
Comparison of Pollen-Collecting Abilities between Apis mellifera L. and Bombus terrestris L. in the Oil Tree Peony Field
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Site
2.2. Experiment Design
2.3. Flower Trait Measurement
2.4. Definition of Flowering Stages
2.5. Pollen Load on Bees
2.6. The Pollen Quantity of P. ostii per Flower and Pollen Deposition per Visit
2.7. The Pollen Viability
2.8. Data Analysis
3. Results
3.1. Observation of Floral Traits
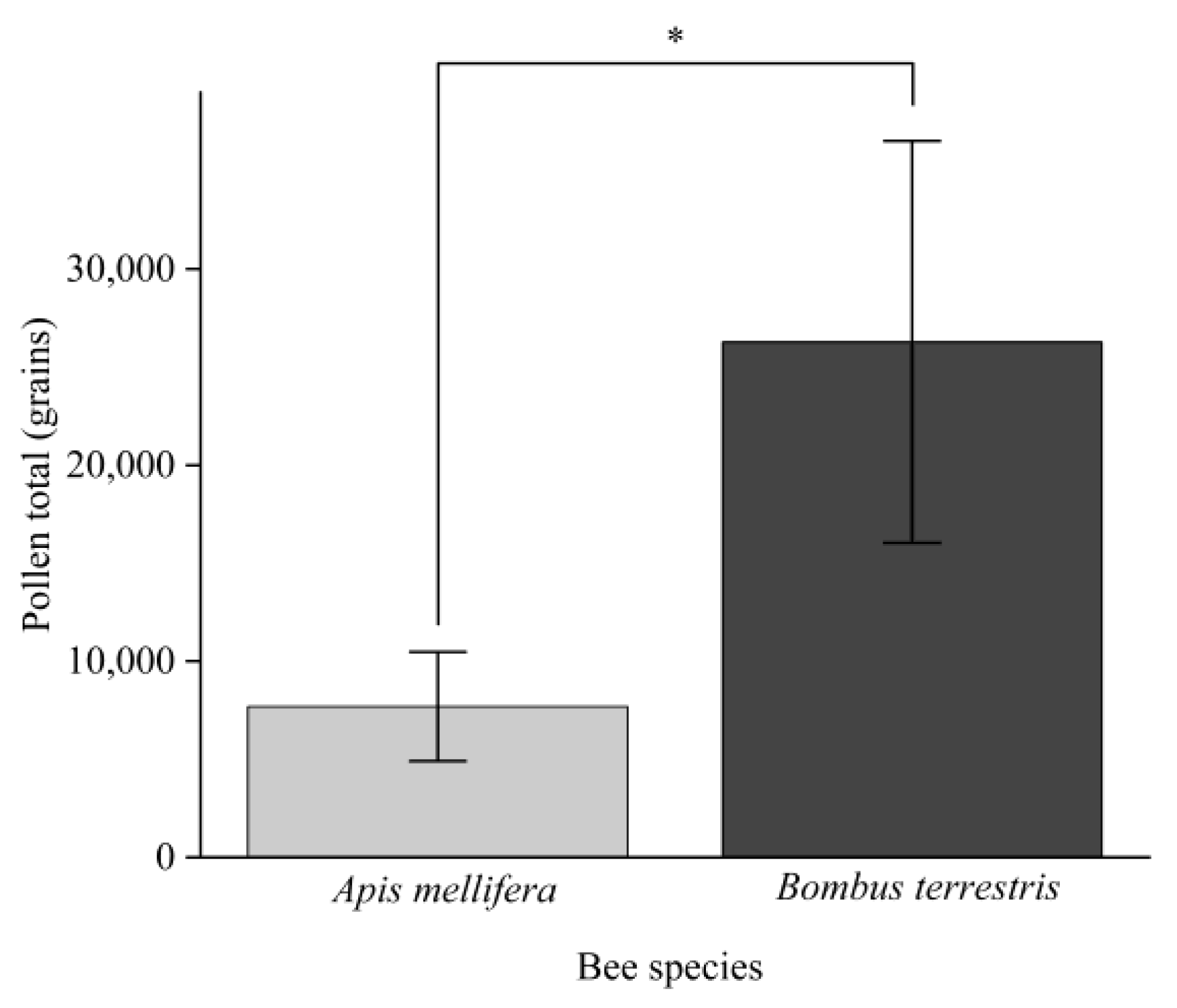
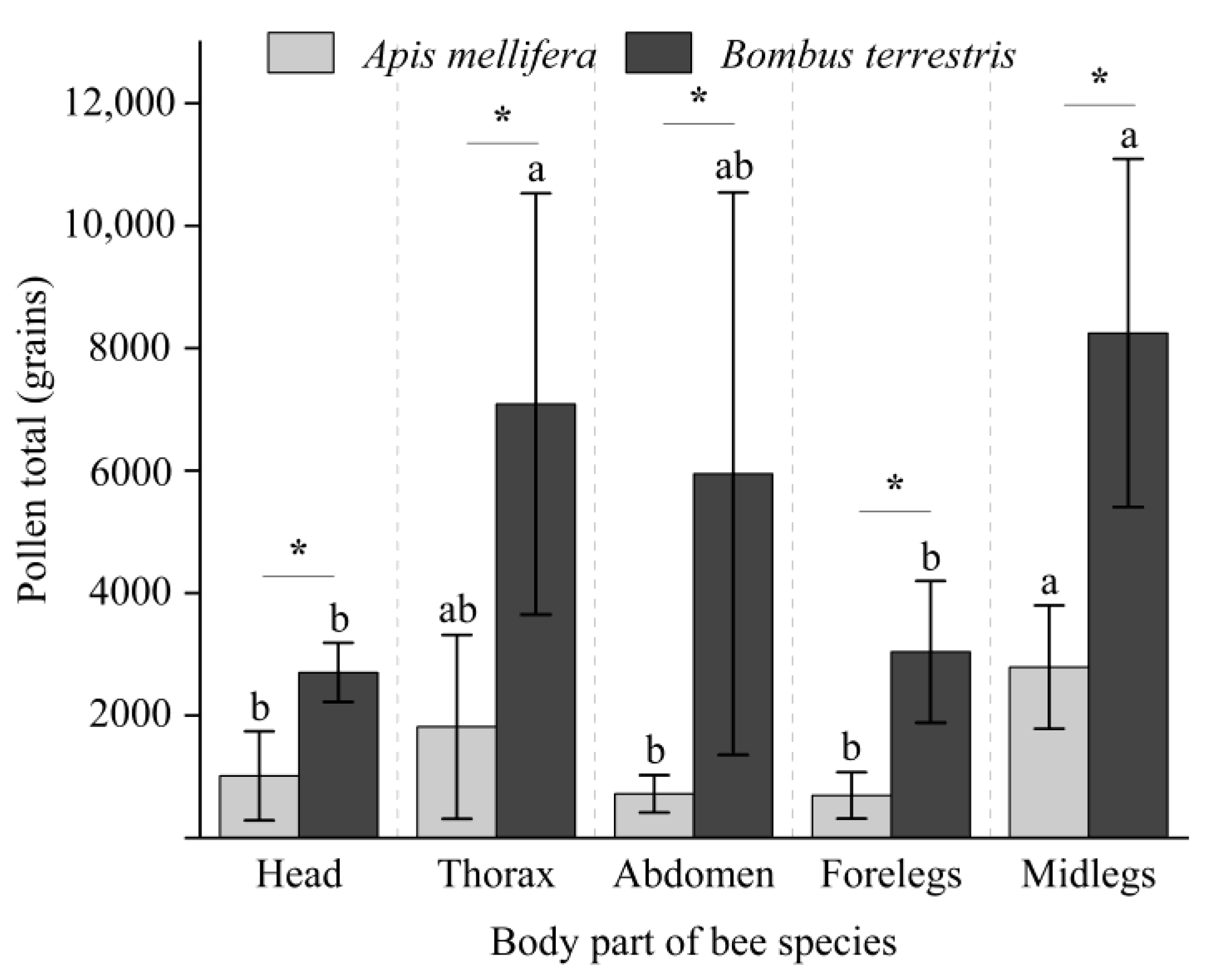
3.2. Pollen Loads on Bees
3.3. Pollen Loads on Bees
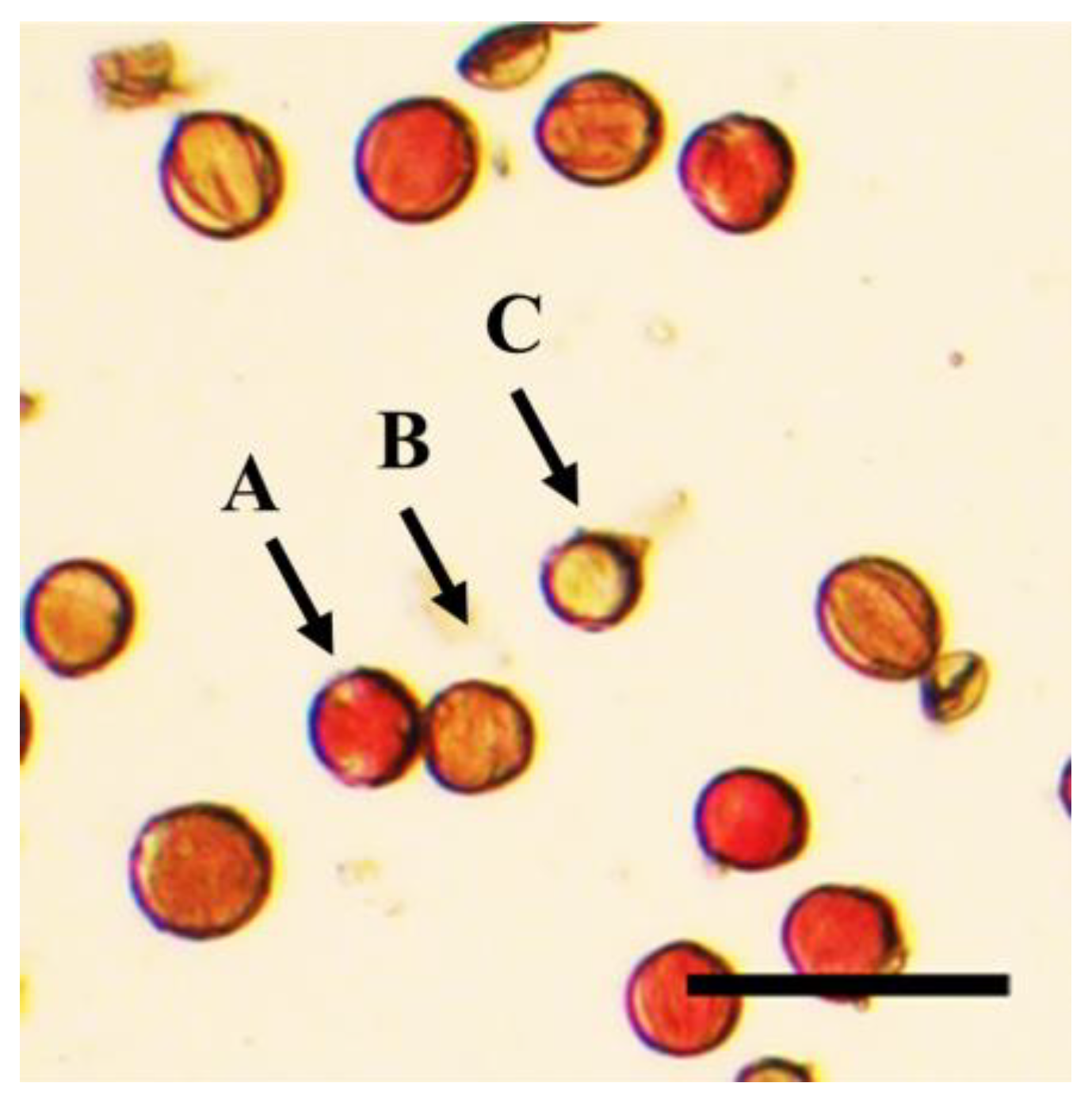
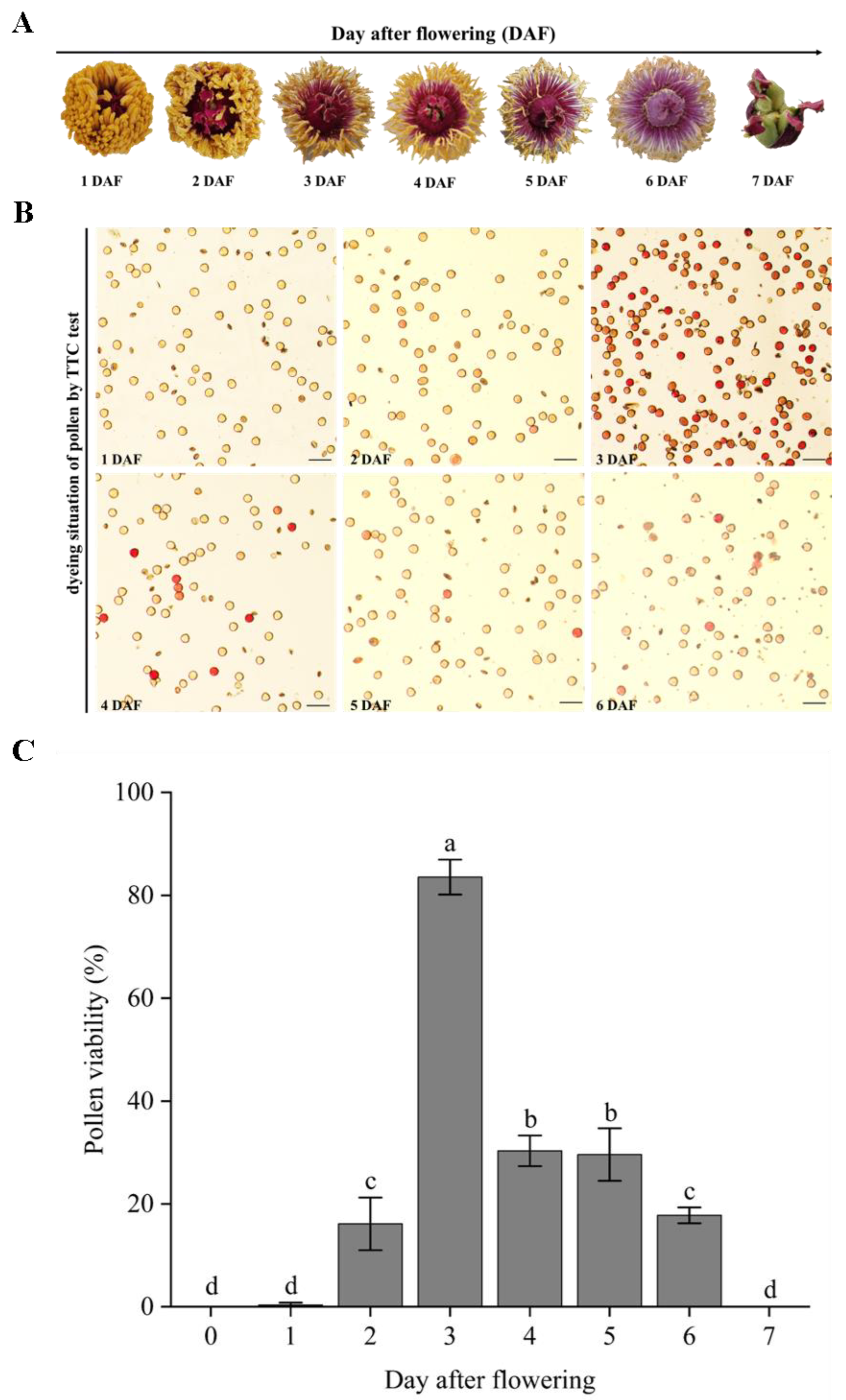
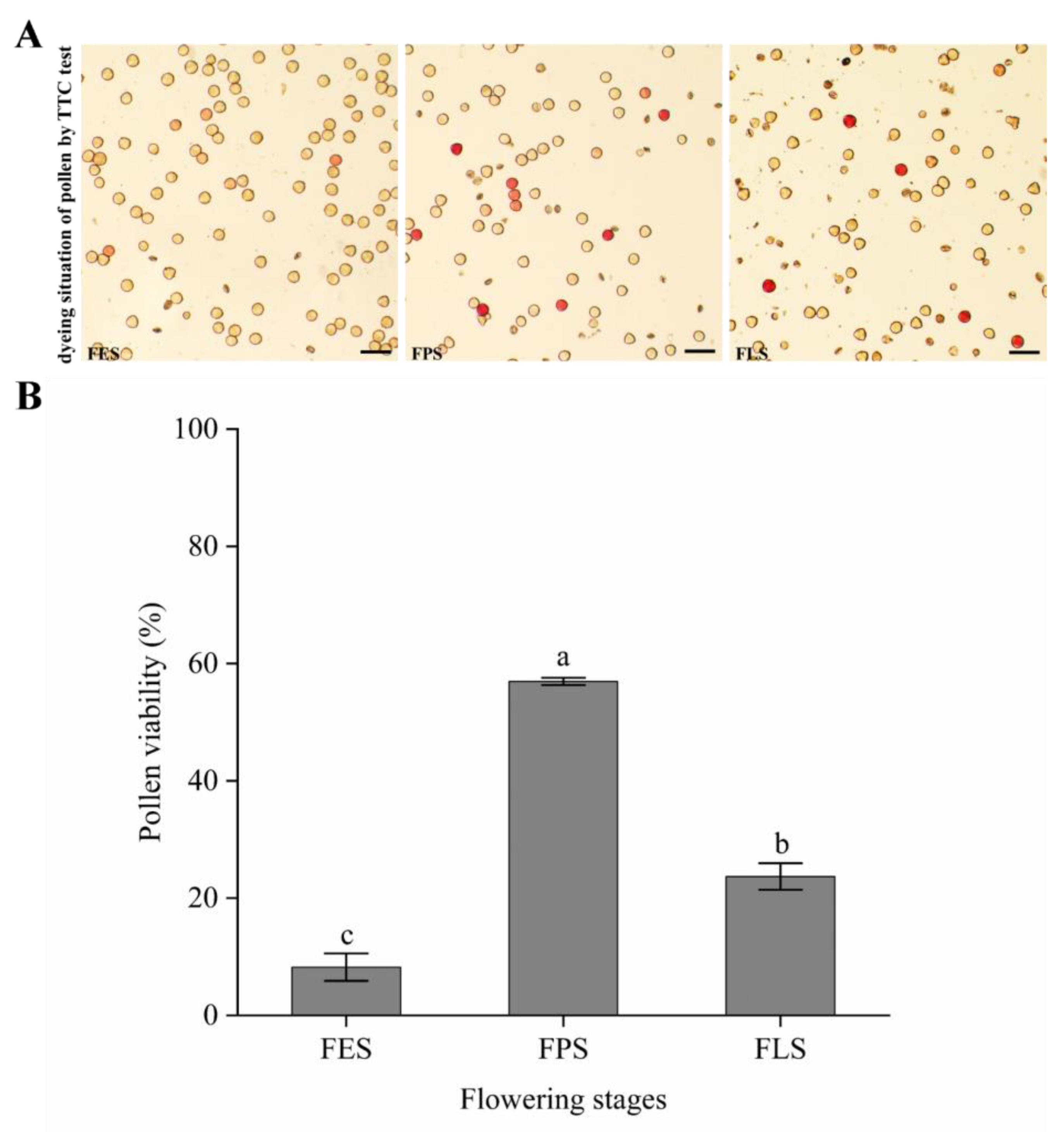
3.4. Pollen Viability of Different Days after Flowering
3.5. Pollen Viability at Different Flowering Stages
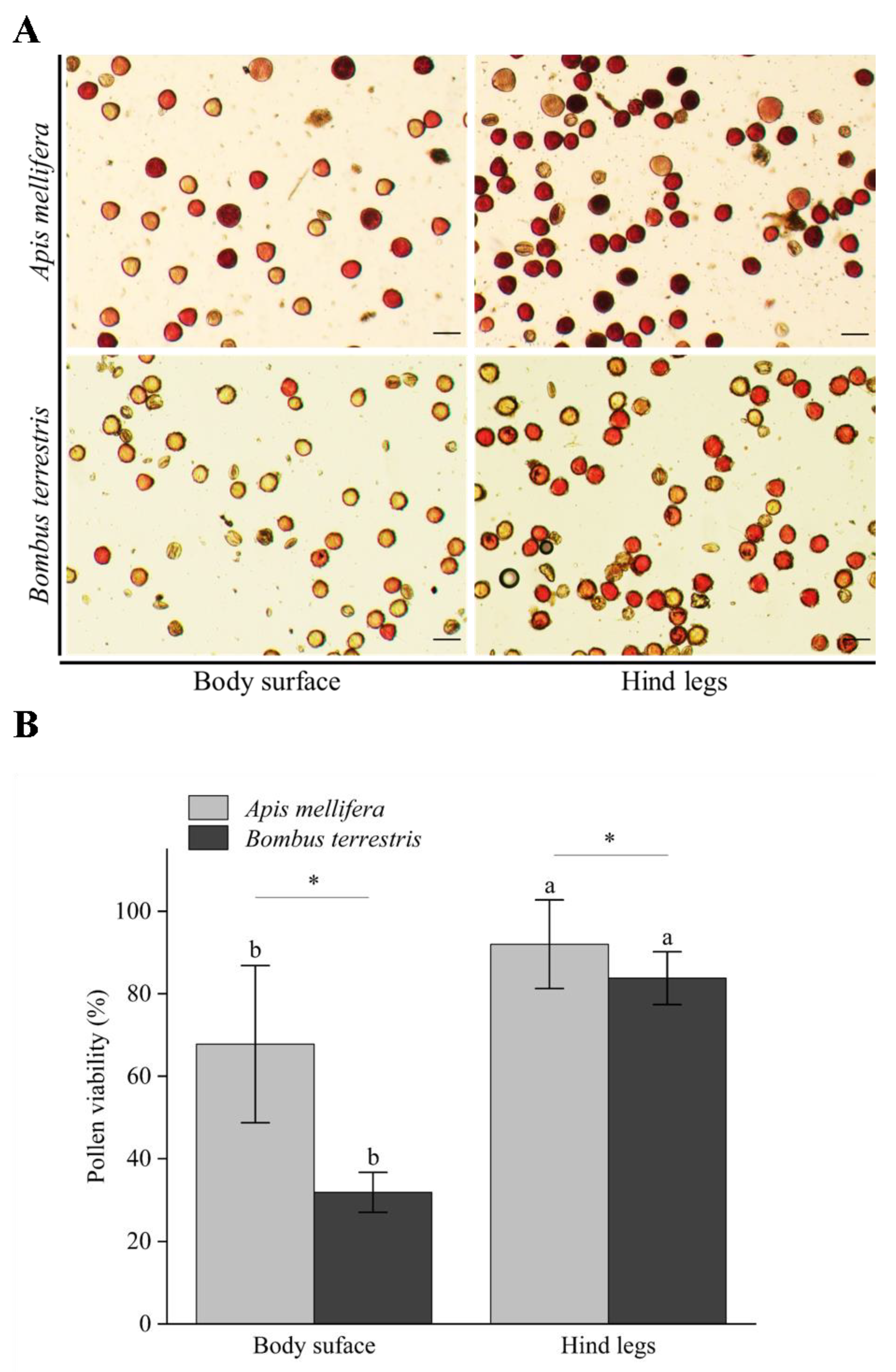
3.6. Pollen Viability Carried by Body Surface and Hind Legs of Different Pollinators
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Klein, A.M.; Vaissière, B.E.; Cane, J.H.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Tscharntke, T. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B Biol. Sci. 2007, 274, 303–313. [Google Scholar] [CrossRef] [Green Version]
- Zych, M.; Goldstein, J.; Roguz, K.; Stpiczyńska, M. The most effective pollinator revisited: Pollen dynamics in a spring-flowering herb. Arthropod-Plant Interact. 2013, 7, 315–322. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Huang, J.X.; Williams, P.H.; Vaissière, B.E.; Zhou, Z.Y.; Gai, Q.B.; Dong, J.; An, J.D. Managed bumblebees outperform honeybees in increasing peach fruit set in China: Different limiting processes with different pollinators. PLoS ONE 2015, 10, e0121143. [Google Scholar] [CrossRef] [Green Version]
