kojima, M.; Tokunaga, H.; Sakakibara, H. Cytokinin Activity of cis-Zeatin and Phenotypic Alterations Induced by Overexpression of Putative cis-Zeatin-O-glucosyltransferase in Rice. Plant Physiol. 2012, 160, 319–331. [Google Scholar] [CrossRef] [Green Version]
- Stirk, W.A.; Václavíková, K.; Novák, O.; Gajdošová, S.; Kotland, O.; Motyka, V.; Strnad, M.; van Staden, J. Involvement of cis-zeatin, dihydrozeatin, and aromatic cytokinins in germination and seedling establishment of maize, oats, and lucerne. J. Plant Growth Regul. 2012, 31, 392–405. [Google Scholar] [CrossRef]
- Schäfer, M.; Brütting, C.; Meza-Canales, I.D.; Grosskinsky, D.K.; Vankova, R.; Baldwin, I.T.; Meldau, S. The role of cis-zeatin-type cytokinins in plant growth regulation and mediating responses to environmental interactions. J. Exp. Bot. 2015, 66, 4873–4884. [Google Scholar] [CrossRef] [Green Version]
- Ružić, D.V.; Vujović, T.I. The effects of cytokinin types and their concentration on in vitro multiplication of sweet cherry cv. Lapins (Prunus avium L.). Hortic. Sci. 2008, 35, 12–21. [Google Scholar] [CrossRef] [Green Version]
- Moura, M.; Candeias, M.I.; Silva, L. In vitro propagation of Viburnum treleasei Gand., an Azorean endemic with high ornamental interest. HortScience 2009, 44, 1668–1671. [Google Scholar] [CrossRef] [Green Version]
- Yunus, M.F.; Aziz, M.A.; Kadir, M.A.; Rashid, A.A. In vitro propagation of Etlingera elatior (Jack) (torch ginger). Sci. Hortic. 2012, 135, 145–150. [Google Scholar] [CrossRef]
- Kulus, D. Influence of growth regulators on the development, quality, and physiological state of in vitro-propagated Lamprocapnos spectabilis (L.) Fukuhara. Vitr. Cell. Dev. Biol. Plant 2020, 56, 447–457. [Google Scholar] [CrossRef]
- Dang, S.; Gao, R.; Zhang, Y.; Feng, Y. In vitro regeneration and its histological characteristics of Dioscorea nipponica Makino. Sci. Rep. 2022, 12, 18436. [Google Scholar] [CrossRef]
- Lomin, S.N.; Krivosheev, D.M.; Steklov, M.Y.; Arkhipov, D.V.; Osolodkin, D.I.; Schmülling, T.; Romanov, G.A. Plant membrane assays with cytokinin receptors underpin the unique role of free cytokinin bases as biologically active ligands. J. Exp. Bot. 2015, 66, 1851–1863. [Google Scholar] [CrossRef] [Green Version]
- Trifunović-Momčilov, M.; Motyka, V.; Dragićević, I.Č.; Petrić, M.; Jevremović, S.; Malbeck, J.; Holík, J.; Dobrev, P.I.; Subotić, A. Endogenous phytohormones in spontaneously regenerated Centaurium erythraea Rafn. plants grown in vitro. J. Plant Growth Regul. 2016, 35, 543–552. [Google Scholar] [CrossRef]
- Raspor, M.; Motyka, V.; Ninković, S.; Dobrev, P.I.; Malbeck, J.; Ćosić, T.; Cingel, A.; Savić, J.; Tadić, V.; Dragićević, I.Č. Endogenous levels of cytokinins, indole-3-acetic acid and abscisic acid in in vitro grown potato: A contribution to potato hormonomics. Sci. Rep. 2020, 10, 3437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, B.; Guo, X.; Wang, Y.; Xiong, Y.; Wang, J.; Hayashi, K.I.; Lei, J.; Zhang, L.; Jiao, Y. Feedback from lateral organs controls shoot apical meristem growth by modulating auxin transport. Dev. Cell 2018, 44, 204–216. [Google Scholar] [CrossRef] [Green Version]
- Scarpella, E.; Barkoulas, M.; Tsiantis, M. Control of leaf and vein development by auxin. Cold Spring Harb. Perspect. Biol. 2010, 2, a001511. [Google Scholar] [CrossRef] [Green Version]
