
Exploring Genetic Diversity in an Ilex crenata Breeding Germplasm
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Plant Material for the Ilex Collection
2.2. Genome Size of the Collected Ilex Accessions
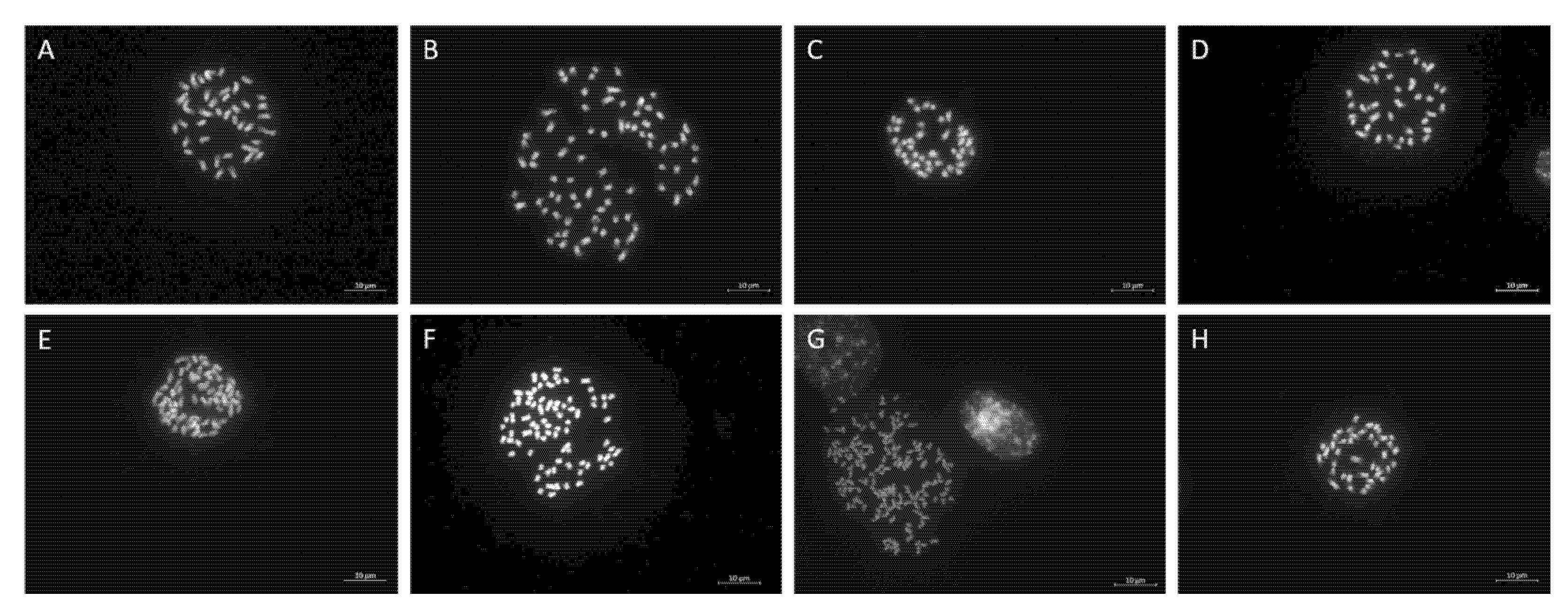
2.3. Chromosome Number of the Collected Ilex Accessions
2.4. GBS Fingerprinting of the ILEX Collection
2.4.1. Library Preparation and Sequencing
2.4.2. GBS Data Analysis
3. Results and Discussion
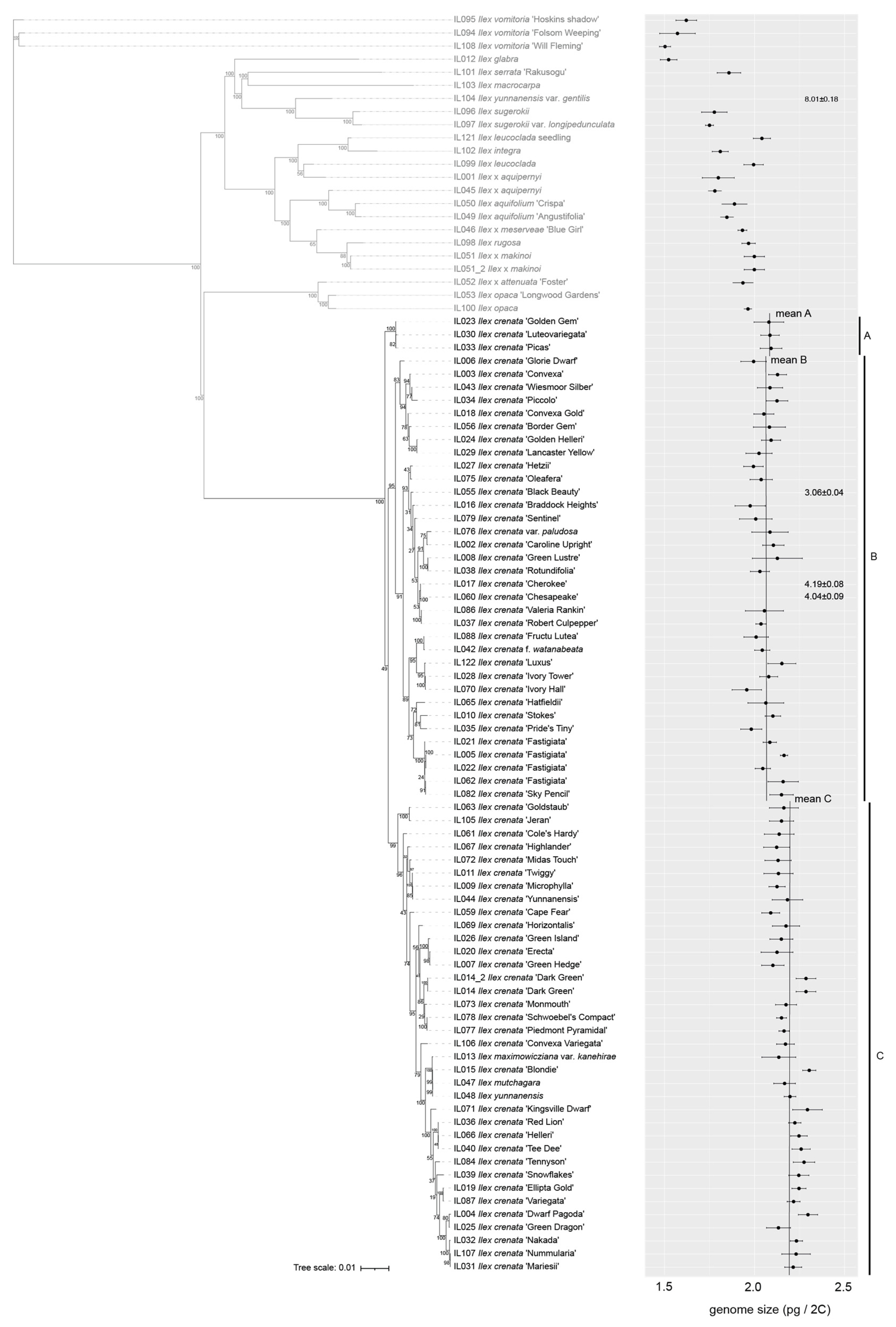
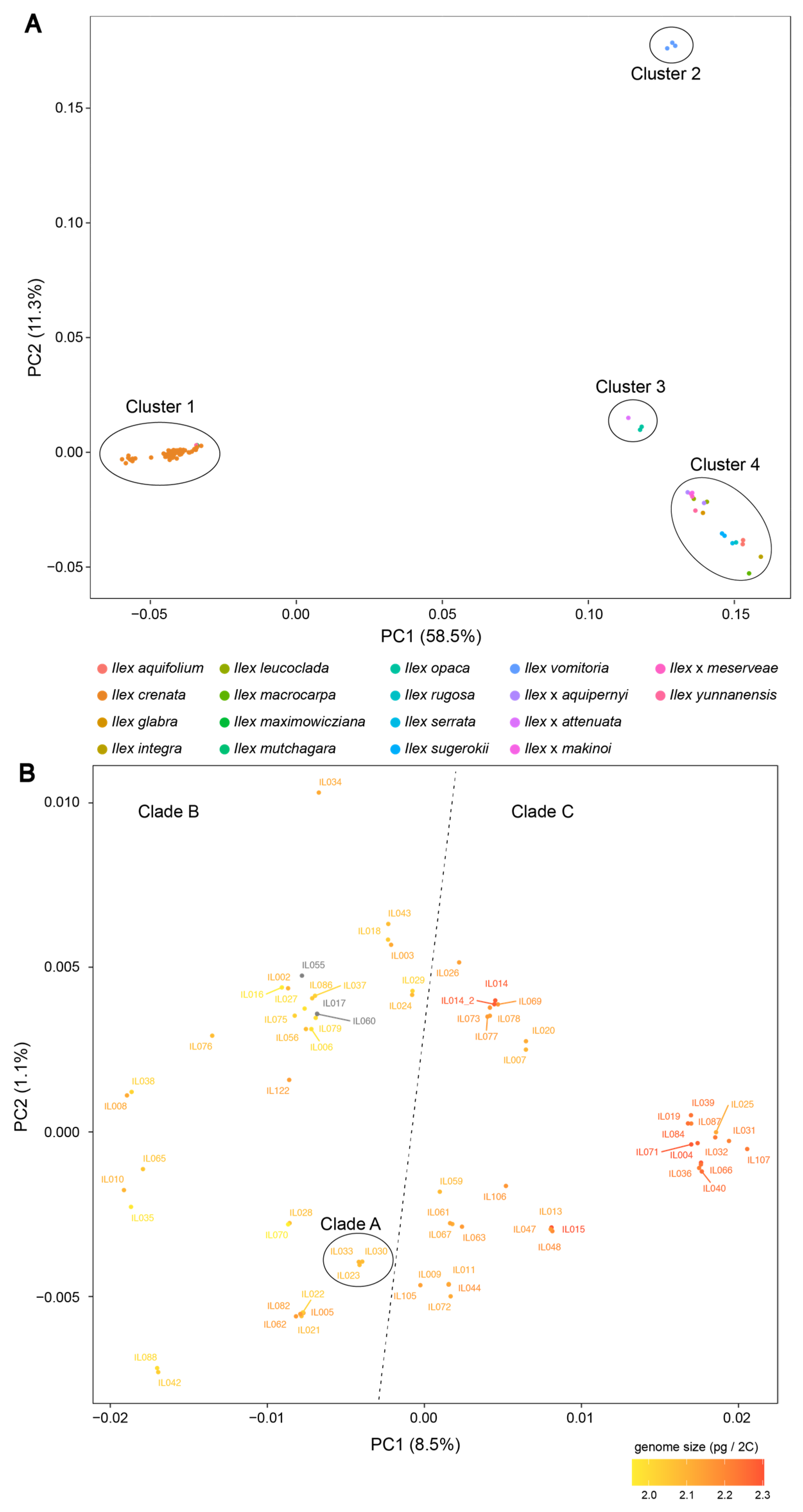
3.1. Genetic Identity
3.2. Genetic Relationships among Ilex Species
3.3. Genetic Relationships of Ilex Crenata Cultivars
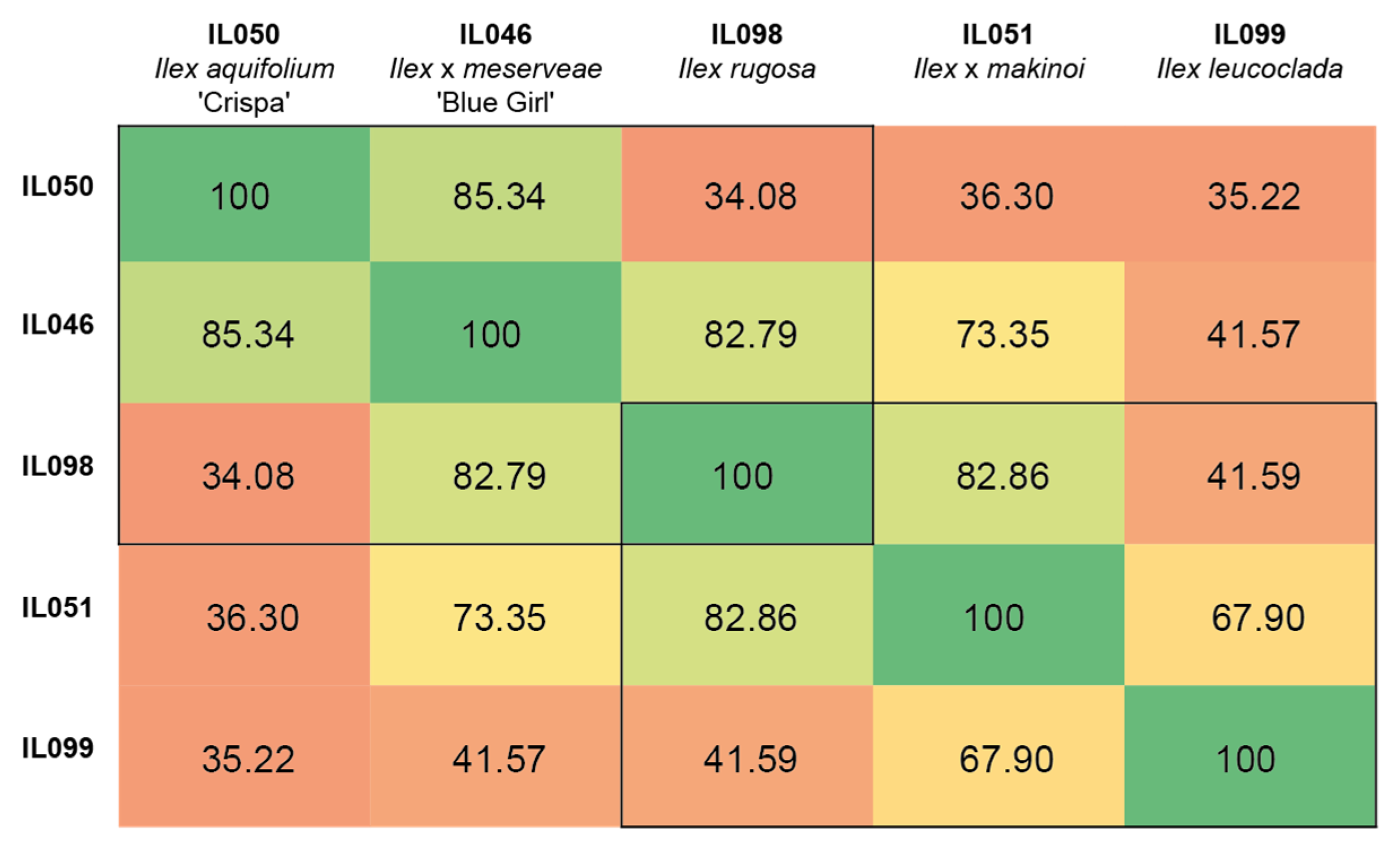
3.4. Genetic Diversity via Crossing and Hybridization
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- POWO. Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. 2022. Available online: https://powo.science.kew.org/ (accessed on 27 February 2023).
- Cuénoud, P.; Del Pero Martinez, M.A.; Loizeau, P.-A.; Spichiger, R.; Andrews, S.; Manen, J.-F. Molecular phylogeny and biogeography of the genus Ilex L. (Aquifoliaceae). Ann. Bot. 2000, 85, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Manen, J.-F.; Barriera, G.; Loizeau, P.-A.; Naciri, Y. The history of extant Ilex species (Aquifoliaceae): Evidence of hybridisation within a Miocene radiation. Mol. Phylogenetics Evol. 2010, 57, 961–977. [Google Scholar] [CrossRef]
- Yao, X.; Lu, Z.; Song, Y.; Hu, X.; Corlett, R.T. A chromosome-scale genome assembly for the holly (Ilex polyneura) provides insights into genomic adaptations to elevation in Southwest China. Hortic. Res. 2022, 9, uhab049. [Google Scholar] [CrossRef]
- Dudley, T.R.; Eisenbeiss, G.R. International Checklist of Cultivated Ilex: Part 2, Ilex crenata Thunberg ex. J.A. Murray; U.S. Department of Agriculture: Washington, DC, USA; U.S. National Arboretum Contribution: Washington, DC, USA, 1992; Volume 6, p. 91. [Google Scholar]