- Costanza, R.; d’ Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’ Neill, R.; Paruelo, J.; et al. The value of the world’s ecosystem services and natural capital. Nature 1997, 387, 253–260. [Google Scholar] [CrossRef]
- Liu, P.F.; Wu, J.; Li, H.Y.; Lin, S.W. Economic values of bee pollination to China’s Agriculture. Sci. Agric. Sin. 2011, 44, 5117–5123. [Google Scholar]
- Khalifa, S.A.M.; Elshafiey, E.H.; Shetaia, A.A.; El-Wahed, A.A.A.; Algethami, A.F.; Musharraf, S.G.; AIAjmi, M.F.; Zhao, C.; Masry, S.H.D.; Abdel-Daim, M.M.; et al. Overview of bee pollination and its economic value for crop production. Insects 2021, 12, 688. [Google Scholar] [CrossRef]
- Biesmeijer, J.C.; Roberts, S.P.M.; Reemer, M.; Ohlemüller, R.; Edwards, M.; Peeters, T.; Schaffers, A.P.; Potts, S.G.; Kleukers, R.; Thomas, C.D.; et al. Parallel declines in pollinators and insect-pollinated plants in Britain and the Netherlands. Science 2006, 313, 351–354. [Google Scholar] [CrossRef]
- An, J.; Chen, W.F. Review of crop pollination by honey bees world-wide. Chin. Agric. Sci. Bull. 2011, 27, 374–382. [Google Scholar]
- Garibaldi, L.A.; Requier, F.; Rollin, O.; Andersson, G.K. Towards an integrated species and habitat management of crop pollination. Curr. Opin. Insect Sci. 2017, 21, 105–114. [Google Scholar] [CrossRef]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild pollinators enhance fruit set of crops regardless of honey bee abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef]
- Garantonakis, N.; Varikou, K.; Birouraki, A.; Edwards, M.; Kalliakaki, V.; Andrinopoulos, F. Comparing the pollination services of honey bees and wild bees in a watermelon field. Sci. Hortic. 2016, 204, 138–144. [Google Scholar] [CrossRef]
- Thomson, J.D.; Goodell, K. Pollen removal and deposition by honeybee and bumblebee visitors to apple and almond flowers. J. Appl. Ecol. 2001, 38, 1032–1044. [Google Scholar] [CrossRef]
