
Analysis of YUC and TAA/TAR Gene Families in Tomato

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Materials
2.2. Experimental Methods
2.2.1. Identification and Naming of Tomato YUC and TAA Family Members
2.2.2. Sequence Analysis of Tomato YUC and TAA Genes
2.2.3. Extraction and Purification of Total RNA from Various Tissues of Tomato
2.2.4. Preparation of cDNA from Various Tissues of Tomato
2.2.5. Analysis of Gene Expression in Various Tissues of Tomato
3. Results
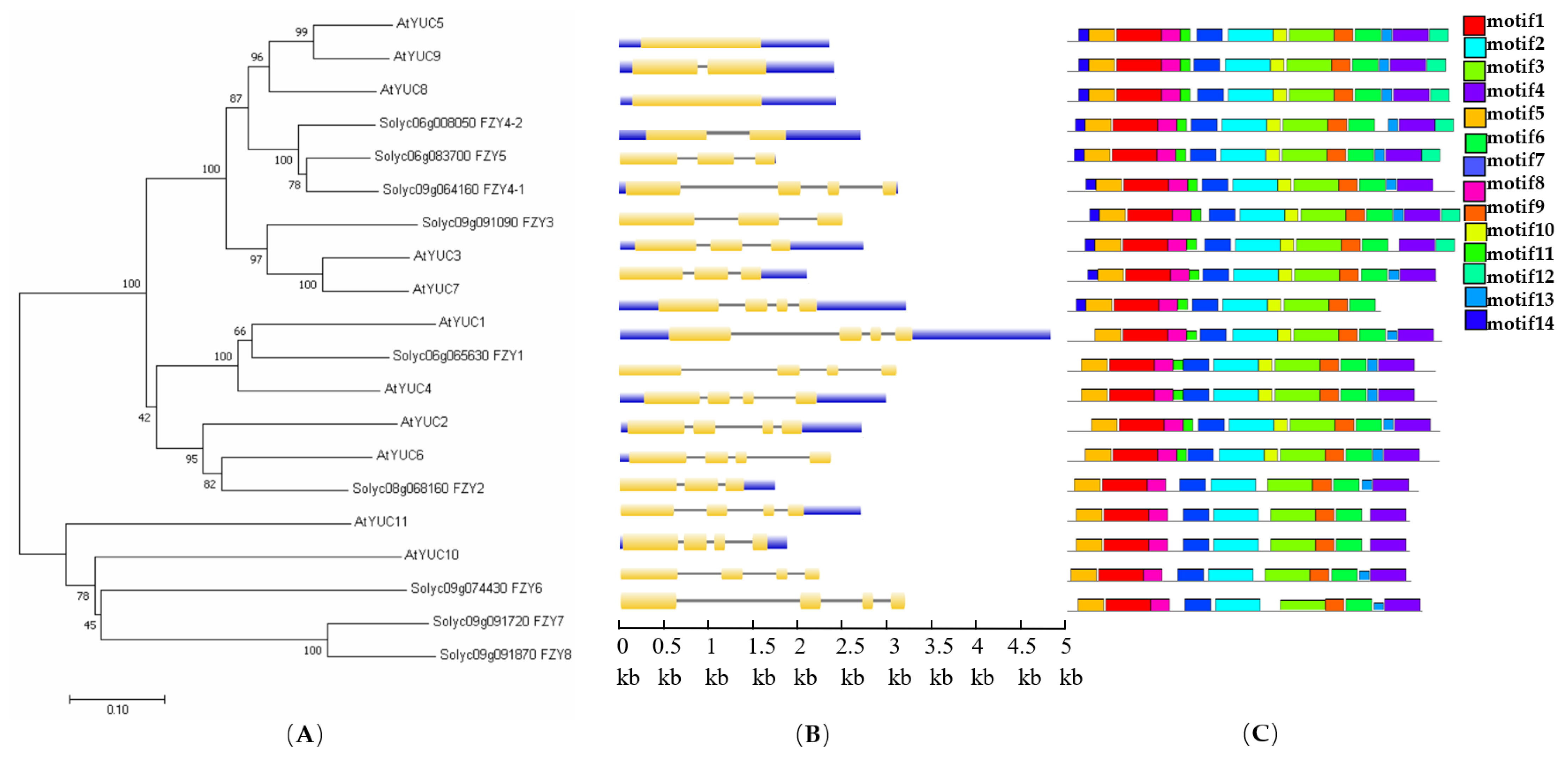
3.1. Searching for and Naming of YUC/FZY Genes in Tomato
3.2. Characteristic Analysis of YUC/FZY Genes in Tomato
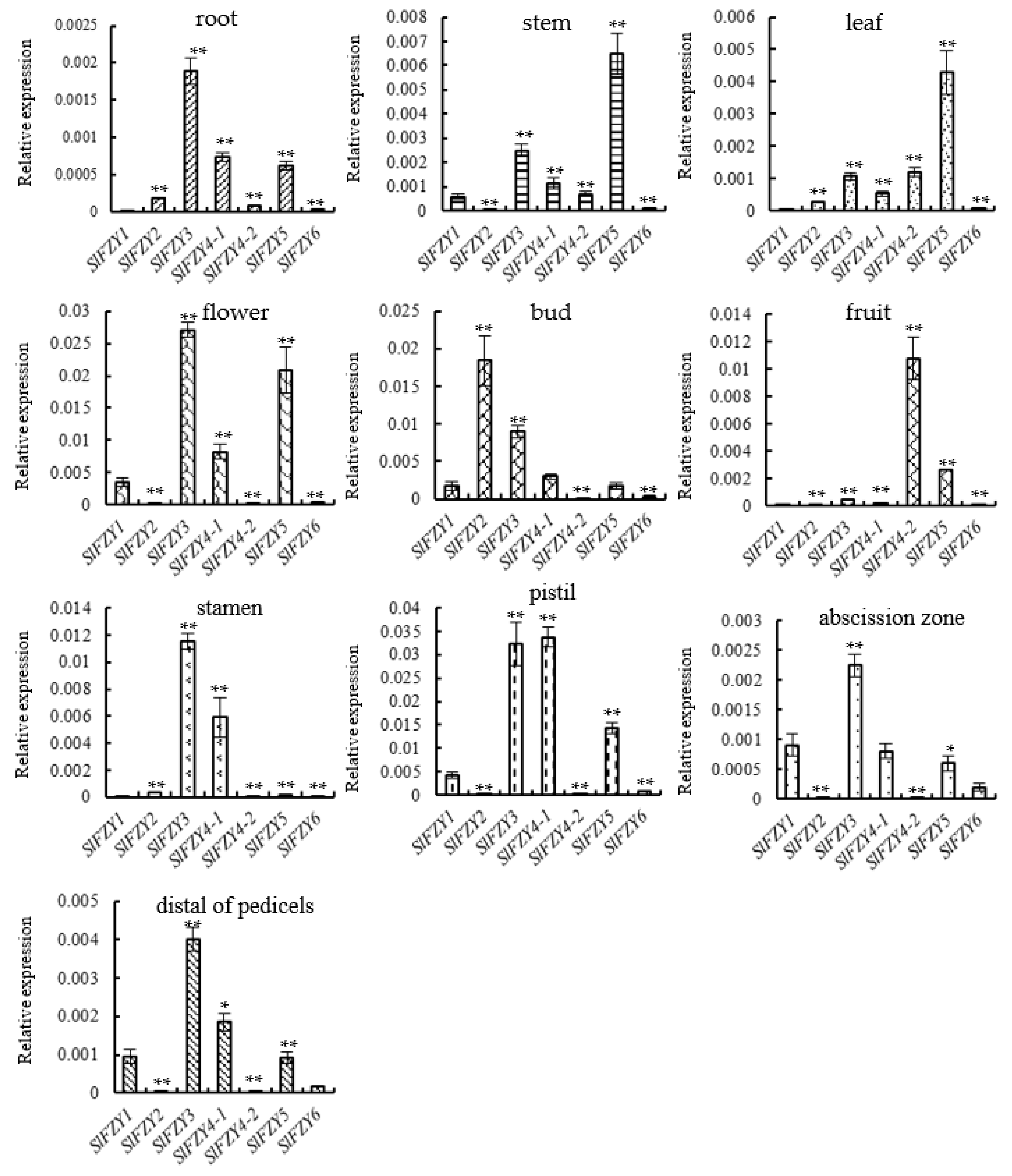
3.3. Analysis of Expression Patterns of YUC/FZY Genes in Various Tissues of Tomato
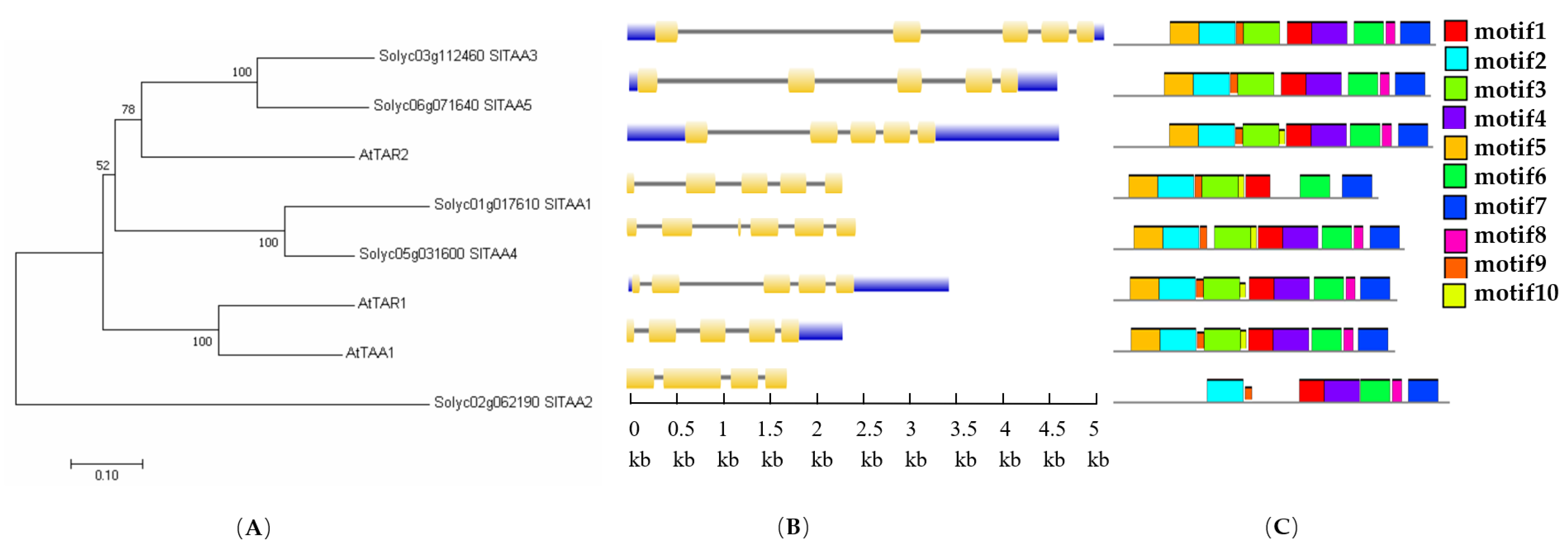
3.4. Identification and Structure Analysis of the Genes Encoding TAA Transaminase in Tomato
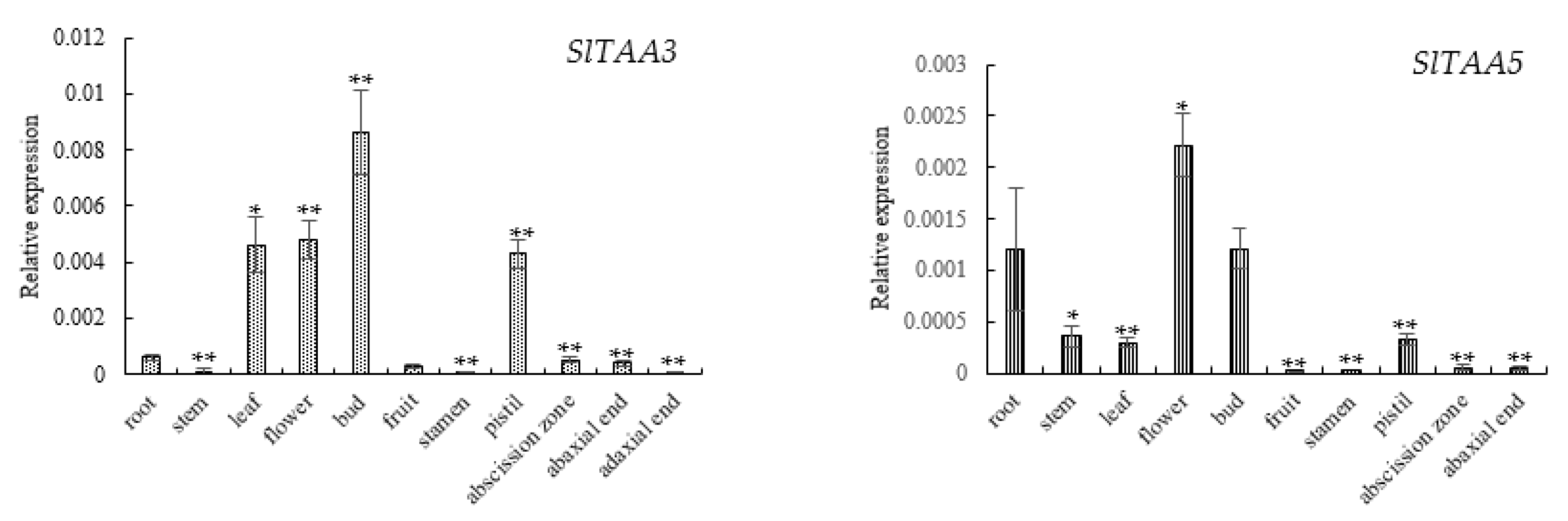
3.5. Expression Analysis of TAA Genes in Various Tissues of Tomato
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lincoln, C.; Britton, J.H.; Estelle, M. Growth and Development of the axr1 Mutants of Arabidopsis. Plant Cell 1990, 2, 1071–1080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collett, C.E.; Harberd, N.P.; Leyser, O. Hormonal Interactions in the Control of Arabidopsis Hypocotyl Elongation. Plant Physiol. 2000, 124, 553–562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, R.S.; Guyomarc’H, S.; Mandel, T.; Reinhardt, D.; Kuhlemeier, C.; Prusinkiewicz, P. A plausible model of phyllotaxis. Proc. Natl. Acad. Sci. USA 2006, 103, 1301–1306. [Google Scholar] [CrossRef] [Green Version]
- Qi, J.; Wang, Y.; Yu, T.; Cunha, A.; Wu, B.; Vernoux, T.; Meyerowitz, E.; Jiao, Y. Auxin depletion from leaf primordia contributes to organ patterning. Proc. Natl. Acad. Sci. USA 2014, 111, 18769–18774. [Google Scholar] [CrossRef] [Green Version]