- Xu, K.-W.; Wei, X.-F.; Lin, C.-X.; Zhang, M.; Zhang, Q.; Zhou, P.; Fang, Y.-M.; Xue, J.-Y.; Duan, Y.-F. The chromosome-level holly (Ilex latifolia) genome reveals key enzymes in triterpenoid saponin biosynthesis and fruit color change. Front. Plant Sci. 2022, 13, 982323. [Google Scholar] [CrossRef] [PubMed]
- Kong, B.L.-H.; Nong, W.; Wong, K.-H.; Law, S.T.-S.; So, W.-L.; Chan, J.J.-S.; Zhang, J.; Lau, T.-W.D.; Hui, J.H.-L.; Shaw, P.-C. Chromosomal level genome of Ilex asprella and insight into antiviral triterpenoid pathway. Genomics 2022, 114, 110366. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Zhang, F.; Corlett, R.T. Utilization of the Hollies (Ilex L. spp.): A Review. Forests 2022, 13, 94. [Google Scholar] [CrossRef]
- Galle, F.C. Hollies: The genus Ilex; Timber Press, Inc.: Portland, OR, USA, 1997; p. 573. [Google Scholar]
- Molnar, T.; Capik, J.M. The Rutgers University woody ornamentals breeding program: Past, present, and future. Acta Hortic. 2013, 990, 11. [Google Scholar] [CrossRef]
- Zhang, L.; Cao, B.; Bai, C. New reports of nuclear DNA content for 66 traditional Chinese medicinal plant taxa in China. Caryologia 2013, 66, 375–383. [Google Scholar] [CrossRef]
- Bai, C.; Alverson, W.S.; Follansbee, A.; Waller, D.M. New reports of nuclear DNA content for 407 U.S. plant species. Ann. Bot. 2012, 110, 1623–1629. [Google Scholar] [CrossRef]
- Sugiura, T. A list of chromosome numbers in angiospermous plants. Shokubutsugaku Zasshi 1931, 45, 353–355. [Google Scholar] [CrossRef]
- Fedorov, A. Chromosome Numbers of Flowering Plants; Reprint 1974; Academy of Sciences of the USSR: Moscow, Russia; Komarov Botanical Institute: Leningrad, Russia, 1969. [Google Scholar]
- Goldblatt, P. New or noteworthy chromosome records in the angiosperms. Ann. Mo. Bot. Gard. 1976, 63, 889–895. [Google Scholar] [CrossRef]
- Rice, A.; Glick, L.; Abadi, S.; Einhorn, M.; Kopelman, N.M.; Salman-Minkov, A.; Mayzel, J.; Chay, O.; Mayrose, I. The Chromosome Counts Database (CCDB)—A community resource of plant chromosome numbers. New Phytol. 2014, 206, 7. [Google Scholar] [CrossRef]
