
Auxin Participates in the Regulation of the Antioxidant System in Malus baccata Borkh. Roots under Sub-Low Temperature by Exogenous Sucrose Application
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Experimental Treatments
2.2. Measurement of Endogenous IAA Contents
2.3. Measurement of Sucrose, Glucose, Fructose, and Sorbitol Contents
2.4. Measurement of MDA and ROS Contents and ROS Staining of Roots
2.5. Measurement of Nonenzymatic Metabolites and Antioxidative Enzyme Activities
2.6. Measurement of G6PDH Activity
2.7. Measurement of NADPH and NADP+ Contents
2.8. Morphometric Index Determination
2.9. Data Statistics and Analysis
3. Results
3.1. Changes of Growth Parameters
3.2. Changes of Endogenous IAA Levels
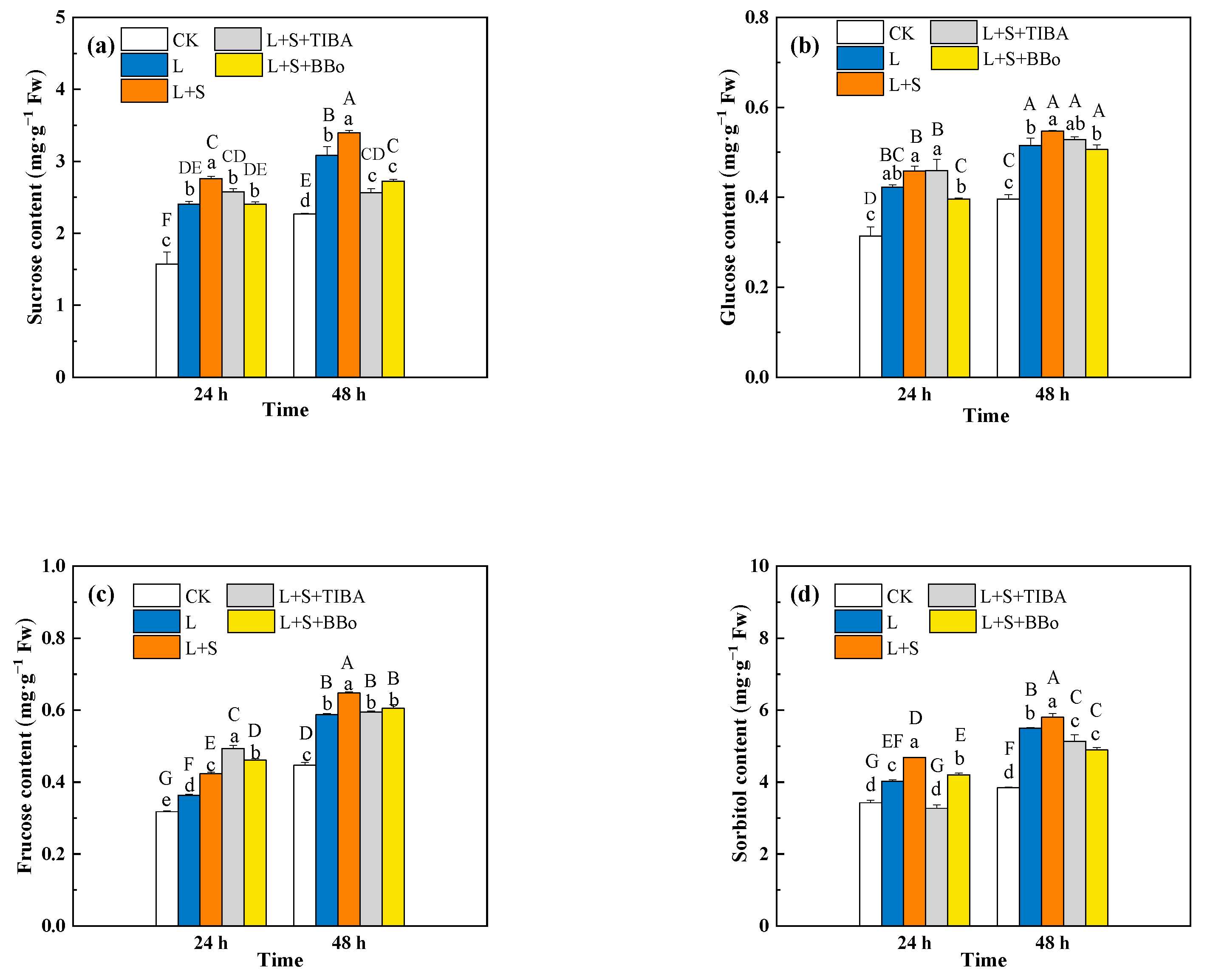
3.3. Changes of Sucrose, Glucose, Fructose, and Sorbitol Contents
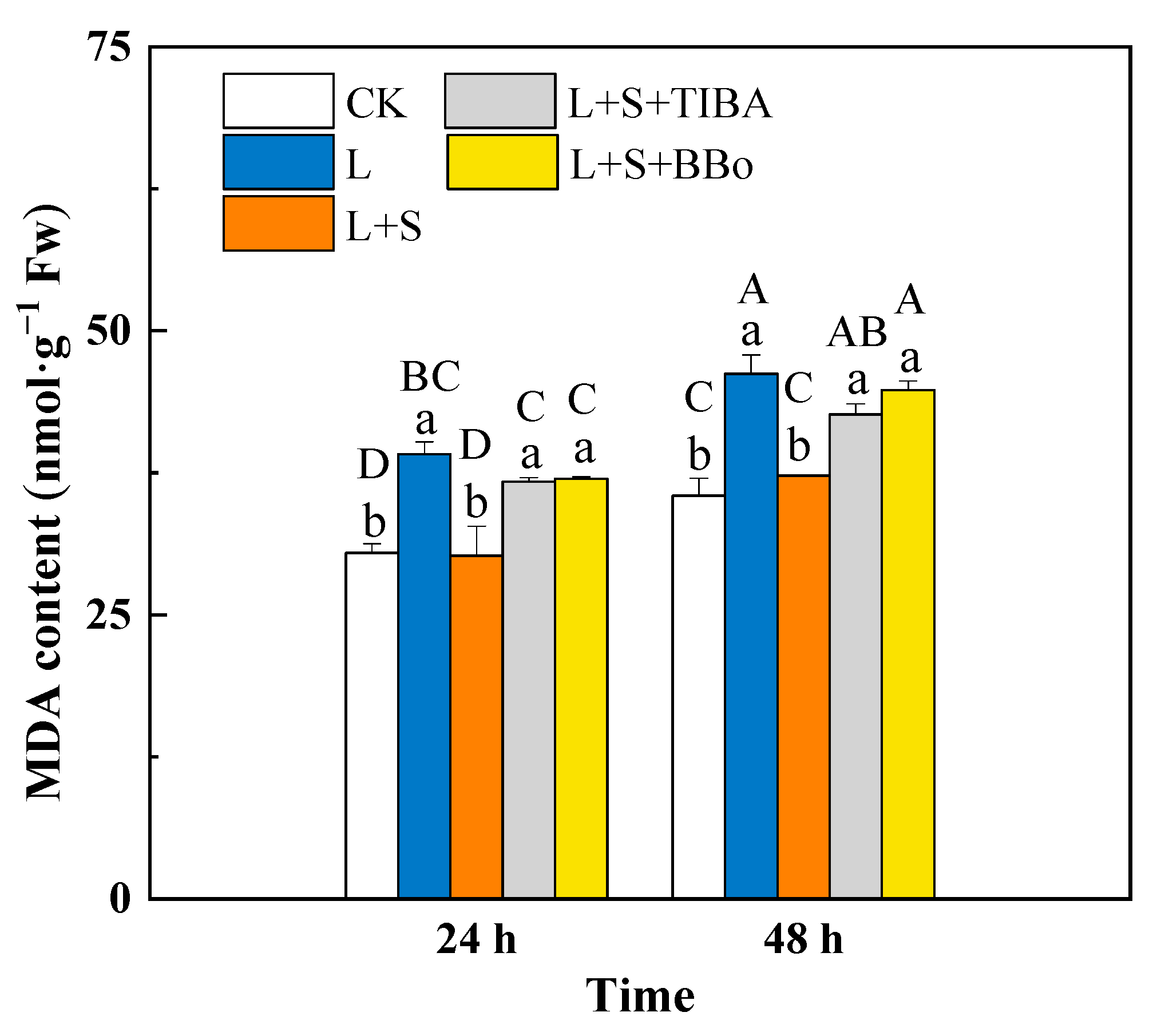
3.4. Changes of MDA Contents
3.5. Changes of ROS Contents
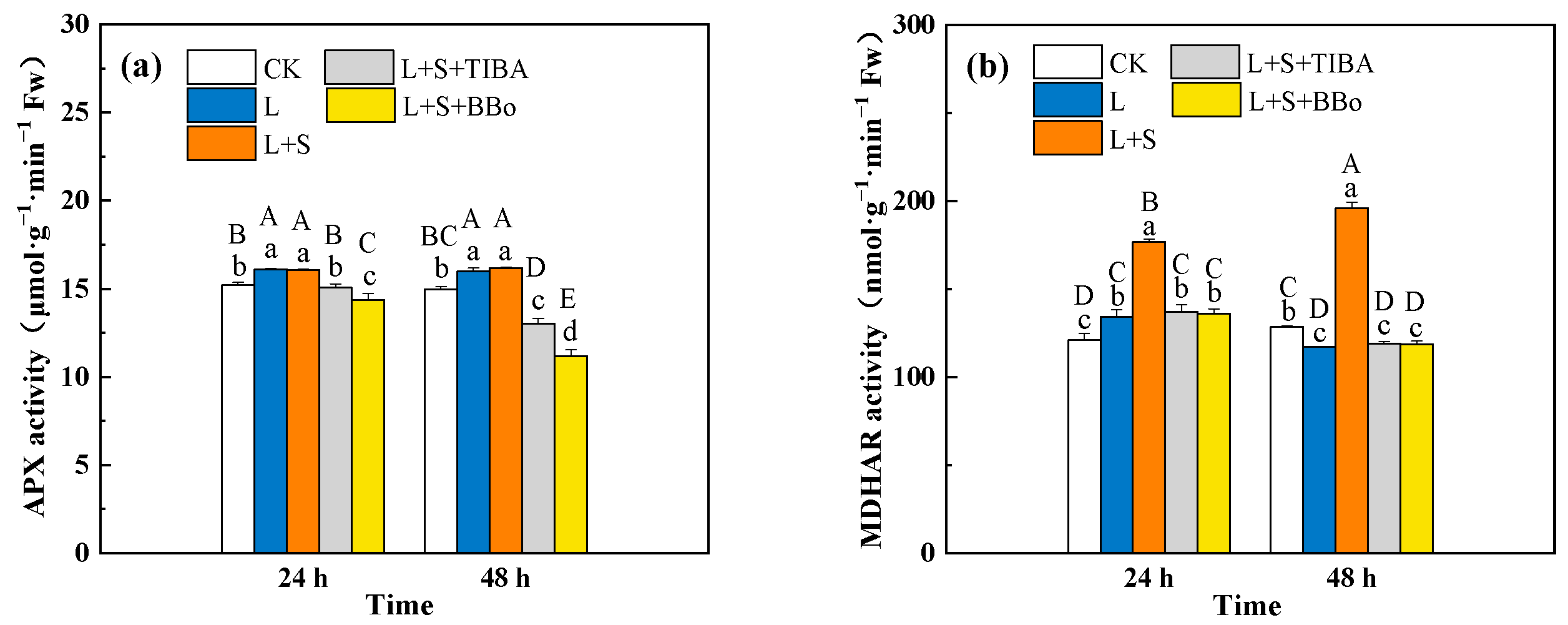
3.6. Changes of SOD, POD, and CAT Activities
3.7. Changes of the AsA-GSH Cycle
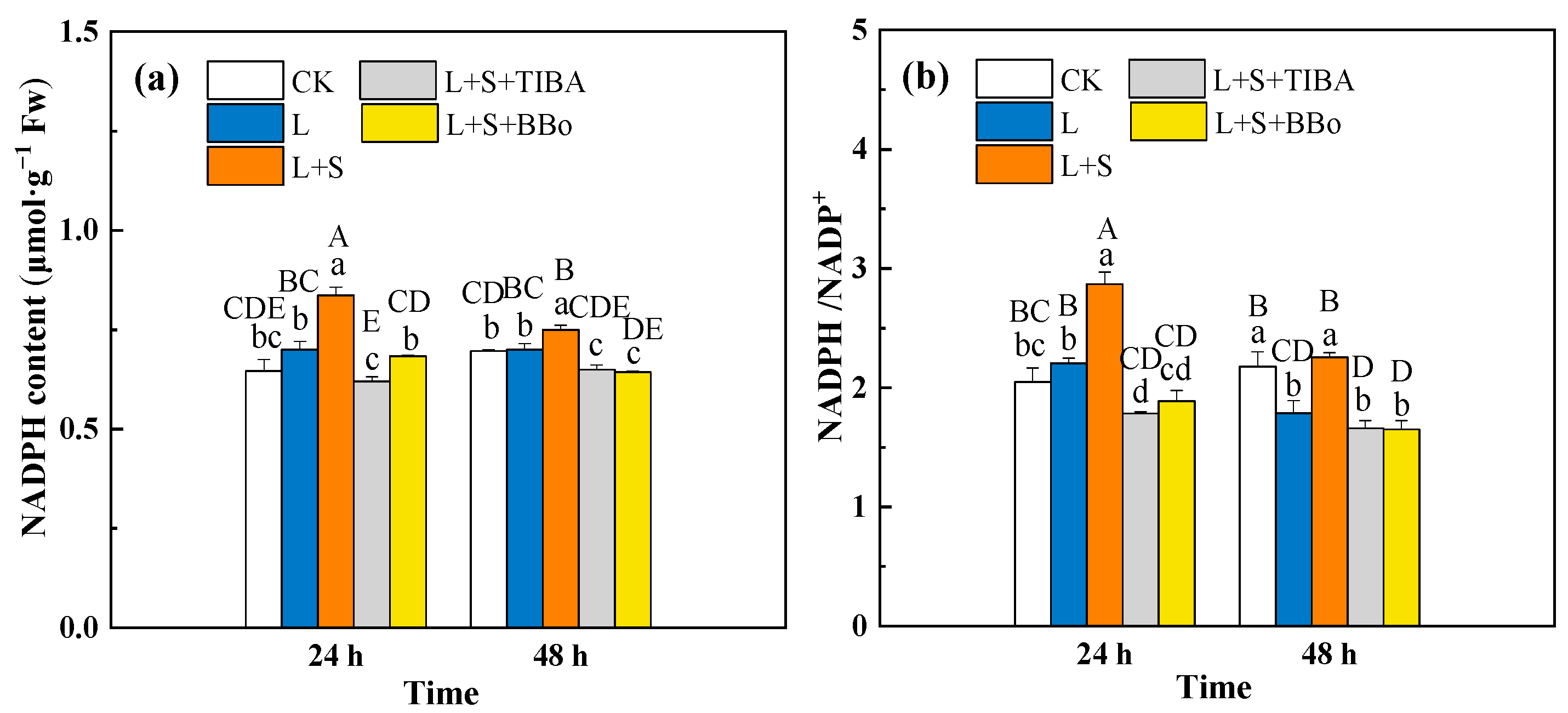
3.8. Changes of G6PDH Activity and NADPH Contents
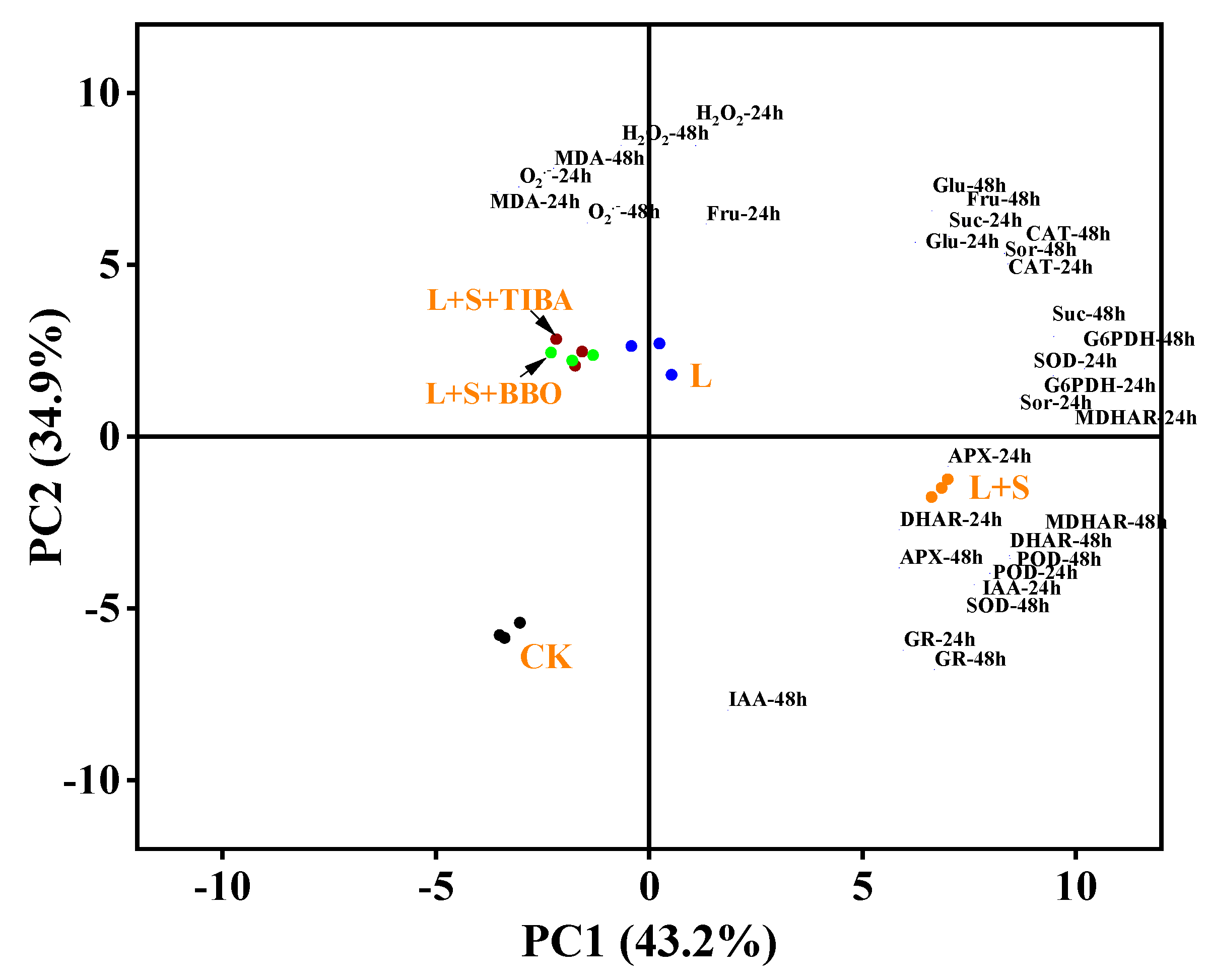
3.9. Principal Component Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jha, U.C.; Bohra, A.; Jha, R. Breeding approaches and genomics technologies to increase crop yield under low-temperature stress. Plant Cell Rep. 2017, 36, 1–35. [Google Scholar] [CrossRef] [PubMed]
- Thomashow, M.F. Plant cold acclimation: Freezing tolerance genes and regulatory mechanisms. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 571–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knight, M.R.; Knight, H. Low-temperature perception leading to gene expression and cold tolerance in higher plants. New Phytol. 2012, 195, 737–751. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Jia, F.F.; Zhang, X.M.; Qiao, Y.X.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Temperature effects on the reactive oxygen species formation and antioxidant defence in roots of two cucurbit species with contrasting root zone temperature optima. Acta Physiol. Plant. 2012, 34, 713–720. [Google Scholar] [CrossRef]