- Stepanova, A.N.; Robertson-Hoyt, J.; Yun, J.; Benavente, L.M.; Xie, D.-Y.; Doležal, K.; Schlereth, A.; Jürgens, G.; Alonso, J.M. TAA1-Mediated Auxin Biosynthesis Is Essential for Hormone Crosstalk and Plant Development. Cell 2008, 133, 177–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, T.; Chen, K.; Dong, N.; Ye, W.; Shan, J.; Lin, H. Tillering and small grain 1 dominates the tryptophan aminotransferase family required for local auxin biosynthesis in rice. J. Integr. Plant Biol. 2019, 62, 581–600. [Google Scholar] [CrossRef] [PubMed]
- Taylor, J.E.; Whitelaw, C.A. Signals in abscission. New Phytol. 2001, 151, 323–340. [Google Scholar] [CrossRef]
- Cheng, Y.; Dai, X.; Zhao, Y. Auxin biosynthesis by the YUCCA flavin monooxygenases controls the formation of floral organs and vascular tissues in Arabidopsis. Genes Dev. 2006, 20, 1790–1799. [Google Scholar] [CrossRef] [Green Version]
- Cecchetti, V.; Celebrin, D.; Napoli, N.; Ghelli, R.; Brunetti, P.; Costantino, P.; Cardarelli, M. An auxin maximum in the middle layer controls stamen development and pollen maturation in Arabidopsis. New Phytol. 2016, 213, 1194–1207. [Google Scholar] [CrossRef] [Green Version]
- Poulet, A.; Kriechbaumer, V. Bioinformatics Analysis of Phylogeny and Transcription of TAA/YUC Auxin Biosynthetic Genes. Int. J. Mol. Sci. 2017, 18, 1791. [Google Scholar] [CrossRef] [Green Version]
- Qin, H.; Zhang, Z.; Wang, J.; Chen, X.; Wei, P.; Huang, R. The activation of OsEIL1 on YUC8 transcription and auxin biosynthesis is required for ethylene-inhibited root elongation in rice early seedling development. PLoS Genet. 2017, 13, e1006955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Q.; Dai, X.; De-Paoli, H.; Cheng, Y.; Takebayashi, Y.; Kasahara, H.; Kamiya, Y.; Zhao, Y. Auxin Overproduction in Shoots Cannot Rescue Auxin Deficiencies in Arabidopsis Roots. Plant Cell Physiol. 2014, 55, 1072–1079. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y. The role of local biosynthesis of auxin and cytokinin in plant development. Curr. Opin. Plant Biol. 2008, 11, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Poulios, S.; Vlachonasios, K.E. Synergistic action of GCN5 and CLAVATA1 in the regulation of gynoecium development in Arabidopsis thaliana. New Phytol. 2018, 220, 593–608. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Prunet, N.; Gan, E.-S.; Wang, Y.; Stewart, D.; Wellmer, F.; Huang, J.; Yamaguchi, N.; Tatsumi, Y.; Kojima, M.; et al. SUPERMAN regulates floral whorl boundaries through control of auxin biosynthesis. EMBO J. 2018, 37, e97499. [Google Scholar] [CrossRef]
- Yuan, H.; Zhao, K.; Lei, H.; Shen, X.; Liu, Y.; Liao, X.; Li, T. Genome-wide analysis of the GH3 family in apple (Malus × domestica). BMC Genom. 2013, 14, 297. [Google Scholar] [CrossRef] [Green Version]
- Stepanova, A.N.; Yun, J.; Robles, L.M.; Novak, O.; He, W.; Guo, H.; Ljung, K.; Alonso, J.M. The Arabidopsis YUCCA1 flavin monooxygenase functions in the indole-3-pyruvic acid branch of auxin biosynthesis. Plant Cell 2011, 23, 3961–3973. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Mashiguchi, K.; Chen, Q.; Kasahara, H.; Kamiya, Y.; Ojha, S.; DuBois, J.; Ballou, D.; Zhao, Y. The biochemical mechanism of auxin biosynthesis by an arabidopsis YUCCA flavin-containing monooxygenase. J. Biol. Chem. 2013, 288, 1448–1457. [Google Scholar] [CrossRef] [Green Version]
- Zhao, C.; Wang, P.; Si, T.; Hsu, C.-C.; Wang, L.; Zayed, O.; Yu, Z.; Zhu, Y.; Dong, J.; Tao, W.A.; et al. MAP Kinase Cascades Regulate the Cold Response by Modulating ICE1 Protein Stability. Dev. Cell 2017, 43, 618–629. [Google Scholar] [CrossRef] [Green Version]
- Mashiguchi, K.; Tanaka, K.; Sakai, T.; Sugawara, S.; Kawaide, H.; Natsume, M.; Hanada, A.; Yaeno, T.; Shirasu, K.; Yao, H.; et al. The main auxin biosynthesis pathway in Arabidopsis. Proc. Natl. Acad. Sci. USA 2011, 108, 18512–18517. [Google Scholar] [CrossRef] [Green Version]
