Brcd1 Is Associated with Plant Height through the Gibberellin Pathway in Brassica rapa L.

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants and Measurement of Their Traits
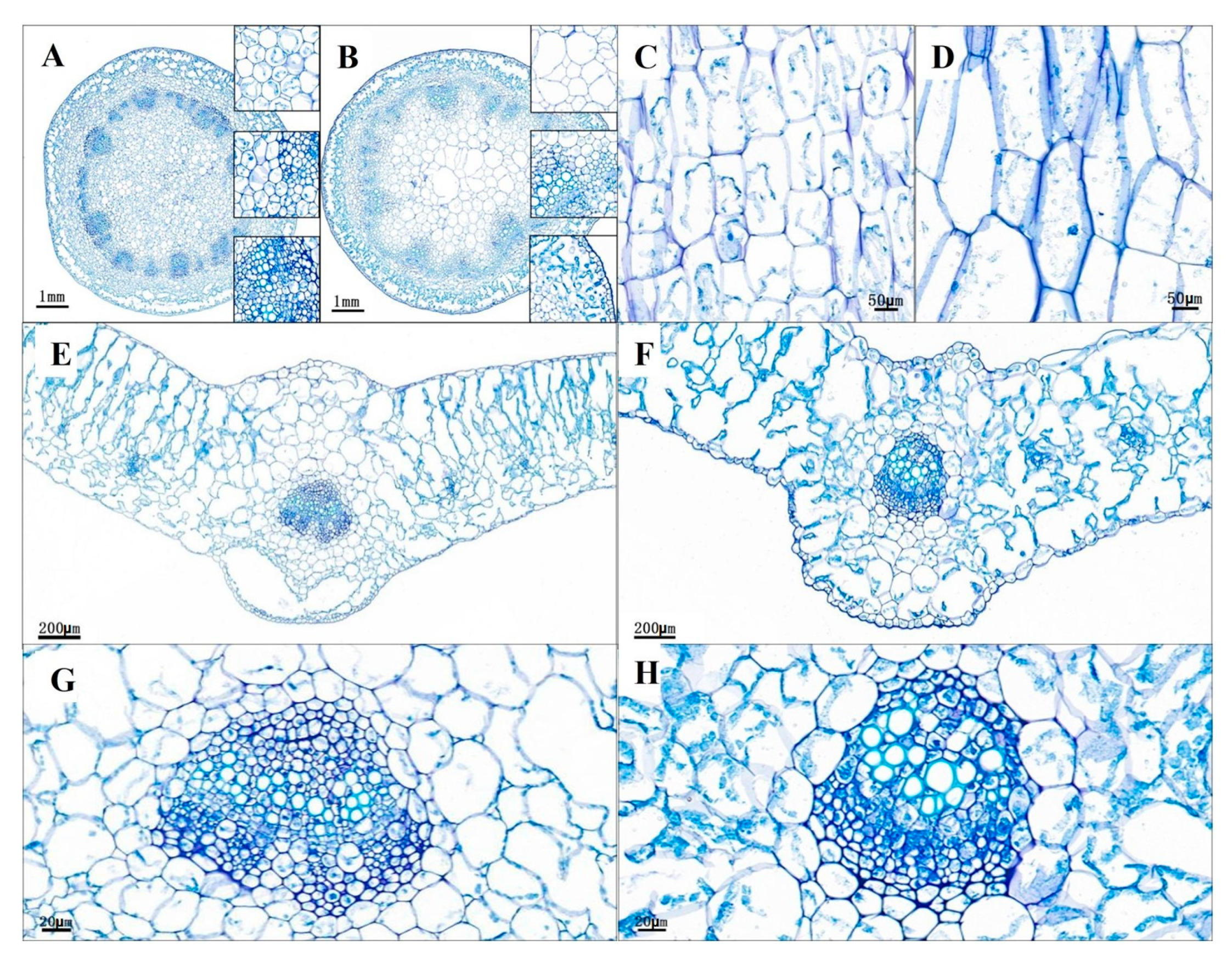
2.2. Stem and Leaf Cell Morphological Characterization
