
Changes in Cell Wall Sugar Neutral Composition Contribute to Apple Texture Loss during Storage among Cultivars

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fruit Material
2.2. Apple Fruit Texture Puncture Test and Compression Observation
2.3. Fruit Physiological Property Indicators
2.3.1. Determination of Ethylene Production Rate, Respiration Rate, and Pectin Content
2.3.2. Preparation of CWM and Determination of Neutral Sugar Composition
2.4. Screening of Candidate Cell Wall-Related Genes in the Apple Genome
2.5. RNA Extraction, cDNA Synthesis, and Real-Time PCR Quantification
2.6. Statistical Analysis
3. Results
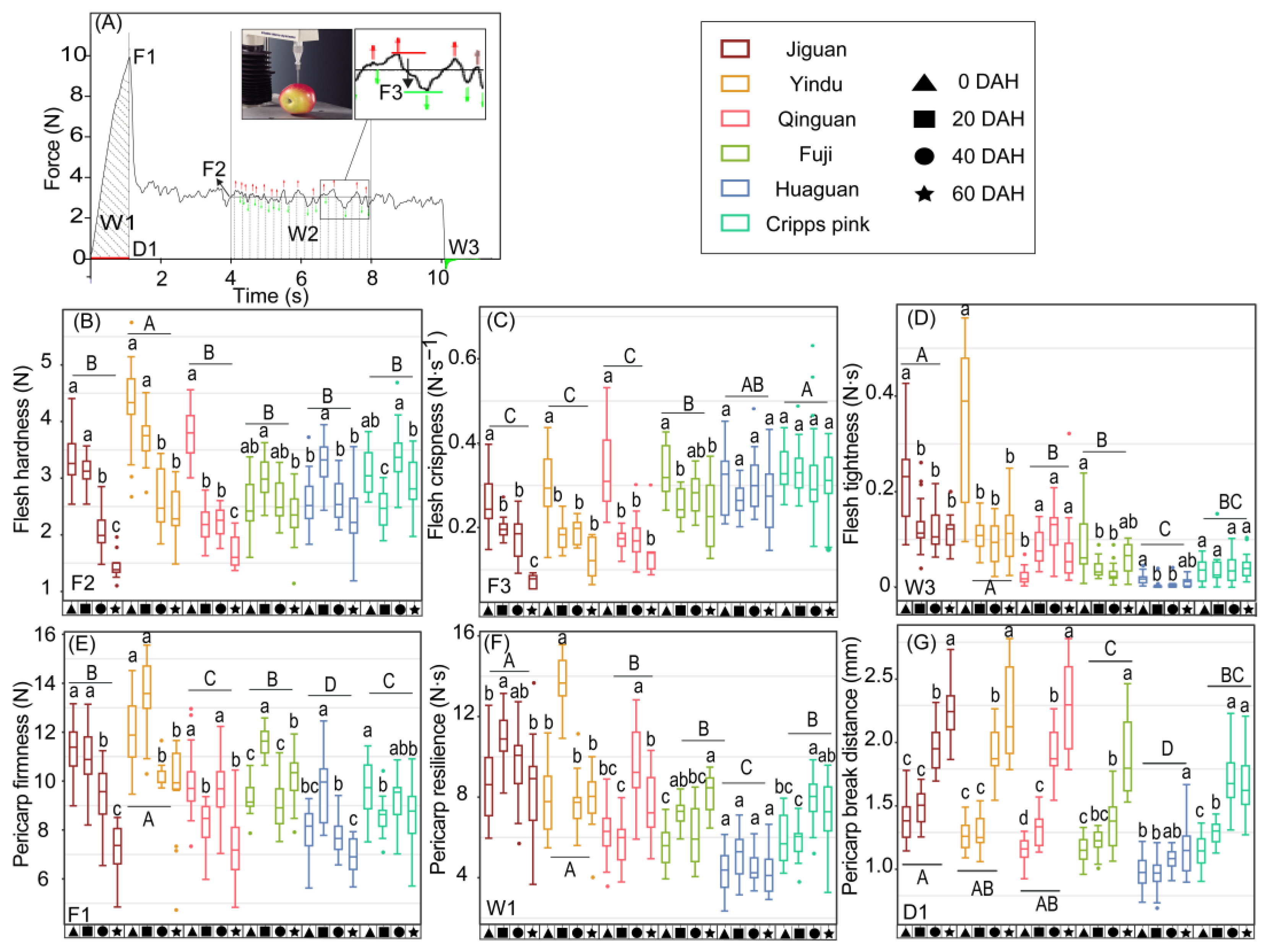
3.1. A Textural Evaluation System of Apple Fruit in Mechanical and Sensorial Ways
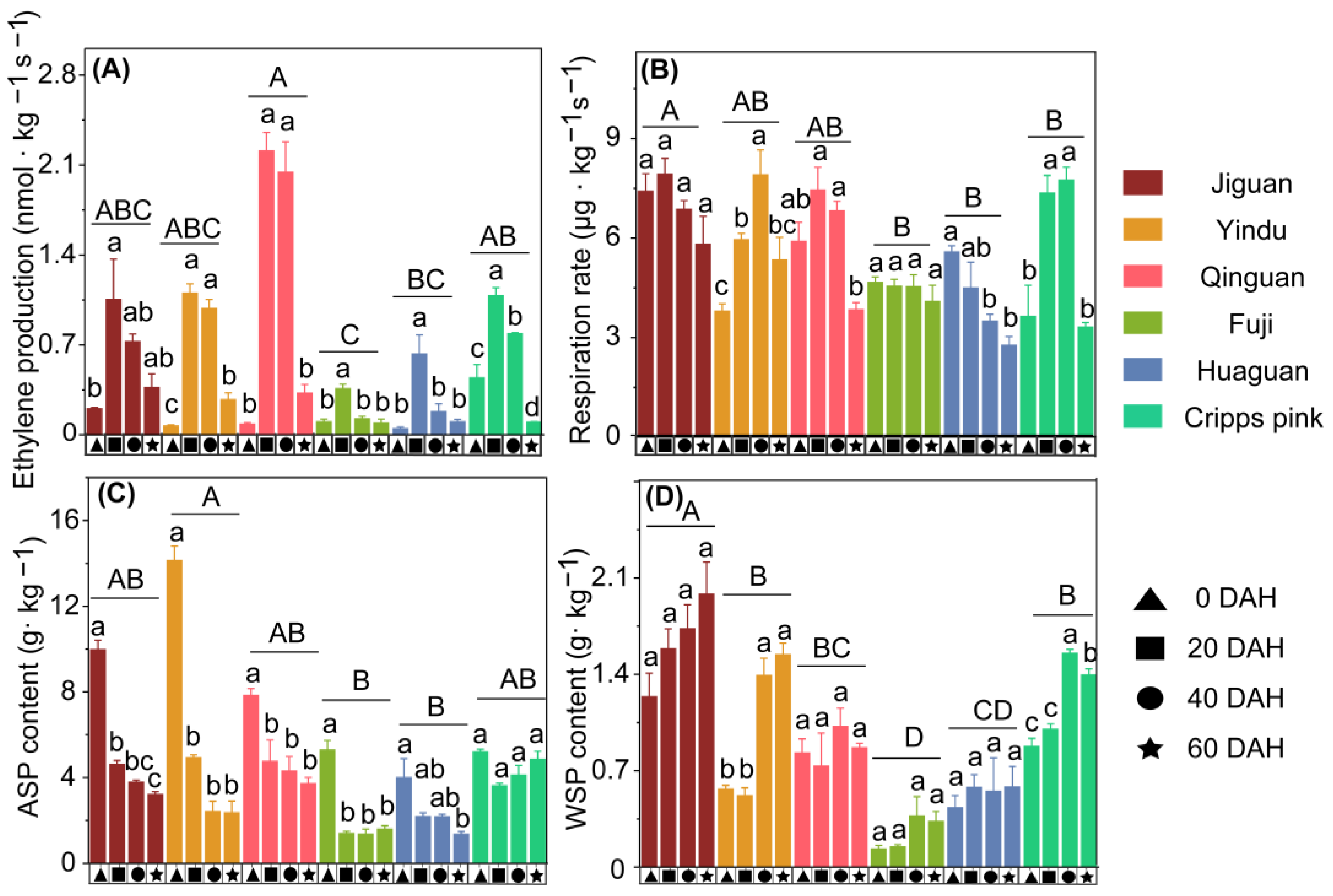
3.2. Fruit Postharvest Physiological Characteristics of Six Apple Cultivars
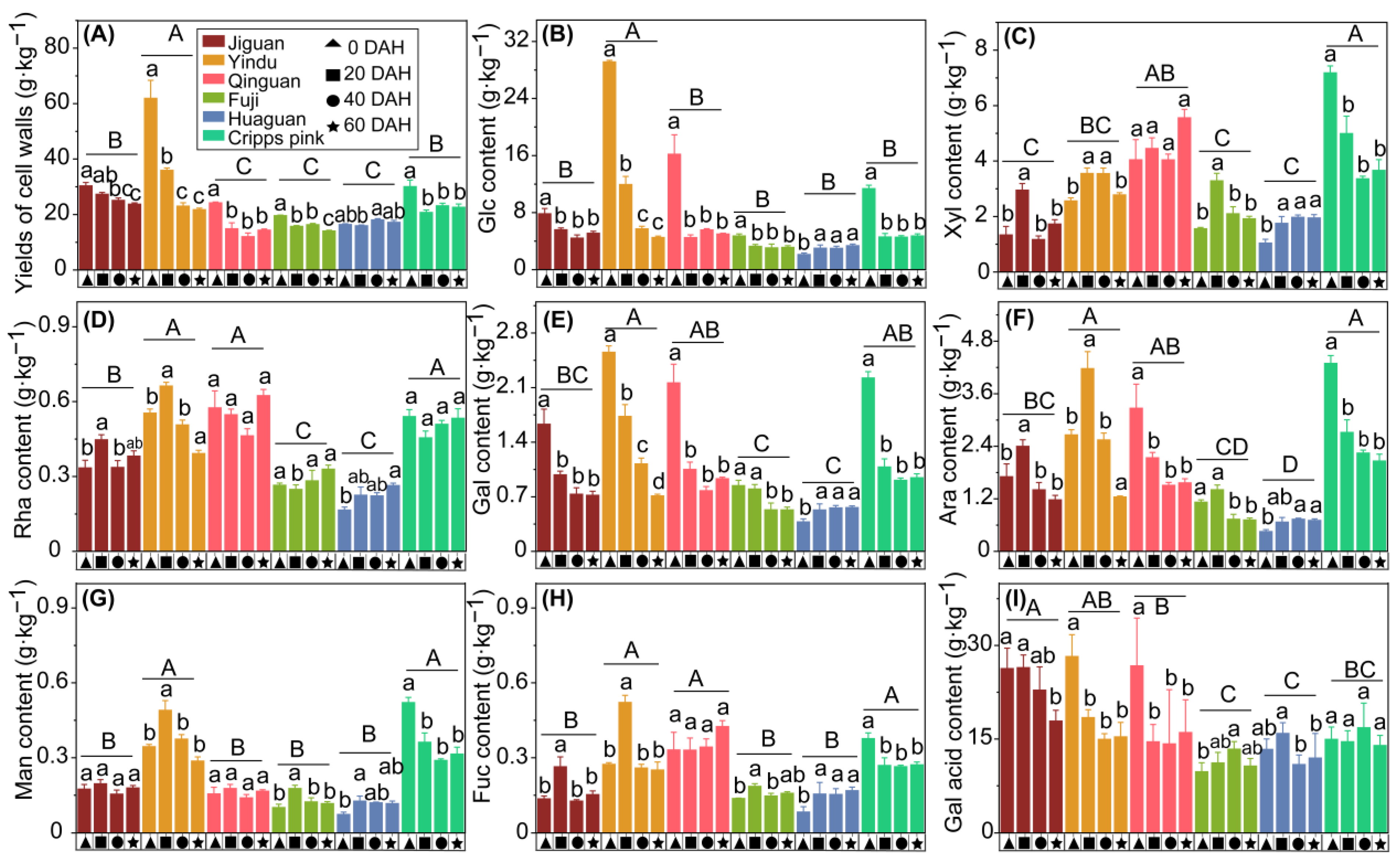
3.3. Cell Wall Material and Its Neutral Sugar Composition Postharvest Changes of Six Apple Cultivars
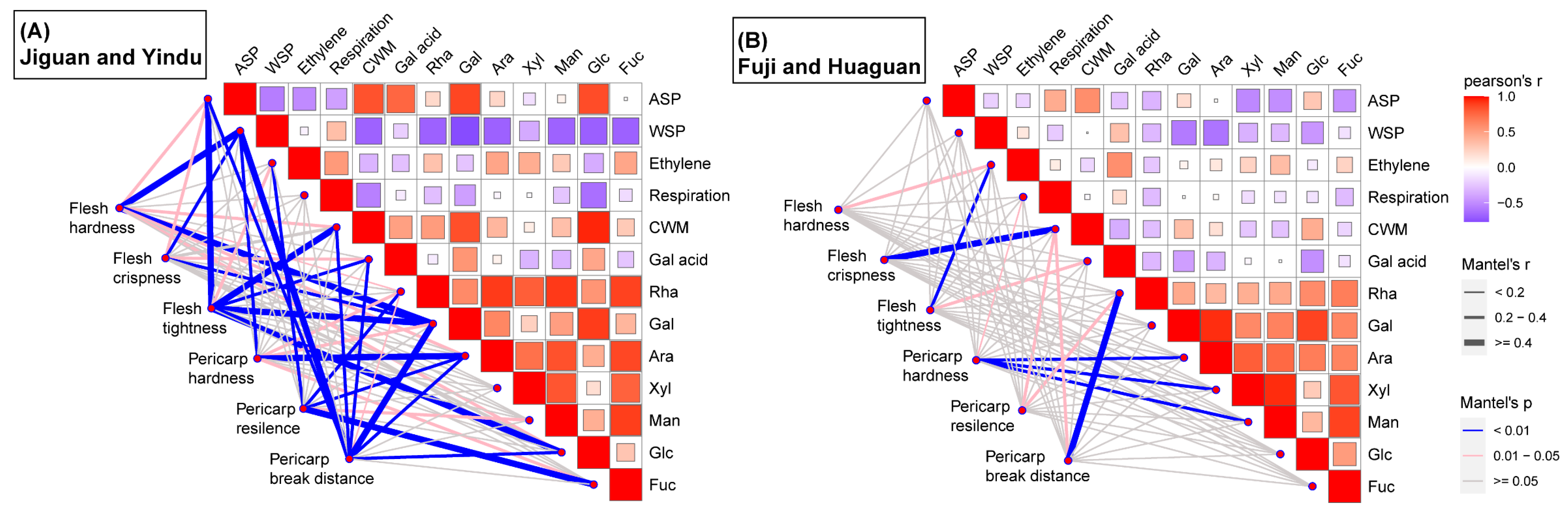
3.4. Correlation of Cultivar Differences in Apple Fruit Texture during Postharvest
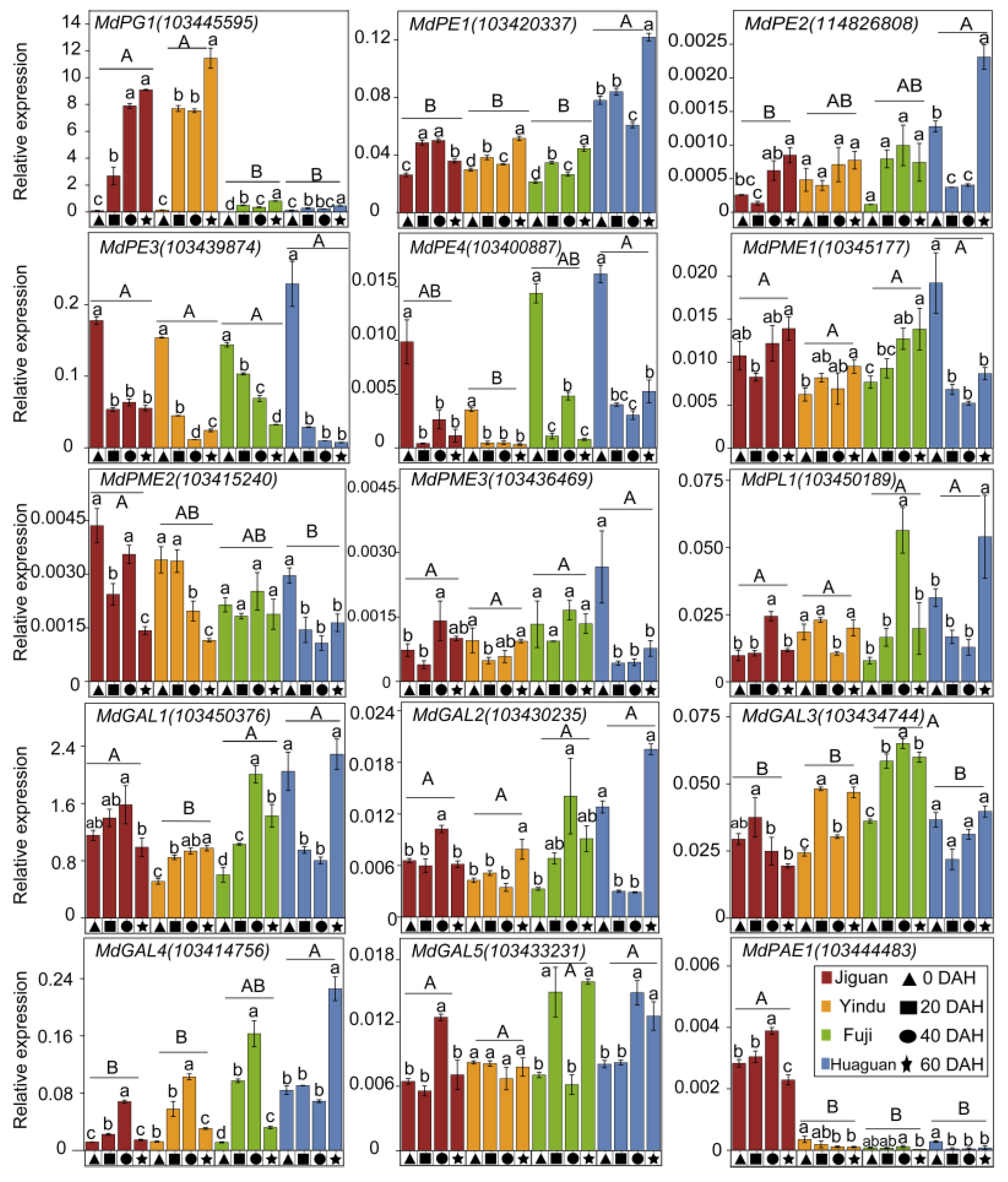
3.5. Cell Wall Degradation-Related Genes Involved in Textural Changes of Apple Fruit during Postharvest Storage
4. Discussion
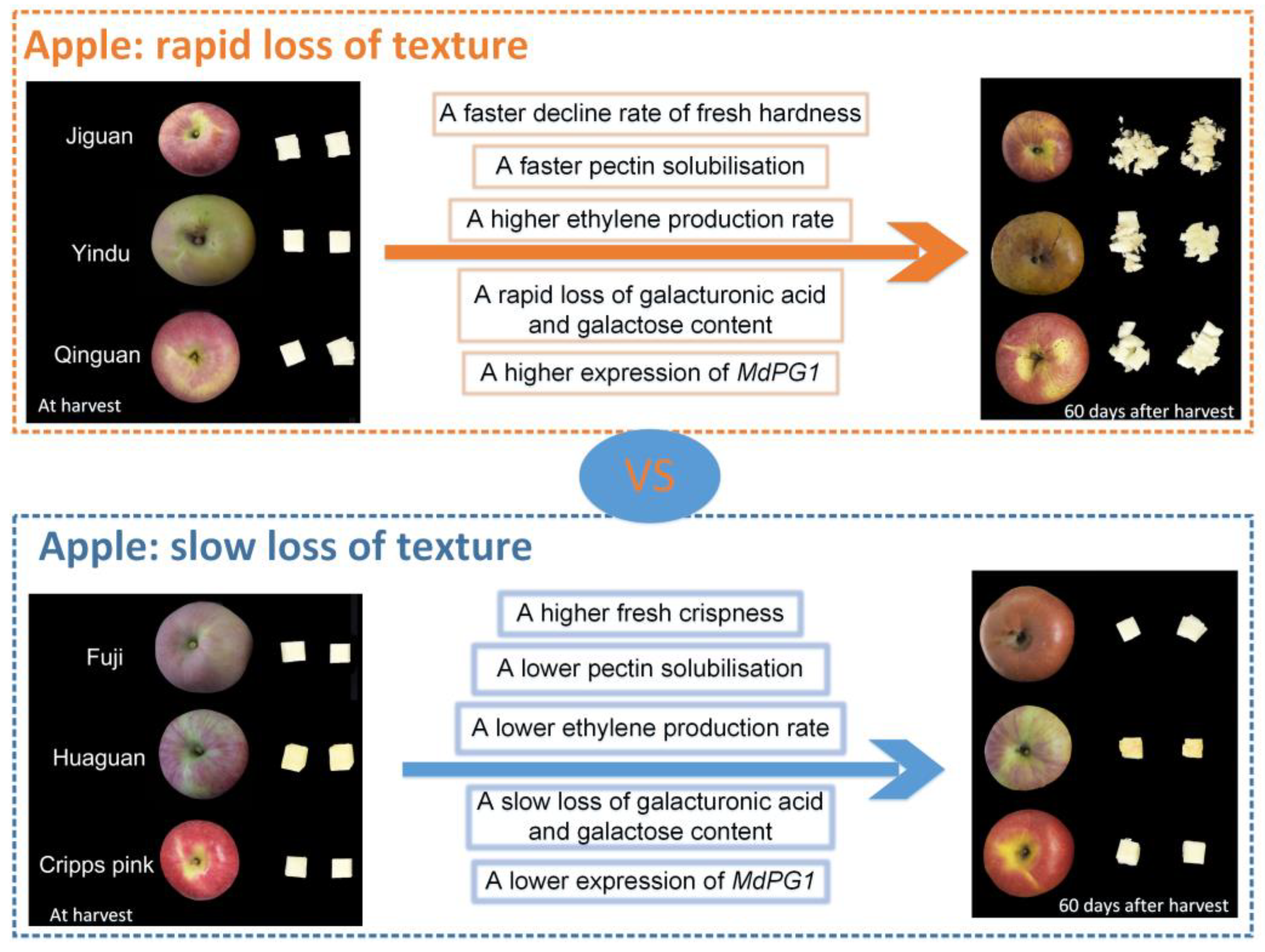
4.1. The Difference in Texture Loss Rate among the Six Cultivars Was Mainly Manifested in the Flesh Hardness and Crispness of Apple
4.2. Cell Wall Neutral Sugar Composition Changes May Contribute to the Apple Texture Loss of Different Cultivars during Storage
4.2.1. The Rapid Loss of Galacturonic Acid Content May Result in the Rapid Loss of Apple Texture
4.2.2. The Rapid Loss of Galactose Content Could Be Responsible for the Rapid Loss of Apple Texture
4.3. A Higher Peak Ethylene Production Rate May Lead to More ASP Translated to WSP and More Cell Wall Neutral Sugar Composition Changes, Resulting in Apple Texture Loss