- Gao, Y.; Dai, X.; Zheng, Z.; Kasahara, H.; Kamiya, Y. Over expression of the bacterial tryptophan oxidase RebO affects auxin biosynthesis and Arabidopsis development. Sci. Bull. 2016, 61, 859–867. [Google Scholar] [CrossRef]
- Zhang, J.; Lin, J.E.; Harris, C.; Pereira, F.C.M.; Wu, F.; Blakeslee, J.J.; Peer, W.A. DAO1 catalyzes temporal and tissue-specific oxidative inactivation of auxin in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2016, 113, 11010–11015. [Google Scholar] [CrossRef] [Green Version]
- Ma, Q.; Dai, X.; Xu, Y.; Guo, J.; Liu, Y.; Chen, N.; Xiao, J.; Zhang, D.; Xu, Z.; Zhang, X.; et al. Enhanced tolerance to chilling stress in OsMYB3R-2 transgenic rice is mediated by alteration in cell cycle and ectopic expression of stress genes. Plant Physiol. 2009, 150, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Dai, X.; Zhao, Y. Auxin Synthesized by the YUCCA Flavin Monooxygenases Is Essential for Embryogenesis and Leaf Formation in Arabidopsis. Plant Cell 2007, 19, 2430–2439. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Liu, H.; Xiong, L. Endogenous auxin and jasmonic acid levels are differentially modulated by abiotic stresses in rice. Front. Plant Sci. 2013, 4, 397. [Google Scholar] [CrossRef] [Green Version]
- Koini, M.A.; Alvey, L.; Allen, T.; Tilley, C.A.; Harberd, N.P.; Whitelam, G.C.; Franklin, K.A. High Temperature-Mediated Adaptations in Plant Architecture Require the bHLH Transcription Factor PIF4. Curr. Biol. 2009, 19, 408–413. [Google Scholar] [CrossRef] [Green Version]
- Chen, L.; Tong, J.; Xiao, L.; Ruan, Y.; Liu, J.; Zeng, M.; Huang, H.; Wang, J.-W.; Xu, L. YUCCA-mediated auxin biogenesis is required for cell fate transition occurring during de novo root organogenesis in Arabidopsis. J. Exp. Bot. 2016, 67, 4273–4284. [Google Scholar] [CrossRef] [Green Version]