2.3. Photosynthetic Index Determination
2.4. Chlorophyll Content Determination
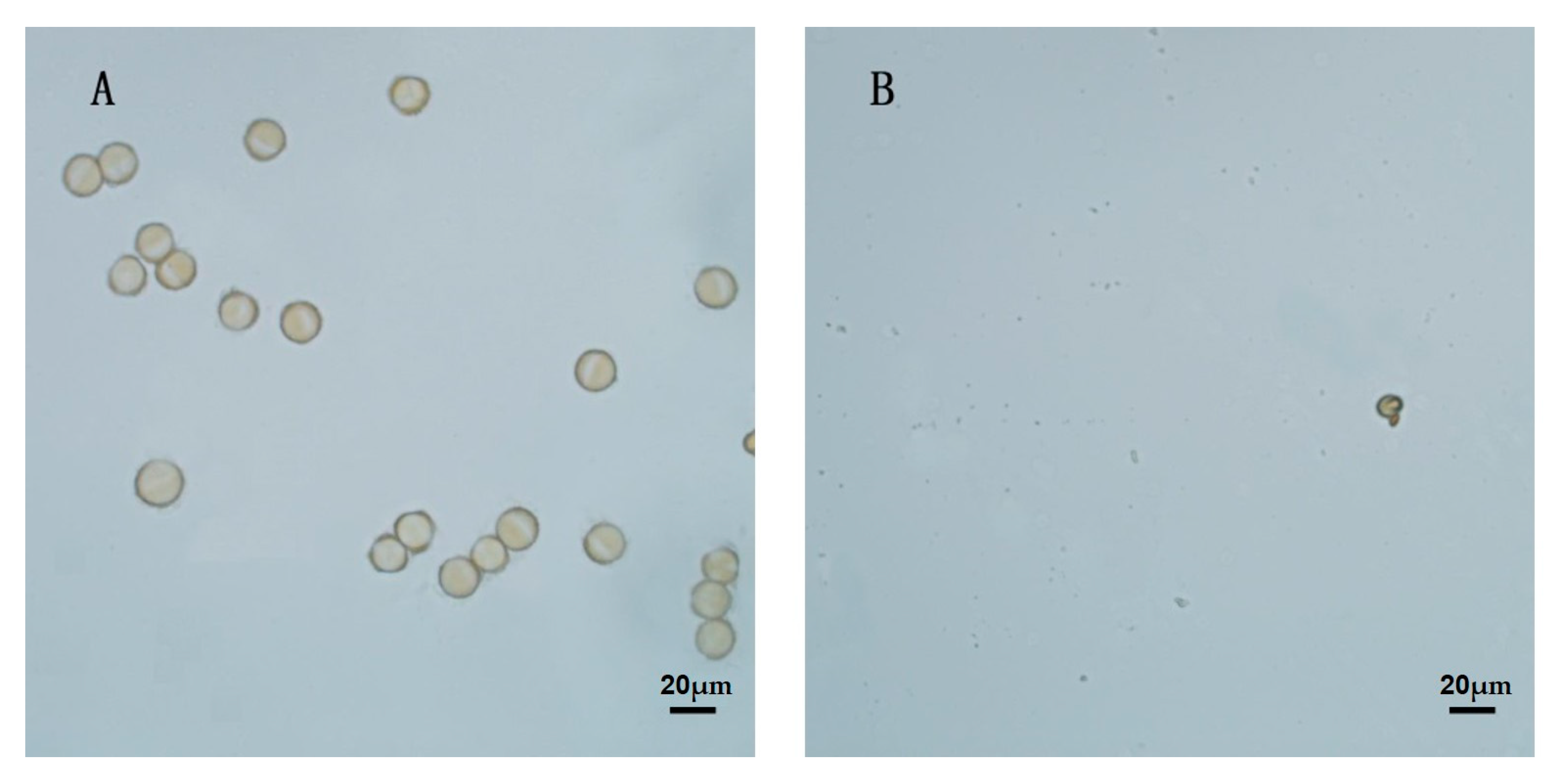
2.5. Pollen Viability Determination
2.6. RNA Isolation and Extreme Pool Construction
2.7. BSR-seq
2.8. Extraction of DNA and the Development of Markers
2.9. Preliminary Localization of Brcd1 Dwarfing Mutation