- Jan, N.; Majeed, U.; Andrabi, K.I.; John, R. Cold stress modulates osmolytes and antioxidant system in Calendula officinalis. Acta Physiol. Plant. 2018, 40, 73. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, X.; Deng, F.; Yuan, R.; Shen, F. Genome-wide characterization and expression analyses of superoxide dismutase (SOD) genes in Gossypium hirsutum. BMC Genom. 2017, 18, 376. [Google Scholar] [CrossRef] [Green Version]
- Hosseini, M.; Maali-Amiri, R.; Mahfoozi, S.; Fowler, D.B.; Mohammadi, R. Developmental regulation of metabolites and low temperature tolerance in lines of crosses between spring and winter wheat. Acta Physiol. Plant. 2016, 38, 87. [Google Scholar] [CrossRef]
- Ren, R.; Li, Z.; Zhang, L.; Zhou, H.; Jiang, X.; Liu, Y. Enzymatic and nonenzymatic antioxidant systems impact the viability of cryopreserved Paeonia suffruticosa pollen. Plant Cell Tissue Organ Cult. (PCTOC) 2021, 144, 233–246. [Google Scholar] [CrossRef]
- Shan, C.; Zhou, Y.; Liu, M. Nitric oxide participates in the regulation of the ascorbate-glutathione cycle by exogenous jasmonic acid in the leaves of wheat seedlings under drought stress. Protoplasma 2015, 252, 1397–1405. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Zuo, T.; Ni, W. Important roles of glycinebetaine in stabilizing the structure and function of the photosystem II complex under abiotic stresses. Planta 2020, 251, 36. [Google Scholar] [CrossRef] [Green Version]
- Goyal, M.; Kaur, N. Low temperature induced oxidative stress tolerance in oats (Avena sativa L.) genotypes. Indian J. Plant Physiol. 2018, 23, 316–324. [Google Scholar] [CrossRef]