- Pucciariello, O.; Legris, M.; Rojas, C.C.; Iglesias, M.J.; Hernando, C.E.; Dezar, C.; Vazquez, M.; Yanovsky, M.J.; Finlayson, S.A.; Prat, S.; et al. Rewiring of auxin signaling under persistent shade. Proc. Natl. Acad. Sci. USA 2018, 115, 5612–5617. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y. Essential Roles of Local Auxin Biosynthesis in Plant Development and in Adaptation to Environmental Changes. Annu. Rev. Plant Biol. 2018, 69, 417–435. [Google Scholar] [CrossRef]
- Zhao, Y.; Christensen, S.K.; Fankhauser, C.; Cashman, J.R.; Cohen, J.D.; Weigel, D.; Chory, J. A role for flavin monooxygenase-like enzymes in auxin biosynthesis. Science 2001, 291, 306–309. [Google Scholar] [CrossRef] [PubMed]
- Cao, X.; Yang, H.; Shang, C.; Ma, S.; Liu, L.; Cheng, J. The Roles of Auxin Biosynthesis YUCCA Gene Family in Plants. Int. J. Mol. Sci. 2019, 20, 6343. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Family | Gene Name | Serial Number | Subcellular Location |
|---|---|---|---|
| YUCCA | SlFZY1 | Solyc06g065630 | Cytoplasmic |
| SlFZY2 | Solyc08g068160 | Mitochondrial | |
| SlFZY3 | Solyc09g091090 | Cytoplasmic | |
| SlFZY4-1 | Solyc09g064160 | Cytoplasmic | |
| SlFZY4-2 | Solyc06g008050 | Cytoplasmic | |
| SlFZY5 | Solyc06g083700 | Cytoplasmic/Mitochondrial | |
| SlFZY6 | Solyc09g074430 | Cytoplasmic | |
| SlFZY7 | Solyc09g091720 | Cytoplasmic/Plasma membrane | |
| SlFZY8 | Solyc09g091870 | Cytoplasmic |
| Gene | Regulatory Components | Number | Sequence | Function |
|---|---|---|---|---|
| SlFZY1 | ATC-motif | 1 | AGTAATCT | part of a conserved DNA module involved in light responsiveness |
| Box 4 | 7 | ATTAAT | part of a conserved DNA module involved in light responsiveness | |
| CAAT-box | 33 | CAAT/CAAAT | common cis-acting element in promoter and enhancer regions | |
| G-box | 1 | CACGAC | cis-acting regulatory element involved in light responsiveness | |
| GA-motif | 1 | ATAGATAA | part of a light-responsive element | |
| GATA-motif | 1 | AAGGATAAGG | part of a light-responsive element | |
| GT1-motif | 3 | GGTTAA | light-responsive element | |