- Artz, D.R.; Nault, B.A. Performance of Apis mellifera, Bombus impatiens, and Peponapis pruinosa (Hymenoptera: Apidae) as Pollinators of Pumpkin. J. Econ. Entomol. 2011, 104, 1153–1161. [Google Scholar] [CrossRef]
- Peat, J.; Darvill, B.; Ellis, J.; Goulson, D. Effects of climate on intra- and interspecific size variation in bumble-bees. Funct. Ecol. 2005, 19, 145–151. [Google Scholar] [CrossRef]
- Zhang, H.; Zhou, Z.Y.; An, J.D. Pollen release dynamics and daily patterns of pollen-collecting activity of honeybee Apis mellifera and bumblebee Bombus lantschouensis in solar greenhouse. Insects 2019, 10, 216. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.H.; Cornille, A.; Giraud, T.; Cheng, F.Y.; Hu, Y.H. Independent domestications of cultivated tree peonies from different wild peony species. Mol. Ecol. 2014, 23, 82–95. [Google Scholar] [CrossRef]
- Zhou, S.L.; Zou, X.H.; Zhou, Z.Q.; Liu, J.; Xu, C.; Yu, J.; Wang, Q.; Zhang, D.M.; Wang, X.Q.; Ge, S.; et al. Multiple species of wild tree peonies gave rise to the ‘king of flowers’ Paeonia suffruticosa Andrews. Proc. R. Soc. B Biol. Sci. 2014, 281, 20141687. [Google Scholar] [CrossRef] [Green Version]
- Hong, D.Y.; Pan, K.Y. Taxonomical history and revision of Paeonia sect. Moutan (Paeoniaceae). Acta Phytotaxon. Sin. 1999, 37, 351–368. [Google Scholar]
- Li, J.J. Studies on the origin of Chinese Mudan (Tree Peony). J. Beijing For. Univ. 1998, 20, 22–26. [Google Scholar]
- Qi, J.C.; Zhou, H.M.; Ma, J.Q.; Li, P. Analysis of the chemical constituents in peony seed oil by GC-MS. Cereals Oils 2005, 11, 22–23. [Google Scholar]
- Yi, J.P.; Zhu, W.X.; Ma, H.D.; Wang, Y.F. Optimization of ultrasonic-assisted extraction of oil from Paeonia suffruticosa Andr. seeds and its GC-MS analysis. Sci. Technol. Food Ind. 2009, 30, 198–201. [Google Scholar]