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mureșan, A.E.; Sestras, A.F.; Militaru, M.; Păucean, A.; Tanislav, A.E.; Pușcaș, A.; Mateescu, M.; Mureșan, V.; Marc, R.A.; Sestras, R.E. Chemometric comparison and classification of 22 apple genotypes based on texture analysis and physico-chemical quality attributes. Horticulturae 2022, 8, 64. [Google Scholar] [CrossRef]
- Rojas-Candelas, L.E.; Chanona-Pérez, J.J.; Méndez Méndez, J.V.; Perea-Flores, M.J.; Cervantes-Sodi, H.F.; Hernández-Hernández, H.M.; Marin-Bustamante, M.Q. Physicochemical, structural and nanomechanical study elucidating the differences in firmness among four apple cultivars. Postharvest Biol. Technol. 2021, 171, 111342. [Google Scholar] [CrossRef]
- Ni, F.; Meng, Q.; Gu, F.; Hu, Y. Building kinetic models for apple crispness to determine the optimal freshness preservation time during shelf life based on spectroscopy. J. Food Process. Preserv. 2020, 44, e14422. [Google Scholar] [CrossRef]
- Trujillo, D.I.; Mann, H.S.; Tong, C.B.S. Examination of Expansin genes as related to apple fruit crispness. Tree Genet. Genomes 2012, 8, 27–38. [Google Scholar] [CrossRef]
- Li, Q.; Xu, R.; Fang, Q.; Yuan, Y.; Cao, J.; Jiang, W. Analyses of microstructure and cell wall polysaccharides of flesh tissues provide insights into cultivar difference in mealy patterns developed in apple fruit. Food Chem. 2020, 321, 126707. [Google Scholar] [CrossRef]
- Segonne, S.M.; Bruneau, M.; Celton, J.M.; Le Gall, S.; Francin-Allami, M.; Juchaux, M.; Laurens, F.; Orsel, M.; Renou, J.P. Multiscale investigation of mealiness in apple: An atypical role for a pectin methylesterase during fruit maturation. BMC Plant Biol. 2014, 14, 375. [Google Scholar] [CrossRef] [Green Version]