- Byrne, M.E. Making leaves. Curr. Opin. Plant Biol. 2012, 15, 24–30. [Google Scholar] [CrossRef]
- Peer, W.A.; Cheng, Y.; Murphy, A.S. Evidence of oxidative attenuation of auxin signalling. J. Exp. Bot. 2013, 64, 2629–2639. [Google Scholar] [CrossRef]
- Novák, O.; Hényková, E.; Sairanen, I.; Kowalczyk, M.; Pospíšil, T.; Ljung, K. Tissue-specific profiling of the Arabidopsis thaliana auxin metabolome. Plant J. 2012, 72, 523–536. [Google Scholar] [CrossRef]
- Kasahara, H. Current aspects of auxin biosynthesis in plants. Biosci. Biotechnol. Biochem. 2016, 80, 34–42. [Google Scholar] [CrossRef]
- Nemoto, K.; Hara, M.; Suzuki, M.; Seki, H.; Muranaka, T.; Mano, Y. The NtAMI1 gene functions in cell division of tobacco BY-2 cells in the presence of indole-3-acetamide. FEBS Lett. 2009, 583, 487–492. [Google Scholar] [CrossRef] [Green Version]
- Ljung, K. Auxin metabolism and homeostasis during plant development. Development 2013, 140, 943–950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiong, L.; Zhu, J.K. Regulation of abscisic acid biosynthesis. Plant Physiol. 2003, 133, 29–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tuteja, N. Abscisic acid and abiotic stress signalling. Plant Signal. Behav. 2007, 2, 135–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, Y.; Borrego, E.; Kolomiets, M.V. Jasmonate biosynthesis, perception and function in plant development and stress responses. In Lipid Metabolism; Baez, R.V., Ed.; IntechOpen: London, UK, 2013; pp. 393–442. ISBN 978-953-51-0944-0. [Google Scholar] [CrossRef] [Green Version]
- Cuesta, C.; Novák, O.; Ordás, R.J.; Fernández, B.; Strnad, M.; Doležal, K.; Rodríguez, A. Endogenous cytokinin profiles and their relationships to between-family differences during adventitious caulogenesis in Pinus pinea cotyledons. J. Plant Physiol. 2012, 169, 1830–1837. [Google Scholar] [CrossRef] [PubMed]
- Kamínek, M.; Motyka, V.; Vaňková, R. Regulation of cytokinin content in plant cells. Physiol. Plant. 1997, 101, 689–700. [Google Scholar] [CrossRef]
- Klemš, M.; Slámová, Z.; Motyka, V.; Malbeck, J.; Trávníčková, A.; Macháčková, I.; Holík, J.; Procházka, S. Changes in cytokinin levels and metabolism in tobacco (Nicotiana tabacum L.) explants during in vitro shoot organogenesis induced by trans-zeatin and dihydrozeatin. Plant Growth Regul. 2011, 65, 427–437. [Google Scholar] [CrossRef]
- Montalbán, I.A.; Novák, O.; Rolčik, J.; Strnad, M.; Moncaleán, P. Endogenous cytokinin and auxin profiles during in vitro organogenesis from vegetative buds of Pinus radiata adult trees. Physiol. Plant. 2013, 148, 214–231. [Google Scholar] [CrossRef]
- Aremu, A.O.; Plačková, L.; Bairu, M.W.; Novák, O.; Plíhalová, L.; Doležal, K.; Finnie, J.F.; Van Staden, J. How does exogenously applied cytokinin type affect growth and endogenous cytokinins in micropropagated Merwilla plumbea? Plant Cell Tiss. Organ Cult. 2014, 118, 245–256. [Google Scholar] [CrossRef]
- Bassil, N.V.; Mok, D.W.S.; Mok, M.C. Partial purification of a cis-trans-isomerase of zeatin from immature seed of Phaseolus vulgaris L. Plant Physiol. 1993, 102, 867–872. [Google Scholar] [CrossRef] [Green Version]
- Kasahara, H.; Takei, K.; Ueda, N.; Hishiyama, S.; Yamaya, T.; Kamiya, Y.; Yamaguchi, S.; Sakakibara, H. Distinct isoprenoid origins of cis- and trans-zeatin biosyntheses in Arabidopsis. J. Biol. Chem. 2004, 279, 14049–14054. [Google Scholar] [CrossRef] [Green Version]