- Li, L.; Lu, X.; Ma, H.; Lyu, D. Comparative proteomic analysis reveals the roots response to low root-zone temperature in Malus baccata. J. Plant Res. 2018, 131, 865–878. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, S.; Meguro-Maoka, A.; Yoshida, M. Analysis of sugar content and expression of sucrose transporter genes (OsSUTs) in rice tissues in response to a chilling temperature. Jpn. Agric. Res. Q. 2017, 51, 137–146. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Gong, M.; Xin, H.; Tang, L.; Dai, D.; Gao, Y.; Liu, C. Effects of chilling stress on the accumulation of soluble sugars and their key enzymes in Jatropha curcas seedlings. Physiol. Mol. Biol. Plants 2018, 24, 857–865. [Google Scholar] [CrossRef] [PubMed]
- Bolouri-Moghaddam, M.R.; Le Roy, K.; Xiang, L.; Rolland, F.; Van den Ende, W. Sugar signalling and antioxidant network connections in plant cells. FEBS J. 2010, 277, 2022–2037. [Google Scholar] [CrossRef]
- Van den Ende, W.; Valluru, R. Sucrose, sucrosyl oligosaccharides, and oxidative stress: Scavenging and salvaging? J. Exp. Bot. 2009, 60, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.Y.; Yang, M.T.; Li, X.; Zhou, Z.Q.; Wang, X.J.; Bai, J.G. Exogenous sucrose increases chilling tolerance in cucumber seedlings by modulating antioxidant enzyme activity and regulating proline and soluble sugar contents. Sci. Hortic. 2014, 179, 67–77. [Google Scholar] [CrossRef]
- Cao, Y.Y.; Yang, M.T.; Chen, S.Y.; Zhou, Z.Q.; Li, X.; Wang, X.J.; Bai, J.G. Exogenous sucrose influences antioxidant enzyme activities and reduces lipid peroxidation in water-stressed cucumber leaves. Biol. Plant. 2015, 59, 147–153. [Google Scholar] [CrossRef]
- Gong, H.L.; Chen, Q.Q. Exogenous sucrose protects potato seedlings against heat stress by enhancing the antioxidant defense system. J. Soil Sci. Plant Nutr. 2021, 21, 1511–1519. [Google Scholar] [CrossRef]