- Li, Y.C. The strategy on the oil tree peony industry in China. Strateg. Study CAE 2014, 16, 58–63. [Google Scholar]
- Zhou, L.; Wang, Y. Development and utilization of oilseed peony and its industrial development strategy in China. World For. Res. 2014, 27, 68–71. [Google Scholar]
- Cui, H.L.; Huang, N.Z.; Yan, H.C.; Liu, J.X. Correlation between yield per plant and main phenotypic traits of Paeonia suffruticosa for oil production. J. S. China Agric. Univ. 2017, 38, 86–91. [Google Scholar]
- Wang, Y.C.; Sun, H.Y.; Li, Y.R.; Chen, H.H.; Li, S.X. Analysis of the variation in yield and oil quality traits of selected Paeonia ostii ‘Feng Dan’ individuals. J. Nanjing For. Univ. Nat. Sci. Ed. 2019, 43, 155–160. [Google Scholar]
- Li, S.S.; Yuan, R.Y.; Chen, L.G.; Wang, L.S.; Hao, X.H.; Wang, L.J.; Zheng, X.C.; Du, H. Systematic qualitative and quantitative assessment of fatty acids in the seeds of 60 tree peony (Paeonia section Moutan DC.) cultivars by GC-MS. Food Chem. 2015, 173, 133–140. [Google Scholar] [CrossRef]
- Jiang, T.H.; Shan, P.P.; Huang, Z.F.; Wen, L.Z.; Sun, C.H.; Liu, K.; Zheng, C.S. Effects of nitrogen fertilization on the nitrogen uptake, accumulation, and seed quality of oil peony. Chin. J. Appl. Ecol. 2016, 27, 3257–3263. [Google Scholar]
- Duan, X.G.; Zhang, L.X.; Liu, W.; Guo, L.L.; Guo, D.L.; Hou, X.G. Effects of nitrogen application on photosynthetic characteristics and yields of oil tree peony Paeonia ostii ‘Feng Dan’. J. Nanjing For. Univ. Nat. Sci. Ed. 2018, 42, 48–54. [Google Scholar]
- Wei, S.Y.; Li, M.; Ji, W.L.; Guo, D.; Zhang, Y.L. Appropriate N, P and K fertilizer rates and combination ratios to increase seed yield and oil production of oil tree peony. J. Plant Nutr. Fertil. 2019, 25, 880–888. [Google Scholar]
- Luo, Y.B.; Pei, Y.L.; Pan, K.Y.; Hong, D.Y. A study on pollination biology of Paeonia Suffruticosa subsp. spontanea (Paeoniaceae). Acta Phytotaxon. Sin. 1998, 36, 134–144. [Google Scholar]