- Altmann, M.; Altmann, S.; Rodriguez, P.A.; Weller, B.; Vergara, L.E.; Palme, J.; Marín-de la Rosa, N.; Sauer, M.; Wenig, M.; Villaécija-Aguilar, J.A.; et al. Extensive signal integration by the phytohormone protein network. Nature 2020, 583, 271–276. [Google Scholar] [CrossRef]
- Romanov, G.A.; Lomin, S.N.; Schmülling, T. Biochemical characteristics and ligand-binding properties of Arabidopsis cytokinin receptor AHK3 compared to CRE1/AHK4 as revealed by a direct binding assay. J. Exp. Bot. 2006, 57, 4051–4058. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.G.; Li, W.; Mao, Y.F.; Zhao, D.L.; Dong, W.; Guo, G.Q. Endogenous hormonal levels in Scutellaria baicalensis calli induced by thidiazuron. Russ. J. Plant Physiol. 2005, 52, 345–351. [Google Scholar] [CrossRef]
- Eklöf, S.; Ȧstot, C.; Blackwell, J.; Moritz, T.; Olsson, O.; Sandberg, G. Auxin-cytokinin interactions in wild-type and transgenic tobacco. Plant Cell Physiol. 1997, 38, 225–235. [Google Scholar] [CrossRef] [Green Version]
- Cheng, Z.J.; Wang, L.; Sun, W.; Zhang, Y.; Zhou, C.; Su, Y.H.; Li, W.; Sun, T.T.; Zhao, X.Y.; Li, X.G.; et al. Pattern of auxin and cytokinin responses for shoot meristem induction results from the regulation of cytokinin biosynthesis by AUXIN RESPONSE FACTOR3. Plant Physiol. 2013, 161, 240–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Che, P.; Lall, S.; Nettleton, D.; Howell, S.H. Gene expression programs during shoot, root, and callus development in Arabidopsis tissue culture. Plant Physiol. 2006, 141, 620–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Che, P.; Lall, S.; Howell, S.H. Developmental steps in acquiring competence for shoot development in Arabidopsis tissue culture. Planta 2007, 226, 1183–1194. [Google Scholar] [CrossRef]
- Motte, H.; Vereecke, D.; Geelen, D.; Werbrouck, S. The molecular path to in vitro shoot regeneration. Biotechnol. Adv. 2014, 32, 107–121. [Google Scholar] [CrossRef]
- Benková, E.; Michniewicz, M.; Sauer, M.; Teichmann, T.; Seifertová, D.; Jürgens, G.; Friml, J. Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell 2003, 115, 591–602. [Google Scholar] [CrossRef] [Green Version]
- Pernisová, M.; Klíma, P.; Horák, J.; Válková, M.; Malbeck, J.; Souček, P.; Reichman, P.; Hoyerová, K.; Dubová, J.; Friml, J.; et al. Cytokinins modulate auxin-induced organogenesis in plants via regulation of the auxin efflux. Proc. Natl. Acad. Sci. USA 2009, 106, 3609–3614. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.Y.; Su, Y.H.; Zhang, C.L.; Wang, L.; Li, X.G.; Zhang, X.S. Differences in capacities of in vitro organ regeneration between two Arabidopsis ecotypes Wassilewskija and Columbia. Plant Cell Tiss. Organ Cult. 2013, 112, 65–74. [Google Scholar] [CrossRef]
- Kakani, A.; Li, G.; Peng, Z. Role of AUX1 in the control of organ identity during in vitro organogenesis and in mediating tissue specific auxin and cytokinin interaction in Arabidopsis. Planta 2009, 229, 645–657. [Google Scholar] [CrossRef] [PubMed]
- Koike, I.; Taniguchi, K.; Shimomura, K.; Umehara, M. Dynamics of endogenous indole-3-acetic acid and cytokinins during adventitious shoot formation in ipecac. J. Plant Growth Regul. 2017, 36, 805–813. [Google Scholar] [CrossRef]