- Wu, P.; Xiao, C.; Cui, J.; Hao, B.; Zhang, W.; Yang, Z.; Ahammed, G.J.; Liu, H.; Cui, H. Nitric oxide and its interaction with hydrogen peroxide enhance plant yolerance to low temperatures by improving the efficiency of the calvin cycle and the ascorbate–glutathione cycle in cucumber seedlings. J. Plant Growth Regul. 2021, 40, 2390–2408. [Google Scholar] [CrossRef]
- Ying, Z. Sugar-induced tolerance to the salt stress in maize seedlings by balancing redox homeostasis. Agric. For. Fish. 2016, 5, 126. [Google Scholar] [CrossRef] [Green Version]
- Wingler, A.; Tijero, V.; Müller, M.; Yuan, B.; Munné-Bosch, S. Interactions between sucrose and jasmonate signalling in the response to cold stress. BMC Plant Biol. 2020, 20, 176. [Google Scholar] [CrossRef]
- Lilley, J.L.; Gee, C.W.; Sairanen, I.; Ljung, K.; Nemhauser, J.L. An endogenous carbon-sensing pathway triggers increased auxin flux and hypocotyl elongation. Plant Physiol. 2012, 160, 2261–2270. [Google Scholar] [CrossRef] [Green Version]
- Sairanen, I.; Novák, O.; Pěnčík, A.; Ikeda, Y.; Jones, B.; Sandberg, G.; Ljung, K. Soluble carbohydrates regulate auxin biosynthesis via PIF proteins in Arabidopsis. Plant Cell 2012, 24, 4907–4916. [Google Scholar]
- Mir, A.R.; Siddiqui, H.; Alam, P.; Hayat, S. Foliar spray of Auxin/IAA modulates photosynthesis, elemental composition, ROS localization and antioxidant machinery to promote growth of Brassica juncea. Physiol. Mol. Biol. Plants 2020, 26, 2503–2520. [Google Scholar] [CrossRef] [PubMed]
- Yemelyanov, V.; Lastochkin, V.; Chirkova, T.; Lindberg, S.; Shishova, M. Indoleacetic acid levels in wheat and rice seedlings under oxygen deficiency and subsequent reoxygenation. Biomolecules 2020, 10, 276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Chen, L.; Ye, T.; Liu, X.; Ding, K.; Chan, Z. Modulation of auxin content in Arabidopsis confers improved drought stress resistance. Plant Physiol. Biochem. 2014, 82, 209–217. [Google Scholar] [CrossRef]