- Han, X.; Cheng, F.Y.; Xiao, J.J.; Wang, Y.L.; Zhang, D.; Wang, Y.; Zhong, Y. Crosses of Paeonia ostii ‘Feng Dan Bai’ as maternal parents and an analysis on the potential in tree peony breeding. J. Beijing For. Univ. 2014, 36, 121–125. [Google Scholar]
- Yang, Y.; Luo, J.T.; Zhang, B.F.; Song, H.X.; Liu, G.L.; Zeng, X.L. Studies on floral characteristics and breeding system of Paeonia decomposita. J. Plant Resour. Environ. 2015, 24, 97–104. [Google Scholar]
- Luo, C.W.; Chen, Y.; Zhang, T. Breeding system and pollination biology of Paeonia ostill T. Hong & J. X. Zhang. J. Nanjing For. Univ. Nat. Sci. Ed. 2019, 43, 37–44. [Google Scholar]
- Luo, C.W.; Chen, Y.; Zhang, T. Pollination efficiency of the major pollinators of Paeonia ostii ‘Feng Dan’. J. Nanjing For. Univ. Nat. Sci. Ed. 2019, 43, 148–154. [Google Scholar]
- Tang, Y.; Yuan, T.; Chen, T.Q. Flowering characteristics and breeding system of Paeonia ludlowii. Acta Bot.Boreal. Occident.Sin. 2021, 41, 0782–0794. [Google Scholar]
- Campbell, D.R.; Halama, K.J. Resource and pollen limitations to lifetime seed production in a natural plant population. Ecology 1993, 74, 1043–1051. [Google Scholar] [CrossRef]
- He, C.L.; Zhang, K.Y.; Hou, X.G.; Han, D.B.; Wang, S.B. Foraging behavior and pollination efficiency of Apis mellifera L. on the oil tree peony ‘Feng Dan’ (Paeonia ostii T. Hong et J.X. Zhang). Insects 2019, 10, 116. [Google Scholar] [CrossRef] [Green Version]
- He, C.L.; Zhang, K.Y.; Han, D.B. Foraging behavior of honeybees (Apis mellifera L.) and ground bumblebees (Bombus terrestris L.) and its influence on seed yield and oil quality of oil tree peony cultivar ‘Fengdan’ (Paeonia ostii T. Hong et J. X. Zhang). Sciendo 2020, 64, 131–142. [Google Scholar] [CrossRef]