- Brookfield, P.L.; Nicoll, S.; Gunson, F.A.; Harker, F.R.; Wohlers, M. Sensory evaluation by small postharvest teams and the relationship with instrumental measurements of apple texture. Postharvest Biol. Technol. 2011, 59, 179–186. [Google Scholar] [CrossRef]
- Johnston, J.W.; Hewett, E.W.; Hertog, M.L.A.T.M. Postharvest softening of apple (Malus domestica) fruit: A Review. N. Z. J. Crop Hortic. Sci. 2002, 30, 145–160. [Google Scholar] [CrossRef]
- Kwon, Y.S.; Kwon, S.-I.; Kim, J.-H.; Park, M.Y.; Park, J.T.; Kim, S.A. Validation assay of Md-ACS1, Md-ACO1, and Md-PG1 molecular markers associated with storability in apples. Korean J. Breed. Sci. 2020, 52, 322–331. [Google Scholar] [CrossRef]
- Tacken, E.; Ireland, H.; Gunaseelan, K.; Karunairetnam, S.; Wang, D.; Schultz, K.; Bowen, J.; Atkinson, R.G.; Johnston, J.W.; Putterill, J.; et al. The role of ethylene and cold temperature in the regulation of the apple polygalacturonase1 gene and fruit softening. Plant Physiol. 2010, 153, 294–305. [Google Scholar] [CrossRef] [Green Version]
- Péneau, S.; Brockhoff, P.B.; Hoehn, E.; Escher, F.; Nuessli, J.; Plads, R.P. Relating consumer evaluation of apple freshness to sensory and physico-chemical measurements. J. Sensory Stud. 2006, 22, 313–335. [Google Scholar] [CrossRef]
- Costa, F.; Cappellin, L.; Fontanari, M.; Longhi, S.; Guerra, W.; Magnago, P.; Gasperi, F.; Biasioli, F. Texture dynamics during postharvest cold storage ripening in apple (Malus × domestica Borkh.). Postharvest Biol. Technol. 2012, 69, 54–63. [Google Scholar] [CrossRef]
- Costa, F.; Cappellin, L.; Longhi, S.; Guerra, W.; Magnago, P.; Porro, D.; Soukoulis, C.; Salvi, S.; Velasco, R.; Biasioli, F.; et al. Assessment of apple (Malus × domestica Borkh.) Fruit texture by a combined acoustic-mechanical profiling strategy. Postharvest Biol. Technol. 2011, 61, 21–28. [Google Scholar] [CrossRef]