- Khan, M.Y.; Prakash, V.; Yadav, V.; Chauhan, D.K.; Prasad, S.M.; Ramawat, N.; Singh, V.P.; Tripathi, D.K.; Sharma, S. Regulation of cadmium toxicity in roots of tomato by indole acetic acid with special emphasis on reactive oxygen species production and their scavenging. Plant Physiol. Biochem. 2019, 142, 193–201. [Google Scholar] [CrossRef]
- Qi, X.; Li, Q.; Shen, J.; Qian, C.; Xu, X.; Xu, Q.; Chen, X. Sugar enhances waterlogging-induced adventitious root formation in cucumber by promoting auxin transport and signalling. Plant Cell Environ. 2020, 43, 1545–1557. [Google Scholar] [CrossRef]
- Du, F.; Xu, J.N.; Li, D.; Wang, X.Y. The identification of novel and differentially expressed apple-tree genes under low-temperature stress using high-throughput Illumina sequencing. Mol. Biol. Rep. 2015, 42, 569–580. [Google Scholar] [CrossRef]
- Farooq, M.A.; Li, L.; Ali, B.; Gill, R.A.; Wang, J.; Ali, S.; Gill, M.B.; Zhou, W. Oxidative injury and antioxidant enzymes regulation in arsenic-exposed seedlings of four Brassica napus L. cultivars. Environ. Sci. Pollut. Res. 2015, 22, 10699–10712. [Google Scholar] [CrossRef]
- Juszczak, I.; Cvetkovic, J.; Zuther, E.; Hincha, D.K.; Baier, M. Natural variation of cold deacclimation correlates with variation of cold-acclimation of the plastid antioxidant system in Arabidopsis thaliana accessions. Front. Plant Sci. 2016, 7, 305. [Google Scholar] [CrossRef] [Green Version]
- Afzal, J.; Hu, C.; Imtiaz, M.; Elyamine, A.M.; Rana, M.S.; Imran, M.; Farag, M.A. Cadmium tolerance in rice cultivars associated with antioxidant enzymes activities and Fe/Zn concentrations. Int. J. Environ. Sci. Technol. 2019, 16, 4241–4252. [Google Scholar] [CrossRef]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y.; Zheng, X.; Wang, Y. Silicon dioxide nanoparticles improve plant growth by enhancing antioxidant enzyme capacity in bamboo (Pleioblastus pygmaeus) under lead toxicity. Trees 2020, 34, 469–481. [Google Scholar] [CrossRef]
- Maksimović, J.D.; Zhang, J.; Zeng, F.; Živanović, B.D.; Shabala, L.; Zhou, M.; Shabala, S. Linking oxidative and salinity stress tolerance in barley: Can root antioxidant enzyme activity be used as a measure of stress tolerance? Plant Soil 2013, 365, 141–155. [Google Scholar] [CrossRef]