- Salvi, E.; Di Mambro, R.; Pacifici, E.; Dello Ioio, R.; Costantino, P.; Moubayidin, L.; Sabatini, S. SCARECROW and SHORTROOT control the auxin/cytokinin balance necessary for embryonic stem cell niche specification. Plant Signal. Behav. 2018, 13, e1507402. [Google Scholar]
- Shao, D.; Ma, Y.; Li, X.; Ga, S.; Ren, Y. The sequence structure and phylogenetic analysis by complete mitochondrial genome of kohlrabi (Brassica oleracea var. gongylodes L.). Mitochondrial DNA B 2021, 6, 2714–2716. [Google Scholar] [CrossRef]
- Atta, R.; Laurens, L.; Boucheron-Dubuisson, E.; Guivarc’h, A.; Carnero, E.; Giraudat-Pautot, V.; Rech, P.; Chriqui, D. Pluripotency of Arabidopsis xylem pericycle underlies shoot regeneration from root and hypocotyl explants grown in vitro. Plant J. 2009, 57, 626–644. [Google Scholar] [CrossRef]
- Rosspopoff, O.; Chelysheva, L.; Saffar, J.; Lecorgne, L.; Gey, D.; Caillieux, E.; Colot, V.; Roudier, F.; Hilson, P.; Berthomé, R.; et al. Direct conversion of root primordium into shoot meristem relies on timing of stem cell niche development. Development 2017, 144, 1187–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berckmans, B.; De Veylder, L. Transcriptional control of the cell cycle. Curr. Opin. Plant Biol. 2009, 12, 599–605. [Google Scholar] [CrossRef]
- Dong, Q.; Zhao, Y.; Jiang, H.; He, H.; Zhu, S.; Cheng, B.; Xiang, Y. Genome-wide identification and characterization of the cyclin gene family in Populus trichocarpa. Plant Cell Tiss. Organ Cult. 2011, 107, 55–67. [Google Scholar] [CrossRef]
- Dewitte, W.; Murray, J.A.H. The plant cell cycle. Annu. Rev. Plant Biol 2003, 54, 235–264. [Google Scholar] [CrossRef] [Green Version]
- Nieuwland, J.; Maughan, S.; Dewitte, W.; Scofield, S.; Sanz, L.; Murray, J.A.H. The D-type cyclin CYCD4;1 modulates lateral root density in Arabidopsis by affecting the basal meristem region. Proc. Natl. Acad. Sci. USA 2009, 106, 22528–22533. [Google Scholar] [CrossRef] [Green Version]
- Barrôco, R.M.; Van Poucke, K.; Bergervoet, J.H.W.; De Veylder, L.; Groot, S.P.C.; Inzé, D.; Engler, G. The role of the cell cycle machinery in resumption of postembryonic development. Plant Physiol. 2005, 137, 127–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montero-Cortés, M.; Rodríguez-Paredes, F.; Burgeff, C.; Pérez-Nuñez, T.; Córdova, I.; Oropeza, C.; Verdeil, J.L.; Saénz, L. Characterisation of a cyclin-dependent kinase (CDKA) gene expressed during somatic embryogenesis of coconut palm. Plant Cell Tiss. Organ Cult. 2010, 102, 251–258. [Google Scholar] [CrossRef]
- Andersen, S.U.; Buechel, S.; Zhao, Z.; Ljung, K.; Novák, O.; Busch, W.; Schuster, C.; Lohmann, J.U. Requirement of B2-type Cyclin-Dependent Kinases for meristem integrity in Arabidopsis thaliana. Plant Cell 2008, 20, 88–100. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, C. Coupling cell proliferation and development in plants. Nat. Cell Biol. 2005, 7, 535–541. [Google Scholar] [CrossRef]
- Planchais, S.; Glab, N.; Tréhin, C.; Perennes, C.; Bureau, J.M.; Meijer, L.; Bergounioux, C. Roscovitine, a novel cyclin-dependent kinase inhibitor, characterizes restriction point and G2/M transition in tobacco BY-2 cell suspension. Plant J. 1997, 12, 191–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Redig, P.; Shaul, O.; Inzé, D.; Van Montagu, M.; Van Onckelen, H. Levels of endogenous cytokinins, indole-3-acetic acid and abscisic acid during the cell cycle of synchronized tobacco BY-2 cells. FEBS Lett. 1996, 391, 175–180. [Google Scholar] [CrossRef]