- Yang, L.; Cong, P.; He, J.; Bu, H.; Qin, S.; Lyu, D. Differential pulp cell wall structures lead to diverse fruit textures in apple (Malus domestica). Protoplasma 2022, 259, 1205–1217. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Quan, P.; Liu, H.; Li, L.; Qi, S.; Zhang, M.; Zhang, B.; Li, H.; Zhao, Y.; Ma, B.; et al. Transcriptomic and metabolic analyses provide new insights into the apple fruit quality decline during long-term cold storage. J. Agric. Food Chem. 2020, 68, 4699–4716. [Google Scholar] [CrossRef] [PubMed]
- Billy, L.; Mehinagic, E.; Royer, G.; Renard, C.M.G.C.; Arvisenet, G.; Prost, C.; Jourjon, F. Relationship between texture and pectin composition of two apple cultivars during storage. Postharvest Biol. Technol. 2008, 47, 315–324. [Google Scholar] [CrossRef]
- Tucker, G.; Yin, X.; Zhang, A.; Wang, M.; Zhu, Q.; Liu, X.; Xie, X.; Chen, K.; Grierson, D. Ethylene and fruit softening. Food Qual. Saf. 2017, 1, 253–267. [Google Scholar] [CrossRef]
- Caffall, K.H.; Mohnen, D. The structure, function, and biosynthesis of plant cell wall pectic polysaccharides. Carbohydr. Res. 2009, 344, 1879–1900. [Google Scholar] [CrossRef] [PubMed]
- Voragen, A.G.J.; Coenen, G.-J.; Verhoef, R.P.; Schols, H.A. Pectin, a versatile polysaccharide present in plant cell walls. Struct. Chem. 2009, 20, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Lahaye, M.; Bouin, C.; Barbacci, A.; Le Gall, S.; Foucat, L. Water and cell wall contributions to apple mechanical properties. Food Chem. 2018, 268, 386–394. [Google Scholar] [CrossRef]
- Ireland, H.S.; Gunaseelan, K.; Muddumage, R.; Tacken, E.J.; Putterill, J.; Johnston, J.W.; Schaffer, R.J. Ethylene regulates apple (Malus × domestica) fruit softening through a dose × time-dependent mechanism and through differential sensitivities and dependencies of cell wall-modifying genes. Plant Cell Physiol. 2014, 55, 1005–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, Y.; Li, B.J.; Su, G.; Zhang, M.; Grierson, D.; Chen, K.S. Transcriptional regulation of fleshy fruit texture. J. Integr. Plant Biol. 2022, 64, 1649–1672. [Google Scholar] [CrossRef]