- Wang, J.; Zeng, Q.; Zhu, J.; Liu, G.; Tang, H. Dissimilarity of ascorbate–glutathione (AsA–GSH) cycle mechanism in two rice (Oryza sativa L.) cultivars under experimental free-air ozone exposure. Agric. Ecosyst. Environ. 2013, 165, 39–49. [Google Scholar] [CrossRef]
- Yu, J.; Cang, J.; Li, Y.; Huang, R.; Lu, Q.; Wang, X.; Liu, L.; Xu, Q.; Zhang, K. Salicylic acid-induced antioxidant protection against low temperature in cold-hardy winter wheat. Acta Physiol. Plant. 2016, 38, 261. [Google Scholar] [CrossRef]
- Li, L.; Lu, X.; Ma, H.; Lyu, D. Jasmonic acid regulates the ascorbate–glutathione cycle in Malus baccata Borkh. roots under low root-zone temperature. Acta Physiol. Plant. 2017, 39, 174. [Google Scholar] [CrossRef]
- Castiglia, D.; Cardi, M.; Landi, S.; Cafasso, D.; Esposito, S. Expression and characterization of a cytosolic glucose 6 phosphate dehydrogenase isoform from barley (Hordeum vulgare) roots. Protein Expr. Purif. 2015, 112, 8–14. [Google Scholar] [CrossRef]
- Tian, Y.; Peng, K.; Bao, Y.; Zhang, D.; Meng, J.; Wang, D.; Wang, X.; Cang, J. Glucose-6-phosphate dehydrogenase and 6-phosphogluconate dehydrogenase genes of winter wheat enhance the cold tolerance of transgenic Arabidopsis. Plant Physiol. Biochem. 2021, 161, 86–97. [Google Scholar] [CrossRef]
- Jung, H.i.; Lee, B.R.; Chae, M.J.; Lee, E.J.; Lee, T.G.; Jung, G.B.; Kim, M.S.; Lee, J. Ascorbate-mediated modulation of cadmium stress responses: Reactive oxygen species and redox status in brassica napus. Front. Plant Sci. 2020, 11, 586547. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A. Auxin: A regulator of cold stress response. Physiol Plant. 2013, 147, 28–35. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Zhang, K.; Wang, W.; Gong, W.; Liu, W.; Chen, H.; Xu, H.; Lu, Y. Low temperature inhibits root growth by reducing auxin accumulation via ARR1/12. Plant Cell Physiol. 2015, 56, 727–736. [Google Scholar] [CrossRef] [Green Version]