- Darlington, C.D.; Wylie, A.P. Chromosome Atlas of Flowering Plants; Allen & Unwin: Crows Nest, Australia, 1955. [Google Scholar]
- Poland, J.A.; Rife, T.W. Genotyping-by-sequencing for plant breeding and genetics. Plant Genome 2012, 5, 92–102. [Google Scholar] [CrossRef] [Green Version]
- Van Oost, E.; Leus, L.; De Rybel, B.; Van Laere, K. Determination of genetic distance, genome size and chromosome numbers to support breeding in ornamental Lavandula species. Agronomy 2021, 11, 2173. [Google Scholar] [CrossRef]
- Greilhuber, J.; Dolezel, J.; Lysák, M.A.; Bennet, M.D. The origin, evolution and proposed stabilization of the terms ‘genome size’ and ‘C-value’ to describe nuclear DNA contents. Ann. Bot. 2005, 95, 255–260. [Google Scholar] [CrossRef] [PubMed]
- Doležel, J.; Greilhuber, J.; Lucretti, S.; Meister, A.; Lysák, M.A.; Nardi, L.; Obermayer, R. Plant genome size estimation by flow cytometry: Inter-laboratory comparison. Ann. Bot. 1998, 82, 17–26. [Google Scholar] [CrossRef] [Green Version]
- Lysak, M.A.; Dolezel, J. Estimation of nuclear DNA content in Sesleria (Poaceae). Caryologia 1998, 51, 123–132. [Google Scholar] [CrossRef]
- Kirov, I.; Divashuk, M.; Van Laere, K.; Soloviev, A.; Khrustaleva, L. An easy “SteamDrop” method for high quality plant chromosome preparation. Mol. Cytogenet. 2014, 7, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kirov, I.; Khrustaleva, L.; Van Laere, K.; Meeus, S.; Romanov, D.; Soloviev, A.; Fesenko, I. DRAWID: User-friendly java software for chromosome measurements and idiogram drawing. Comp. Cytogenet. 2017, 11, 10. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem. Bull. 1987, 19, 11–15. [Google Scholar]
- Schaumont, D. GBprocesS: Genotyping-by-Sequencing Data Processing Toolkit. Available online: https://gitlab.com/ilvo/GBprocesS. (accessed on 27 May 2021).