- Ross, G.; Wegrzyn, T.; Macrae, E.; Redgwell, R. Apple beta-galactosidase activity against cell wall polysaccharides and characterization of a related cDNA clone. Plant Physiol. 1994, 106, 521–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goulao, L.F.; Cosgrove, D.J.; Oliveira, C.M. Cloning, Characterisation and Expression analyses of cDNA clones encoding cell wall-modifying enzymes isolated from ripe apples. Postharvest Biol. Technol. 2008, 48, 37–51. [Google Scholar] [CrossRef]
- Nobile, P.M.E.; Wattebled, F.; Quecini, V.; Girardi, C.L.; Lormeau, M.; Laurens, F. Identification of a novel α-L-arabinofuranosidase gene associated with mealiness in apple. J. Exp. Bot. 2011, 62, 4309–4321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, F.; Peace, C.P.; Stella, S.; Serra, S.; Musacchi, S.; Bazzani, M.; Sansavini, S.; Van de Weg, W.E. QTL dynamics for fruit firmness and softening around an ethylene-dependent polygalacturonase gene in apple (Malus × domestica Borkh.). J. Exp. Bot. 2010, 61, 3029–3039. [Google Scholar] [CrossRef] [Green Version]
- Atkinson, R.G.; Sutherland, P.W.; Johnston, S.L.; Gunaseelan, K.; Hallett, I.C.; Mitra, D.; Brummell, D.A.; Schröder, R.; Johnston, J.W.; Schaffer, R.J. Down-regulation of polygalacturonase1 alters firmness, tensile strength and water loss in apple (Malus × domestica) fruit. BMC Plant Biol. 2012, 12, 11–21. [Google Scholar] [CrossRef] [Green Version]
- Wu, B.; Shen, F.; Chen, C.J.; Liu, L.; Wang, X.; Zheng, W.Y.; Deng, Y.; Wang, T.; Huang, Z.Y.; Xiao, C.; et al. Natural variations in a pectin acetylesterase gene, MdPAE10, contribute to prolonged apple fruit shelf life. Plant Genome 2021, 14. [Google Scholar] [CrossRef]