- Laureys, F.; Dewitte, W.; Witters, E.; Van Montagu, M.; Inzé, D.; Van Onckelen, H. Zeatin is indispensable for the G2-M transition in tobacco BY-2 cells. FEBS Lett. 1998, 426, 29–32. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Diederich, L.; John, P.C.L. The cytokinin requirement for cell division in cultured Nicotiana plumbaginifolia cells can be satisfied by yeast Cdc25 protein tyrosine phosphatase. Implications for mechanisms of cytokinin response and plant development. Plant Physiol. 2005, 137, 308–316. [Google Scholar] [CrossRef] [Green Version]
- Schaller, G.E.; Street, I.H.; Kieber, J.J. Cytokinin and the cell cycle. Curr. Opin. Plant Biol. 2014, 21, 7–15. [Google Scholar] [CrossRef]
- Duclercq, J.; Sangwan-Norreel, B.; Catterou, M.; Sangwan, R.S. De novo shoot organogenesis: From art to science. Trends Plant Sci. 2011, 16, 597–606. [Google Scholar] [CrossRef]
- Mishra, B.S.; Sharma, M.; Laxmi, A. Role of sugar and auxin crosstalk in plant growth and development. Physiol. Plant. 2022, 174, e13546. [Google Scholar] [CrossRef]
- Hara, M.; Oki, K.; Hoshino, K.; Kuboi, T. Enhancement of anthocyanin biosynthesis by sugar in radish (Raphanus sativus) hypocotyl. Plant Sci. 2003, 164, 259–265. [Google Scholar] [CrossRef]
- Song, X.; Guo, H.; Liu, Y.; Wan, F.; Zhang, J.; Chang, X. Effects of salicylic acid and sucrose on pigment content in Pistacia chinensis leaves. Sci. Hortic. 2020, 259, 108783. [Google Scholar] [CrossRef]
- Solfanelli, C.; Poggi, A.; Loreti, E.; Alpi, A.; Perata, P. Sucrose-specific induction of the anthocyanin biosynthetic pathway in Arabidopsis. Plant Physiol. 2006, 140, 637–646. [Google Scholar] [CrossRef] [Green Version]
- Wasternack, C.; Strnad, M. Jasmonates are signals in the biosynthesis of secondary metabolites—Pathways, transcription factors and applied aspects—A brief review. New Biotechnol. 2019, 48, 1–11. [Google Scholar] [CrossRef]
- LeClere, S.; Schmelz, E.A.; Chourey, P.S. Sugar levels regulate tryptophan-dependent auxin biosynthesis in developing maize kernels. Plant Physiol. 2010, 153, 306–318. [Google Scholar] [CrossRef] [Green Version]
- Sakr, S.; Wang, M.; Dédaldéchamp, F.; Perez-Garcia, M.D.; Ogé, L.; Hamama, L.; Atanassova, R. The sugar-signaling hub: Overview of regulators and interaction with the hormonal and metabolic network. Int. J. Mol. Sci. 2018, 19, 2506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meitzel, T.; Radchuk, R.; McAdam, E.L.; Thormählen, I.; Feil, R.; Munz, E.; Hilo, A.; Geigenberger, P.; Ross, J.J.; Lunn, J.E.; et al. Trehalose 6-phosphate promotes seed filling by activating auxin biosynthesis. New Phytol. 2021, 229, 1553–1565. [Google Scholar] [CrossRef] [PubMed]
- Sairanen, I.; Novák, O.; Pěnčík, A.; Ikeda, Y.; Jones, B.; Sandberg, G.; Ljung, K. Soluble Carbohydrates Regulate Auxin Biosynthesis via PIF Proteins in Arabidopsis. Plant Cell 2012, 24, 4907–4916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McAdam, E.L.; Meitzel, T.; Quittenden, L.J.; Davidson, S.E.; Dalmais, M.; Bendahmane, A.I.; Thompson, R.; Smith, J.J.; Nichols, D.S.; Urquhart, S.; et al. Evidence that auxin is required for normal seed size and starch synthesis in pea. New Phytol. 2017, 216, 193–204. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.H.; Endo, A.; Zhou, L.; Penney, J.; Chen, H.C.; Arroyo, A.; Leon, P.; Nambara, E.; Asami, T.; Seo, M.; et al. A unique short-chain dehydrogenase/reductase in Arabidopsis glucose signaling and abscisic acid biosynthesis and functions. Plant Cell 2002, 14, 2723–2743. [Google Scholar] [CrossRef] [PubMed]