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd mergeR. Bioinformatics 2013, 30, 614–620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hapke, A.; Thiele, D. GIbPSs: A toolkit for fast and accurate analyses ofgenotyping-by-sequencing data without a reference genome. Mol. Ecol. Resour. 2016, 16, 11. [Google Scholar] [CrossRef] [PubMed]
- Van Poucke, K.; Haegeman, A.; Goedefroit, T.; Focquet, F.; Leus, L.; Jung, M.H.; Nave, C.; Redondo, M.A.; Husson, C.; Kostov, K.; et al. Unravelling hybridization in Phytophthora using phylogenomics and genome size estimation. IMA Fungus 2021, 12, 16. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef]
- Mowforth, M.A. Variation in Nuclear DNA Amounts in Flowering Plants: An Ecological Analysis; University of Sheffield: Sheffield, UK, 1986. [Google Scholar]
- Noirot, M.; Barre, P.; Louarn, J.; Duperray, C.; Hamon, S. Nucleus-cytosol interactions—A source of stoichiometric error in flow cytometric estimation of nuclear DNA content in plants. Ann. Bot. 2000, 86, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Greilhuber, J. Intraspecific variation in genome size in angiosperms: Identifying its existence. Ann. Bot. 2005, 95, 91–98. [Google Scholar] [CrossRef] [Green Version]
- Noirot, M.; Poncet, V.; Barre, P.; Hamon, P.; Hamon, S.; De Kochko, A. Genome size variations in diploid African Coffea species. Ann. Bot. 2003, 92, 709–714. [Google Scholar] [CrossRef] [Green Version]
- Bilinski, P.; Albert, P.S.; Berg, J.J.; Birchler, J.A.; Grote, M.N.; Lorant, A.; Quezada, J.; Swarts, K.; Yang, J.; Ross-Ibarra, J. Parallel altitudinal clines reveal trends in adaptive evolution of genome size in Zea mays. PLoS Genet. 2018, 14, e1007162. [Google Scholar] [CrossRef] [Green Version]
- Díez, C.M.; Gaut, B.S.; Meca, E.; Scheinvar, E.; Montez-Hernandez, S.; Eguiarte, L.E.; Tenaillon, M.I. Genome size variation in wild and cultivated maize along altitudinal gradients. New Phytol. 2013, 199, 264–276. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| ILVO Accession Number | Species and Cultivar Name | Collection of Origin | Arboretum Accession Number | Mean Genome Size (pg/2C ± sd) | Chromosome Number |