- Chang, H.; Tong, C.B.S. The use of a combination of instrumental methods to assess change in sensory crispness during storage of a “Honeycrisp” apple breeding family. J. Texture Stud. 2018, 228–239. [Google Scholar] [CrossRef]
- Hou, J.; Sun, Y.; Chen, F.; Yu, L.; Mao, Q.; Wang, L.; Guo, X.; Liu, C. Analysis of microstructures and macrotextures for different apple cultivars based on parenchyma morphology. Microsc. Res. Tech. 2016, 79, 304–312. [Google Scholar] [CrossRef]
- He, Y.; Li, J.; Ban, Q.; Han, S.; Rao, J. Role of brassinosteroids in persimmon (Diospyros kaki L.) fruit ripening. J. Agric. Food Chem. 2018, 66, 2637–2644. [Google Scholar] [CrossRef] [PubMed]
- Renard, C.M.G.C.; Ginies, C. Comparison of the cell wall composition for flesh and skin from five different plums. Food Chem. 2009, 114, 1042–1049. [Google Scholar] [CrossRef]
- Englyst, H.; Wiggins, H.S.; Cummings, J.H. Determination of the non-starch polysaccharides in plant foods by gas-liquid chromatography of constituent sugars as alditol acetates. Analyst 1982, 107, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Busatto, N.; Tadiello, A.; Moretto, M.; Farneti, B.; Populin, F.; Vrhovsek, U.; Commisso, M.; Sartori, E.; Sonego, P.; Biasioli, F.; et al. Ethylene-auxin crosstalk regulates postharvest fruit ripening process in apple. Fruit Res. 2021, 1, 13. [Google Scholar] [CrossRef]
- Liu, X.; Renard, C.M.G.C.; Rolland-Sabaté, A.; Bureau, S.; Le Bourvellec, C. Modification of apple, beet and kiwifruit cell walls by boiling in acid conditions: Common and specific responses. Food Hydrocoll. 2021, 112. [Google Scholar] [CrossRef]
- Han, S.; Nan, Y.; Qu, W.; He, Y.; Ban, Q.; Lv, Y.; Rao, J. Exogenous γ-aminobutyric acid treatment that contributes to regulation of malate metabolism and ethylene synthesis in apple fruit during storage. J. Agric. Food Chem. 2018, 66, 13473–13482. [Google Scholar] [CrossRef]