- Barbier, F.; Peron, T.; Lecerf, M.; Perez-Garcia, M.D.; Barriere, Q.; Rolcik, J.; Boutet-Mercey, S.; Citerne, S.; Lemoine, R.; Porcheron, B.; et al. Sucrose is an early modulator of the key hormonal mechanisms controlling bud outgrowth in Rosa hybrida. J. Exp. Bot. 2015, 66, 2569–2582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meitzel, T.; Radchuk, R.; McAdam, E.; Thormählen, I.; Feil, R.; Munz, E.; Hilo, A.; Geigenberger, P.; Ross, J.; Lunn, J. Trehalose 6-phosphate promotes seed filling by activating auxin biosynthesis. New Phytol. 2021, 229, 1553–1565. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L.; Dalal, M.; Verma, R.K.; Kumar, S.V.V.; Yadav, S.K.; Pushkar, S.; Kushwaha, S.R.; Bhowmik, A.; Chinnusamy, V. Auxin protects spikelet fertility and grain yield under drought and heat stresses in rice. Environ. Exp. Bot. 2018, 150, 9–24. [Google Scholar] [CrossRef]
- Zhang, S.; Peng, F.; Xiao, Y.; Wang, W.; Wu, X. Peach PpSnRK1 participates in sucrose-mediated root growth through auxin signaling. Front. Plant Sci. 2020, 11, 409. [Google Scholar] [CrossRef]
- Zhang, X.W.; Liu, F.J.; Zhai, J.; Li, F.D.; Bi, H.G.; Ai, X.Z. Auxin acts as a downstream signaling molecule involved in hydrogen sulfide-induced chilling tolerance in cucumber. Planta 2020, 251, 69. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time | CK | L | L + S | L + S + TIBA | L + S + BBo |
|---|---|---|---|---|---|
| 8: 00 | Distilled water | Distilled water | Distilled water | TIBA | BBo |
| 16: 00 | Distilled water | Distilled water | Sucrose | Sucrose | Sucrose |
| Treatments | Plant Height | Stem Diameter | Total Biomass | Root Biomass | Shoot Biomass | S:R Ratio |
|---|---|---|---|---|---|---|
| (cm) | (mm) | (g Dw *) | (g Dw *) | (g Dw *) | ||
| CK | 43.13 ± 1.59 a | 4.77 ± 0.11 a | 7.30 ± 0.12 a | 2.34 ± 0.16 a | 4.96 ± 0.24 a | 2.15 ± 0.23 a |