|---|---|---|---|---|---|
| IL001 | Ilex x aquipernyi | Esveld | 1.80 ± 0.09 | 40 | |
| IL002 | Ilex crenata ‘Caroline Upright’ | Esveld | 2.11 ± 0.06 | ||
| IL003 | Ilex crenata ‘Convexa’ | Esveld | 2.13 ± 0.05 | ||
| IL004 | Ilex crenata ‘Dwarf Pagoda’ | Esveld | 2.30 ± 0.05 | ||
| IL005 a | Ilex crenata ‘Fastigiata’ | Esveld | 2.16 ± 0.02 | ||
| IL006 | Ilex crenata ‘Glorie Dwarf’ | Esveld | 1.99 ± 0.07 | ||
| IL007 | Ilex crenata ‘Green Hedge’ | Esveld | 2.10 ± 0.06 | ||
| IL008 | Ilex crenata ‘Green Lustre’ | Esveld | 2.02 ± 0.14 | ||
| IL009 | Ilex crenata ‘Microphylla’ | Esveld | 2.16 ± 0.05 | ||
| IL010 | Ilex crenata ‘Stokes’ | Esveld | 2.10 ± 0.04 | ||
| IL011 | Ilex crenata ‘Twiggy’ | Esveld | 2.13 ± 0.08 | ||
| IL012 | Ilex glabra | Esveld | 1.52 ± 0.05 | 40 | |
| IL013 b,* | Ilex maximowicziana var. kanehirae | Esveld | 2.14 ± 0.09 | 40 | |
| IL014 c | Ilex crenata ‘Dark Green’ | Plant Select | 2.18 ± 0.05 | ||
| IL015 b | Ilex crenata ‘Blondie’ | Plant Select | 2.30 ± 0.04 | ||
| IL016 | Ilex crenata ‘Braddock Heights’ | Het Leen | 00004670 | 1.98 ± 0.08 | 40 |
| IL017 | Ilex crenata ‘Cherokee’ | Het Leen | 00004020 | 4.19 ± 0.08 | 80 |
| IL018 | Ilex crenata ‘Convexa Gold’ | Het Leen | 00004672 | 2.05 ± 0.06 | |
| IL019 | Ilex crenata ‘Ellipta Gold’ | Het Leen | 00004017 | 2.25 ± 0.04 | |
| IL020 | Ilex crenata ‘Erecta’ | Het Leen | 00004021 | 2.12 ± 0.09 | |
| IL021 a | Ilex crenata ‘Fastigiata’ | Het Leen | 00001571 | 2.09 ± 0.04 | 40 |
| IL022 a | Ilex crenata ‘Fastigiata’ | Het Leen | 00001904 | 2.05 ± 0.04 | |
| IL023 | Ilex crenata ‘Golden Gem’ | Het Leen | 00001905 | 2.08 ± 0.08 | 40 |
| IL024 d | Ilex crenata ‘Golden Helleri’ | Het Leen | 00004014 | 2.09 ± 0.05 | |
| IL025 | Ilex crenata ‘Green Dragon’ | Het Leen | 00004013 | 2.13 ± 0.07 | |
| IL026 | Ilex crenata ‘Green Island’ | Het Leen | 00004090 | 2.12 ± 0.06 | |