- Zhou, Z.Y.; Zhang, H.; Liang, C.; Zou, Y.; Dong, J.; Yuan, X.L.; Huang, J.X.; An, J.D. Foraging preference of the honeybee Apis mellifera and the bumblebee Bombus lantschouensis (Hymenptera: Apidae) in peach greenhouse. Acta Entomol. Sinia 2015, 58, 1315–1321. [Google Scholar]
- Ye, Z.W.; Du, J.H.; Sun, M.S.; Li, L.L.; Zhang, S.L. Cluster analysis for the quantity and germinating characteristics of the pollen from 92 peach cultivars. Acta Hortic. Sin. 2010, 37, 525–531. [Google Scholar]
- Li, H.Y.; Luo, A.C.; Hao, Y.J.; Dou, F.X.; Kou, R.M.; Orr, M.C.; Zhu, C.D.; Huang, D.Y. Comparison of the pollination efficiency of Apis cerana with wild bees in oil-seed camellia fields. Basic Appl. Ecol. 2021, 56, 250–258. [Google Scholar] [CrossRef]
- Hu, S.Y. Experimental methods in plant embryology (I) determination of pollen viability. Chin. Bull. Bot. 1993, 10, 60–62. [Google Scholar]
- Gong, Y.B.; Huang, S.Q. On methodology of foraging behavior of pollinating insects. Biodivers. Sci. 2007, 15, 576–583. [Google Scholar]
- Zhang, K.Y.; He, C.L.; Chen, L.L.; Han, D.B.; Hou, X.G. Study of pollination ecology of bumblebees on Paeonia ostii ‘Fengdan’. J. Henan Agric. Sci. 2019, 48, 102–109. [Google Scholar]
- Robertson, A.W. The relationship between floral display size, pollen carryover and geitonogamy in Myosotis colensoi (Kirk) Macbride (Boraginaceae). Biol. J. Linn. Soc. 1992, 46, 333–349. [Google Scholar] [CrossRef]
- de Vega, C.; Arista, M.; Ortiz, P.L.; Herrera, C.M.; Talavera, S. The ant-pollination system of Cytinus hypocistis (Cytinaceae), a Mediterranean root holoparasite. Ann. Bot. 2009, 103, 1065–1075. [Google Scholar] [CrossRef] [PubMed]
- Luo, C.W. Pollination efficiency of major pollinators for Jatropha curcas. Southwest China J. Agric. Sci. 2018, 31, 849–855. [Google Scholar]