- Li, T.; Xu, Y.; Zhang, L.; Ji, Y.; Tan, D.; Yuan, H.; Wang, A. The jasmonate-activated transcription factor mdmyc2 regulates ethylene response factor and ethylene bio-synthetic genes to promote ethylene biosynthesis during apple fruit ripening. Plant Cell. 2017, 29, 1316–1334. [Google Scholar] [CrossRef] [Green Version]
- Barreiro Elorza, P.; Ortiz, C.; Ruiz-Altisent, M.; De Smedt, V.; Schotte, S.; Andani, Z.; Wakeling, I.; Beyts, P.K. Comparison between sensory and instrumental measurements for mealiness assessment in apples. A collaborative test. J. Texture Stud. 1998, 29, 509–525. [Google Scholar] [CrossRef] [Green Version]
- Chauvin, M.A.; Ross, C.F.; Pitts, M.; Kupferman, E.; Swanson, B. Relationship between instrumental and sensory determination of apple and pear texture. J. Food Qual. 2010, 33, 181–198. [Google Scholar] [CrossRef]
- Wang, D.; Yeats, T.H.; Uluisik, S.; Rose, J.K.C.; Seymour, G.B. Fruit softening: Revisiting the role of pectin. Trends Plant Sci. 2018, 23, 302–310. [Google Scholar] [CrossRef]
- Ng, J.K.T.; Schröder, R.; Brummell, D.A.; Sutherland, P.W.; Hallett, I.C.; Smith, B.G.; Melton, L.D.; Johnston, J.W. Lower cell wall pectin solubilisation and galactose loss during early fruit development in apple (Malus × domestica) cultivar “Scifresh” are associated with slower softening rate. J. Plant Physiol. 2015, 176, 129–137. [Google Scholar] [CrossRef]
- Schäfer, J.; Bunzel, M. Maturation-related modifications of cell wall structures of kohlrabi (Brassica oleracea var. Gongylodes). Eur. Food Res. Technol. 2018, 244, 893–902. [Google Scholar] [CrossRef]
- Yang, H.; Liu, J.; Dang, M.; Zhang, B.; Li, H.; Meng, R.; Qu, D.; Yang, Y.; Zhao, Z. Analysis of β-galactosidase during fruit development and ripening in two different texture types of apple cultivars. Front. Plant Sci. 2018, 9, 539. [Google Scholar] [CrossRef]