| IL027 | Ilex crenata ‘Hetzii’ | Het Leen | 00004022 | 1.99 ± 0.05 | 40 |
| IL028 | Ilex crenata ‘Ivory Tower’ | Het Leen | 00004673 | 2.08 ± 0.05 | |
| IL029 d | Ilex crenata ‘Lancaster Yellow’ | Het Leen | 00004024 | 2.03 ± 0.07 | |
| IL030 | Ilex crenata ‘Luteovariegata’ | Het Leen | 00005561 | 2.09 ± 0.05 | |
| IL031 | Ilex crenata ‘Mariesii’ | Het Leen | 00004010 | 2.22 ± 0.05 | |
| IL032 | Ilex crenata ‘Nakada’ | Het Leen | 00004016 | 2.23 ± 0.03 | |
| IL033 | Ilex crenata ‘Picas’ | Het Leen | 00004091 | 2.09 ± 0.06 | |
| IL034 | Ilex crenata ‘Piccolo’ | Het Leen | 00004004 | 2.13 ± 0.06 | |
| IL035 | Ilex crenata ‘Pride’s Tiny’ | Het Leen | 00004023 | 1.98 ± 0.06 | 40 |
| IL036 | Ilex crenata ‘Red Lion’ | Het Leen | 00004018 | 2.22 ± 0.03 | |
| IL037 | Ilex crenata ‘Robert Culpepper’ | Het Leen | 00004011 | 2.04 ± 0.03 | |
| IL038 | Ilex crenata ‘Rotundifolia’ | Het Leen | 00004671 | 2.03 ± 0.05 | |
| IL039 | Ilex crenata ‘Snowflakes’ | Het Leen | 00004669 | 2.25 ± 0.06 | |
| IL040 | Ilex crenata ‘Tee Dee’ | Het Leen | 00004025 | 2.26 ± 0.05 | |
| IL042 | Ilex crenata f. watanabeata | Het Leen | 00005253 | 2.04 ± 0.04 | |
| IL043 | Ilex crenata ‘Wiesmoor Silber’ | Het Leen | 00004012 | 2.09 ± 0.07 | |
| IL044 | Ilex crenata ‘Yunnanensis’ | Het Leen | 00004019 | 2.18 ± 0.09 | |
| IL045 | Ilex x aquipernyi | Het Leen | 00005002 | 1.78 ± 0.04 | |
| IL046 | Ilex x meserveae ‘Blue Girl’ | Het Leen | 00001829 | 1.93 ± 0.02 | 40 |
| IL047 b,* | Ilex mutchagara Makinoi | Het Leen | 00001901 | 2.17 ± 0.06 | |
| IL048 b,* | Ilex yunnanensis | Het Leen | 00004449 | 2.20 ± 0.03 | |
| IL049 | Ilex aquifolium ‘Angustifolia’ | Het Leen | 00001859 | 1.85 ± 0.04 | |
| IL050 | Ilex aquifolium ‘Crispa’ | Het Leen | 00001873 | 1.90 ± 0.07 | |
| IL051 c | Ilex x makinoi Hara | Het Leen | 00004998 | 2.00 ± 0.06 | 40 |
| IL052 | Ilex x attenuata ‘Foster’ | Het Leen | 00004994 | 1.89 ± 0.05 | |
| IL053 | Ilex opaca ‘Longwood Gardens’ | Het Leen | 00004993 | ||