| LAMP-element | 1 | CTTTATCA | part of a light-responsive element | |
| TATA-box | 26 | TATA/TATAA/TATAAGAA/TATAAATA/TATATTTATATTT | core promoter element around -30 of transcription start | |
| TC-rich repeats | 1 | GTTTTCTTAC | cis-acting element involved in defense and stress responsiveness | |
| SlFZY2 | AAGAA-motif | 1 | GAAAGAA | cis-acting element involved in abscisic acid responsiveness |
| ABRE | 1 | ACGTG | cis-acting element involved in abscisic acid responsiveness | |
| ARE | 1 | AAACCA | cis-acting regulatory element essential for anaerobic induction | |
| TCA-element | 1 | TCAGAAGAGG | cis-acting element involved in salicylic acid responsiveness | |
| TCCC-motif | 1 | TCTCCCT | part of a light-responsive element | |
| TCT-motif | 1 | TCTTAC | part of a light-responsive element | |
| TGACG-motif | 1 | TGACG | cis-acting regulatory element involved in MeJA responsiveness | |
| I-box | 1 | AGATAAGG | part of a light-responsive element | |
| MBS | 1 | CAACTG | MYB binding site involved in drought inducibility | |
| P-box | 1 | CCTTTTG | gibberellin-responsive element | |
| CAT-box | 1 | GCCACT | cis-acting regulatory element related to meristem expression | |
| CGTCA-motif | 1 | CGTCA | cis-acting regulatory element involved in MeJA responsiveness | |
| G-box | 1 | CACGTC | ||
| GA-motif | 1 | ATAGATAA | ||
| GT1-motif | 1 | GGTTAA | ||
| Box 4 | 3 | ATTAAT | ||
| TATA-box | 83 | TATATA/TATA/ATT ATA/ATATAT/TATA CA/TATAAAA | ||
| SlFZY3 | A-box | 1 | CCGTCC | cis-acting regulatory element |
| O2-site | 1 | GTTGACGTGA | cis-acting regulatory element involved in zein metabolism | |
| AuxRR-core | 1 | GGTCCAT | cis-acting regulatory element involved in auxin responsiveness | |
| TATA-box | 42 | TATA/ATATAT/TATAA ATA/TATAAA/TAT AA/TATAAAT | ||
| Box 4 | 8 | ATTAAT | ||
| CAAT-box | 43 | CAAAT/CAAT | ||
| TC-rich repeats | 1 | GTTTTCTTAC | ||
| G-box | 1 | CACGAC | ||
| GATA-motif | 1 | AAGATAAGATT | ||
| GT1-motif | 3 | GGTTAA/GGTTAAT | ||