- Harb, J.; Gapper, N.E.; Giovannoni, J.J.; Watkins, C.B. Molecular analysis of softening and ethylene synthesis and signaling pathways in a non-softening apple cultivar, “Honeycrisp” and a rapidlysoftening cultivar, “McIntosh”. Postharvest Biol. Technol. 2012, 64, 94–103. [Google Scholar] [CrossRef]
- Win, N.M.; Yoo, J.; Naing, A.H.; Kwon, J.G.; Kang, I.K. 1-Methylcyclopropene (1-MCP) treatment delays modification of cell wall pectin and fruit softening in “Hwangok” and “Picnic” apples during cold storage. Postharvest Biol. Technol. 2021, 180, 111599. [Google Scholar] [CrossRef]
- Sutherland, P.W.; Fullerton, C.G.; Schröder, R.; Hallett, I.C. Cell wall changes in Actinidia arguta during softening. Sci. Hortic. 2017, 226, 173–183. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivars | Crispness | Hardness | Mealiness |
|---|---|---|---|
| Jiguan | 2.3 ± 0.45 c | 3.4 ± 0.55 b | 8.5 ± 0.61 a |
| Yindu | 2.1 ± 0.74 c | 3.4 ± 1.14 b | 8.9 ± 0.22 a |
| Qinguan | 5 ± 1.22 b | 5.4 ± 0.89 ab | 5.6 ± 0.54 b |
| Fuji | 8.2 ± 0.84 a | 4.4 ± 1.81 b | 2.9 ± 0.96 c |
| Huaguan | 8.1 ± 1.02 a | 4.2 ± 2.16 b | 2.7 ± 0.44 c |
| Cripps pink | 7.4 ± 1.39 a | 6.4 ± 1.14 a | 3 ± 0.79 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Lin, S.; Zhang, M.; Lv, Y.; Ma, Y.; Rao, J.; Zhu, Q. Changes in Cell Wall Sugar Neutral Composition Contribute to Apple Texture Loss during Storage among Cultivars. Horticulturae 2023, 9, 292. https://doi.org/10.3390/horticulturae9030292
Liu H, Lin S, Zhang M, Lv Y, Ma Y, Rao J, Zhu Q. Changes in Cell Wall Sugar Neutral Composition Contribute to Apple Texture Loss during Storage among Cultivars. Horticulturae. 2023; 9(3):292. https://doi.org/10.3390/horticulturae9030292
Chicago/Turabian StyleLiu, Hui, Shiyu Lin, Mengyuan Zhang, Yanrong Lv, Yanping Ma, Jingping Rao, and Qinggang Zhu. 2023. "Changes in Cell Wall Sugar Neutral Composition Contribute to Apple Texture Loss during Storage among Cultivars" Horticulturae 9, no. 3: 292. https://doi.org/10.3390/horticulturae9030292