| IL055 | Ilex crenata ‘Black Beauty’ | Bokrijk | 19991318 | 3.06 ± 0.04 | 60 |
| IL056 | Ilex crenata ‘Border Gem’ | Bokrijk | 19991317 | 2.07 ± 0.09 | |
| IL059 | Ilex crenata ‘Cape Fear’ | Bokrijk | 19991320 | 2.09 ± 0.05 | |
| IL060 | Ilex crenata ‘Chesapeake’ | Bokrijk | 19900267 | 4.04 ± 0.09 | 80 |
| IL061 | Ilex crenata ‘Cole’s Hardy’ | Bokrijk | 1983ACQI175 | 2.14 ± 0.08 | |
| IL062 a | Ilex crenata ‘Fastigiata’ | Bokrijk | 19900529 | 2.16 ± 0.08 | |
| IL063 | Ilex crenata ‘Goldstaub’ | Bokrijk | 19940813 | 2.16 ± 0.08 | |
| IL065 | Ilex crenata ‘Hatfieldii’ | Bokrijk | 19950491 | 2.06 ± 0.10 | |
| IL066 | Ilex crenata ‘Helleri’ | Bokrijk | 19940347 | 2.25 ± 0.05 | |
| IL067 | Ilex crenata ‘Highlander’ | Bokrijk | 19950296 | 2.12 ± 0.07 | |
| IL069 | Ilex crenata ‘Horizontalis’ | Bokrijk | 19910107 | 2.18 ± 0.08 | |
| IL070 | Ilex crenata ‘Ivory Hall’ | Bokrijk | 19991306 | 2.09 ± 0.08 | 40 |
| IL071 | Ilex crenata ‘Kingsville Dwarf’ | Bokrijk | 19940329 | 2.29 ± 0.08 | |
| IL072 | Ilex crenata ‘Midas Touch’ | Bokrijk | 19900222 | 2.13 ± 0.07 | |
| IL073 | Ilex crenata ‘Monmouth’ | Bokrijk | 19950499 | 2.18 ± 0.06 | |
| IL075 | Ilex crenata ‘Oleafera’ | Bokrijk | 19980688 | 2.04 ± 0.06 | |
| IL076 | Ilex crenata var. paludosa | Bokrijk | 19991354 | 2.05 ± 0.10 | 40 |
| IL077 | Ilex crenata ‘Piedmont Pyramidal’ | Bokrijk | 19900271 | 2.16 ± 0.03 | |
| IL078 | Ilex crenata ‘Schwoebel’s Compact’ | Bokrijk | 19950345 | 2.15 ± 0.03 | |
| IL079 | Ilex crenata ‘Sentinel’ | Bokrijk | 1988ACQI162 | 2.01 ± 0.09 | |
| IL082 a | Ilex crenata ‘Sky Pencil’ | Bokrijk | 20040562 | 2.15 ± 0.07 | |
| IL084 | Ilex crenata ‘Tennyson’ | Bokrijk | 19900395 | 2.28 ± 0.06 | |
| IL086 | Ilex crenata ‘Valeria Rankin’ | Bokrijk | 19900274 | 2.08 ± 0.11 | |
| IL087 | Ilex crenata ‘Variegata’ | Bokrijk | 19980670 | 2.22 ± 0.04 | |
| IL088 | Ilex crenata ‘Fructu Lutea’ | Bokrijk | 19890583 | 2.01 ± 0.07 | |