- Wu, T.; Tang, J.; Huang, S.Q. Foraging behavior and pollination efficiency of generalist insects in an understory dioecious shrub Helwingia japonica. Am. J. Bot. 2020, 107, 1274–1282. [Google Scholar] [CrossRef]
- Zhang, K.Y.; He, C.L.; Wang, S.B.; Hou, X.G. Influence of pollination methods on fruit development, fruit yield and oil quality in oil tree peony. Sci. Hortic. 2022, 295, 110877. [Google Scholar] [CrossRef]
- Zhang, K.Y.; Li, Y.Y.; Sun, K.L.; Bao, J.Y.; He, C.L.; Hou, X.G. Supplementary honey bee (Apis mellifera L.) pollination enhances fruit growth rate and fruit yield in Paeonia ostii (family: Paeoniaceae). PLoS ONE 2022, 17, e0272921. [Google Scholar] [CrossRef]
- Velthuis, H.H.W.; Doorn, A.V. A century of advances in bumblebee domestication and the economic and environmental aspects of its commercialization for pollination. Apidologie 2006, 37, 421–451. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.Q.; Guo, Y.H.; Pan, M.Q.; Chen, J.K. Floral syndrome and insect pollination of Liriodendron chinense. Acta Bot. Sin. 1999, 41, 241–248. [Google Scholar]
- Huang, S.Q.; Guo, Y.H. Research progress of pollination biology. Chin. Sci. Bull. 2000, 45, 225–237. [Google Scholar]
- Huang, S.Q. Studies on plant- pollinator interaction and its significances. Biodivers. Sci. 2007, 15, 569–575. [Google Scholar]
- Snow, A.A.; Spora, T.P. Individual variation in the vigor of self pollen and selfed progeny in Hibiscus moscheutos (Malvaceae). Am. J. Bot. 1993, 80, 160–164. [Google Scholar] [CrossRef]
- Wang, Q.L.; Lu, L.D.; Wu, X.Q.; Chen, Z.K.; Lin, J.X. Pollen preservation and its viability test. Chin. Bull. Bot. 2002, 19, 365–373. [Google Scholar]
- Yin, J.L.; Zhao, H.E. Summary of influencial factors on pollen viability and its preservation methods. Chin. Agric. Sci. Bull. 2005, 21, 110–113. [Google Scholar]