- Seo, M.; Koshiba, T. Complex regulation of ABA biosynthesis in plants. Trends Plant Sci. 2002, 7, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S.; Kamiya, Y. Gibberellin biosynthesis: Its regulation by endogenous and environmental signals. Plant Cell Physiol. 2000, 41, 251–257. [Google Scholar] [CrossRef] [Green Version]
- Gordon, S.P.; Heisler, M.G.; Reddy, G.V.; Ohno, C.; Das, P.; Meyerowitz, E.M. Pattern formation during de novo assembly of the Arabidopsis shoot meristem. Development 2007, 134, 3539–3548. [Google Scholar] [CrossRef] [Green Version]
- Huang, W.L.; Lee, C.H.; Chen, Y.R. Levels of endogenous abscisic acid and indole-3-acetic acid influence shoot organogenesis in callus cultures of rice subjected to osmotic stress. Plant Cell Tiss. Organ Cult. 2012, 108, 257–263. [Google Scholar] [CrossRef]
- Hwang, I.; Sheen, J. Two-component circuitry in Arabidopsis cytokinin signal transduction. Nature 2001, 413, 383–389. [Google Scholar] [CrossRef]
- Kushwah, S.; Laxmi, A. The interaction between glucose and cytokinin signal transduction pathway in Arabidopsis thaliana. Plant Cell Environ. 2014, 37, 235–253. [Google Scholar] [CrossRef]
- Kiba, T.; Takebayashi, Y.; Kojima, M.; Sakakibara, H. Sugar-induced de novo cytokinin biosynthesis contributes to Arabidopsis growth under elevated CO2. Sci. Rep. 2019, 9, 7765. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Le Gourrierec, J.; Jiao, F.; Demotes-Mainard, S.; Perez-Garcia, M.D.; Ogé, L.; Hamama, L.; Crespel, L.; Bertheloot, J.; Chen, J.; et al. Convergence and divergence of sugar and cytokinin signaling in plant development. Int. J. Mol. Sci. 2021, 22, 1282. [Google Scholar] [CrossRef]
- Barbier, F.; Péron, T.; Lecerf, M.; Perez-Garcia, M.D.; Barrière, Q.; Rolčík, J.; Boutet-Mercey, S.; Citerne, S.; Lemoine, R.; Porcheron, B.; et al. Sucrose is an early modulator of the key hormonal mechanisms controlling bud outgrowth in Rosa hybrida. J. Exp. Bot. 2015, 66, 2569–2582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kushwah, S.; Laxmi, A. The interaction between glucose and cytokinin signaling in controlling Arabidopsis thaliana seedling root growth and development. Plant Signal. Behav. 2017, 12, e1312241. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ćosić, T.; Raspor, M.; Motyka, V.; Cingel, A.; Ninković, S. In Vitro Growth and Regeneration of Brassica oleracea var. gongylodes: A Decade of Research. Horticulturae 2023, 9, 674. https://doi.org/10.3390/horticulturae9060674
Ćosić T, Raspor M, Motyka V, Cingel A, Ninković S. In Vitro Growth and Regeneration of Brassica oleracea var. gongylodes: A Decade of Research. Horticulturae. 2023; 9(6):674. https://doi.org/10.3390/horticulturae9060674
Chicago/Turabian StyleĆosić, Tatjana, Martin Raspor, Václav Motyka, Aleksandar Cingel, and Slavica Ninković. 2023. "In Vitro Growth and Regeneration of Brassica oleracea var. gongylodes: A Decade of Research" Horticulturae 9, no. 6: 674. https://doi.org/10.3390/horticulturae9060674