| IL094 | Ilex vomitoria ‘Folsom Weeping’ | Bokrijk | 20200779 | 1.57 ± 0.10 | |
| IL095 | Ilex vomitoria ‘Hoskins shadow’ | Bokrijk | 20110310 | 1.58 ± 0.06 | |
| IL096 | Ilex sugerokii | Bokrijk | 19980060 | 1.78 ± 0.07 | |
| IL097 | Ilex sugerokii var. longipedunculata | Bokrijk | 19900505 | 1.75 ± 0.02 | |
| IL098 | Ilex rugosa | Bokrijk | 20070853 | 1.97 ± 0.04 | 40 |
| IL099 | Ilex leucoclada | Bokrijk | 20020979 | 2.00 ± 0.05 | 40 |
| IL100 | Ilex opaca | Bokrijk | 19940225 | 1.94 ± 0.02 | |
| IL101 | Ilex serrata ‘Rakusogu’ | Bokrijk | 0000ACQI10 | 1.86 ± 0.07 | |
| IL102 | Ilex integra | Bokrijk | 20120839 | 1.81 ± 0.04 | |
| IL103 | Ilex macrocarpa | Bokrijk | 19960445 | ||
| IL104 | Ilex yunnanensis var. gentilis | Bokrijk | 00004515 | 8.01 ± 0.18 | 160 |
| IL105 | Ilex crenata ‘Jeran’ (Bokrijk select) | Bokrijk | 20140097 | 2.15 ± 0.07 | |
| IL106 | Ilex crenata ‘Convexa Variegata’ | Bokrijk | 19900459 | 2.17 ± 0.05 | |
| IL107 | Ilex crenata ‘Nummularia’ | Bokrijk | 19991312 | 2.23 ± 0.08 | 40 |
| IL108 | Ilex vomitoria ‘Will Fleming’ | Bokrijk | 20080892 | 1.50 ± 0.03 | |
| IL121 | Ilex leucoclada seedling | Bokrijk | |||
| IL122 | Ilex crenata ‘Luxus’ | Plant Select | 2.15 ± 0.08 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geukens, E.; Haegeman, A.; Van Meulder, J.; Van Laere, K.; Smolders, E.; Ruttink, T.; Leus, L. Exploring Genetic Diversity in an Ilex crenata Breeding Germplasm. Horticulturae 2023, 9, 485. https://doi.org/10.3390/horticulturae9040485
Geukens E, Haegeman A, Van Meulder J, Van Laere K, Smolders E, Ruttink T, Leus L. Exploring Genetic Diversity in an Ilex crenata Breeding Germplasm. Horticulturae. 2023; 9(4):485. https://doi.org/10.3390/horticulturae9040485
Chicago/Turabian StyleGeukens, Esther, Annelies Haegeman, Jef Van Meulder, Katrijn Van Laere, Erik Smolders, Tom Ruttink, and Leen Leus. 2023. "Exploring Genetic Diversity in an Ilex crenata Breeding Germplasm" Horticulturae 9, no. 4: 485. https://doi.org/10.3390/horticulturae9040485