| L | 38.00 ± 1.17 c | 4.62 ± 0.05 a | 6.18 ± 0.09 b | 2.20 ± 0.01 a | 3.98 ± 0.08 b | 1.81 ± 0.03 ab |
| L + S | 43.33 ± 0.60 a | 4.63 ± 0.04 a | 6.87 ± 0.06 a | 2.34 ± 0.01 a | 4.53 ± 0.05 a | 1.94 ± 0.02 a |
| L + S + TIBA | 39.13 ± 1.25 b | 4.52 ± 0.07 a | 5.51 ± 0.31 c | 2.30 ± 0.07 a | 3.21 ± 0.27 c | 1.39 ± 0.11 b |
| L + S + BBo | 39.60 ± 0.17 b | 4.51 ± 0.07 a | 5.79 ± 0.12 bc | 2.09 ± 0.16 a | 3.70 ± 0.09 bc | 1.80 ± 0.17 ab |
| Treatments | Total Root Length | Root Surface Area | Root Volume | Root Average Diameter |
|---|---|---|---|---|
| (cm) | (cm2) | (cm3) | (mm) | |
| CK | 2604.15 ± 158.67 ab | 392.63 ± 28.48 a | 4.16 ± 0.25 a | 0.48 ± 0.01 a |
| L | 2248.48 ± 103.54 bc | 326.84 ± 9.39 b | 3.80 ± 0.34 ab | 0.46 ± 0.01 ab |
| L + S | 2711.92 ± 170.54 a | 380.23 ± 18.45 ab | 3.91 ± 0.30 ab | 0.46 ± 0.01 ab |
| L + S + TIBA | 1988.79 ± 51.13 c | 249.09 ± 23.32 c | 3.05 ± 0.16 bc | 0.43 ± 0.02 b |
| L + S + BBo | 1986.72 ± 70.80 c | 221.15 ± 23.02 c | 2.70 ± 0.28 c | 0.43 ± 0.01 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Yang, B.; Zhao, X.; Wang, P.; Lyu, D.; Qin, S. Auxin Participates in the Regulation of the Antioxidant System in Malus baccata Borkh. Roots under Sub-Low Temperature by Exogenous Sucrose Application. Horticulturae 2023, 9, 297. https://doi.org/10.3390/horticulturae9030297
Li L, Yang B, Zhao X, Wang P, Lyu D, Qin S. Auxin Participates in the Regulation of the Antioxidant System in Malus baccata Borkh. Roots under Sub-Low Temperature by Exogenous Sucrose Application. Horticulturae. 2023; 9(3):297. https://doi.org/10.3390/horticulturae9030297
Chicago/Turabian StyleLi, Lijie, Beilei Yang, Xiaoping Zhao, Peng Wang, Deguo Lyu, and Sijun Qin. 2023. "Auxin Participates in the Regulation of the Antioxidant System in Malus baccata Borkh. Roots under Sub-Low Temperature by Exogenous Sucrose Application" Horticulturae 9, no. 3: 297. https://doi.org/10.3390/horticulturae9030297