- Kocot, J.; Kiełczykowska, M.; Luchowska-Kocot, D.; Kurzepa, J.; Musik, I. Antioxidant potential of propolis, bee pollen, and royal jelly: Possible medical application. Oxidative Med. Cell. Longev. 2018, 2018, 7074209. [Google Scholar] [CrossRef]
- Parker, A.J.; Tran, J.L.; Ison, J.L.; Bai, J.D.K.; Weis, A.E.; Thomson, J.D. Pollen packing affects the function of pollen on corbiculate bees but not non-corbiculate bees. Arthropod-Plant Interact. 2015, 9, 197–203. [Google Scholar] [CrossRef]
- Sliva, T.M.S.; Camara, C.A.; Lins, A.C.D.S.; Barbosa-Filho, J.M.; da Silva, E.M.S.; Freitas, B.M.; dos Santos, F.D.A.R. Chemical composition and free radical scavenging activity of pollen loads from stingless bee Melipona subnitida Ducke. J. Food Compos. Anal. 2006, 19, 507–511. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bee Species | Pollen Deposition per Visit | Pollen Transfer Efficiency (%) |
|---|---|---|
| A. mellifera | 27,000.00 a ± 5613.70 (n = 3) | 0.17 |
| B. terrestris | 7746.67 b ± 3089.68 (n = 3) | 0.05 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, J.; Zhang, K.; He, X.; Chen, Z.; Wang, J.; He, C.; Hou, X. Comparison of Pollen-Collecting Abilities between Apis mellifera L. and Bombus terrestris L. in the Oil Tree Peony Field. Horticulturae 2023, 9, 658. https://doi.org/10.3390/horticulturae9060658
Bao J, Zhang K, He X, Chen Z, Wang J, He C, Hou X. Comparison of Pollen-Collecting Abilities between Apis mellifera L. and Bombus terrestris L. in the Oil Tree Peony Field. Horticulturae. 2023; 9(6):658. https://doi.org/10.3390/horticulturae9060658
Chicago/Turabian StyleBao, Junyi, Kaiyue Zhang, Xiangnan He, Zhanfeng Chen, Junying Wang, Chunling He, and Xiaogai Hou. 2023. "Comparison of Pollen-Collecting Abilities between Apis mellifera L. and Bombus terrestris L. in the Oil Tree Peony Field" Horticulturae 9, no. 6: 658. https://doi.org/10.3390/horticulturae9060658