| TCA-element | 1 | TCAGAAGAGG | ||
| CGTCA-motif | 1 | CGTCA | ||
| TCCC-motif | 1 | TCTCCCT | ||
| TGACG-motif | 1 | TGACG | ||
| SlFZY4-1 | Box 4 | 5 | ATTAAT | |
| CAAT-box | 45 | CCAAT/CAAT/ CAAAT | ||
| TATA-box | 65 | TATA/ATATAT/TATA AATA/TATAAA/T ATAA/TATAAAT | ||
| ARE | 1 | AAACCA | ||
| TCA-element | 1 | CCATCTTTTT | ||
| SlFZY4-2 | MRE | 1 | AACCTAA | MYB binding site involved in light responsiveness |
| ATCT-motif | 1 | AATCTAATCC | part of a conserved DNA module involved in light responsiveness | |
| MSA-like | 1 | TCAAACGGT | cis-acting element involved in cell cycle regulation | |
| LTR | 1 | CCGAAA | cis-acting element involved in low-temperature responsiveness | |
| CAAT-box | 32 | CAAAT/CAAT | ||
| LAMP-element | 1 | CTTTATCA | ||
| Box 4 | 4 | ATTAAT | ||
| G-Box | 3 | CACGTT | ||
| CGTCA-motif | 2 | CGTCA | ||
| TCA-element | 1 | TCAGAAGAGG | ||
| TGACG-motif | 2 | TGACG | ||
| I-box | 1 | ATGATAAGGTC | ||
| ARE | 1 | AAACCA | ||
| AuxRR-core | 1 | GGTCCAT | ||
| SlFZY5 | GARE-motif | 4 | TCTGTTG | gibberellin-responsive element |
| chs-CMA1a | 3 | TTACTTAA | part of a light-responsive element | |
| GCN4_motif | 1 | TGAGTCA | cis-regulatory element involved in endosperm expression | |
| GATA-motif | 2 | GATAGGG/GATAGG | ||
| G-box | 5 | GCCACGTGGA/CACGTG/CCACGTAA/TACGTG | ||
| GT1-motif | 7 | GGTTAAT/GGTTAA | ||
| Box 4 | 5 | ATTAAT | ||
| TATA-box | 77 | TACAAAA/ATATAT/TATA/TATAA/TATAAAA/TATAAA/TATATAA | ||
| CAAT-box | 48 | CAAT/CCAAT/CAACCAACTCC/CAAAT | ||
| TCA-element | 1 | CCATCTTTTT | ||
| TCT-motif | 1 | TCTTAC | ||
| ARE | 3 | AAACCA | ||
| I-box | 1 | GGATAAGGTG | ||
| TGACG-motif | 2 | TGACG | ||
| CGTCA-motif | 2 | CGTCA | ||
| ABRE | 3 | CACGTG | ||
| LTR | 1 | CCGAAA | ||
| SlFZY6 | AT-rich sequence | 1 | TAAAATACT | element for maximal elicitor-mediated activation |
| AT-rich element | 1 | ATAGAAATCAA | binding site of AT-rich DNA-binding protein (ATBP-1) | |
| TATA-box | 53 | TATTTAAA/ATATAT/TATA/ATTATA/TATAA/TAAAGATT/TATATA/TATATTTATATTT/TATAAAT | ||
| Box 4 | 1 | ATTAAT | ||
| CAAT-box | 20 | CAAT/CAAAT | ||
| GT1-motif | 1 | GGTTAA | ||
| ARE | 3 | AAACCA | ||
| TCCC-motif | 1 | TCTCCCT | ||
| TCA-element | 1 | CCATCTTTTT | ||
| MRE | 1 | AACCTAA | ||
| chs-CMA1a | 2 | TTACTTAA | ||
| SlFZY7 | ACA-motif | 1 | AATTACAGCCATT | part of gapA in (gapA-CMA1) involved with light responsiveness |
| 3-AF1 binding site | 1 | TAAGAGAGGAA | light-responsive element | |
| WUN-motif | 1 | AAATTTCCT | wound-responsive element | |
| Gap-box | 2 | CAAATGAA(A/G)A | part of a light-responsive element | |
| TATA-box | 33 | TATAA/TATA/ATATAT/TATATA/ATATAA/TATACA/TATAAAA | ||
| G-Box | 1 | CACGTT | ||
| Box 4 | 3 | ATTAAT | ||
| CAAT-box | 40 | CAAT/CAAAT/CCAAT | ||
| TGACG-motif | 1 | TGACG | ||
| TCA-element | 1 | CCATCTTTTT | ||
| ABRE | 1 | ACGTG | ||
| CGTCA-motif | 1 | CGTCA | ||
| LTR | 1 | CCGAAA | ||
| ATCT-motif | 1 | AATCTAATCC | ||
| SlFZY8 | G-box | 4 | TACGTG/GCCACGTGGA | |
| TGACG-motif | 2 | TGACG | ||
| CGTCA-motif | 2 | CGTCA | ||
| ABRE | 5 | ACGTG/GACACGT GGC/CACGTG/ | ||
| CAT-box | 1 | GCCACT | ||
| ARE | 2 | AAACCA | ||
| TCA-element | 1 | CCATCTTTTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, S.; Xiang, H.; Yang, X.; Ye, Y.; Ma, Y.; Han, L.; Xu, T.; Liu, Y.; Wang, F.; Qi, M.; et al. Analysis of YUC and TAA/TAR Gene Families in Tomato. Horticulturae 2023, 9, 665. https://doi.org/10.3390/horticulturae9060665
Meng S, Xiang H, Yang X, Ye Y, Ma Y, Han L, Xu T, Liu Y, Wang F, Qi M, et al. Analysis of YUC and TAA/TAR Gene Families in Tomato. Horticulturae. 2023; 9(6):665. https://doi.org/10.3390/horticulturae9060665
Chicago/Turabian StyleMeng, Sida, Hengzuo Xiang, Xiaoru Yang, Yunzhu Ye, Yuying Ma, Leilei Han, Tao Xu, Yufeng Liu, Feng Wang, Mingfang Qi, and et al. 2023. "Analysis of YUC and TAA/TAR Gene Families in Tomato" Horticulturae 9, no. 6: 665. https://doi.org/10.3390/horticulturae9060665