2.10. Exogenous GA3 Treatment and Candidate Gene Annotation Analysis
2.11. Candidate Gene Expression Determination
2.12. Sequence Analysis of Candidate Genes
3. Results
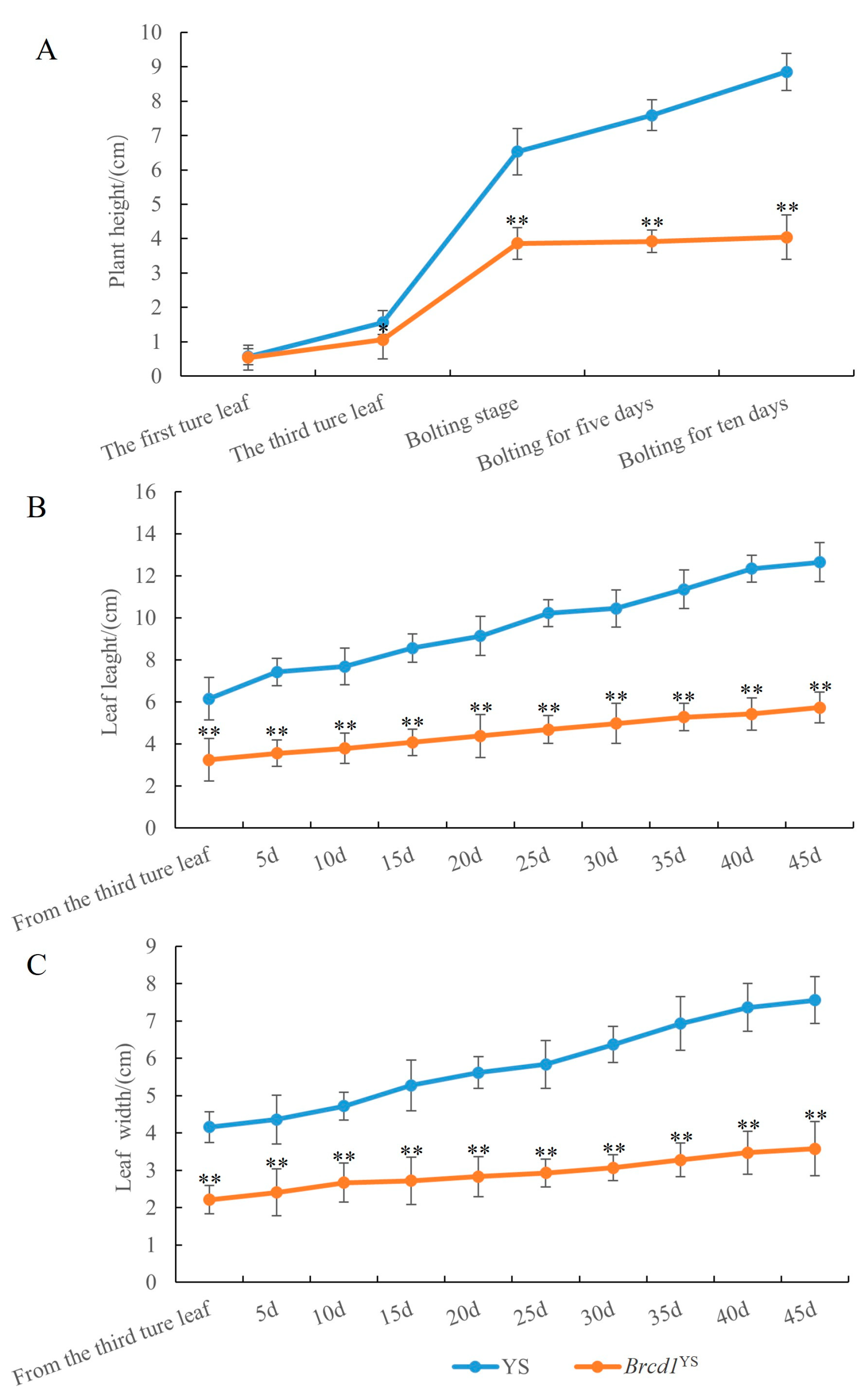
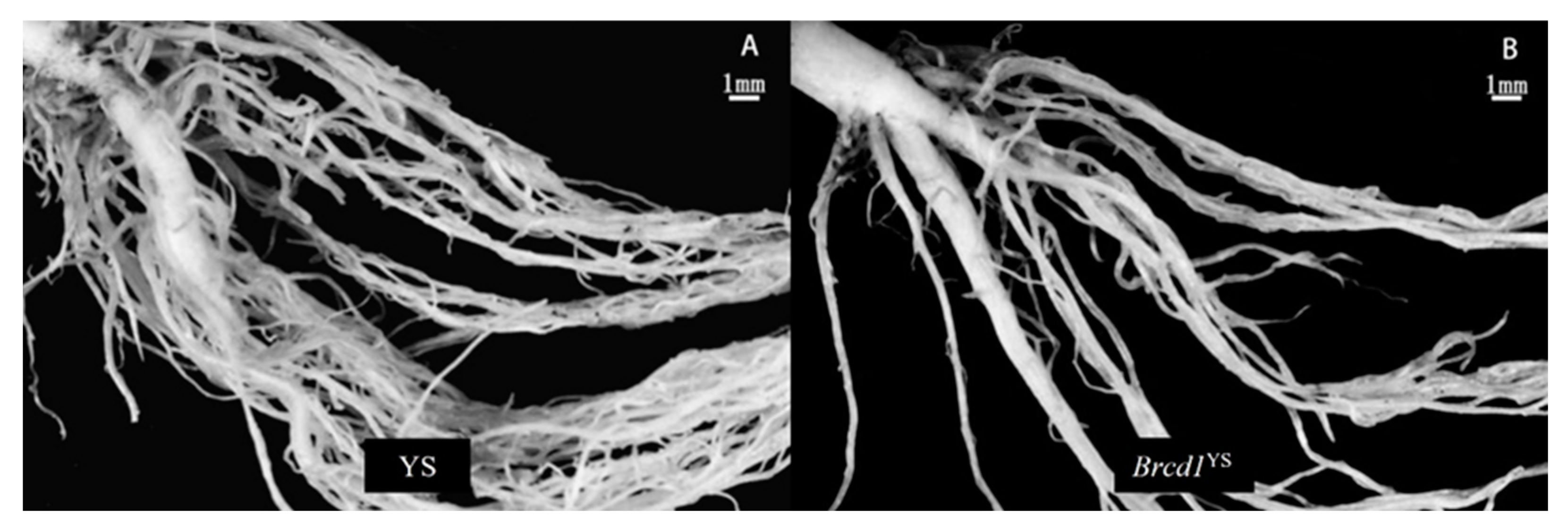
3.1. Phenotypic Characterization of the Dwarf Mutant
3.2. Analysis of Physiological Indices
3.3. Identification Stem and Leaf Cell Morphologies of the Brcd1YS Mutant
3.4. Primary Mapping and Validation of Brcd1 Locus
3.5. Brcd1 Mutant Plants Could Be Rescued by Exogenous GA3 Treatment
3.6. Prediction of Candidate Genes
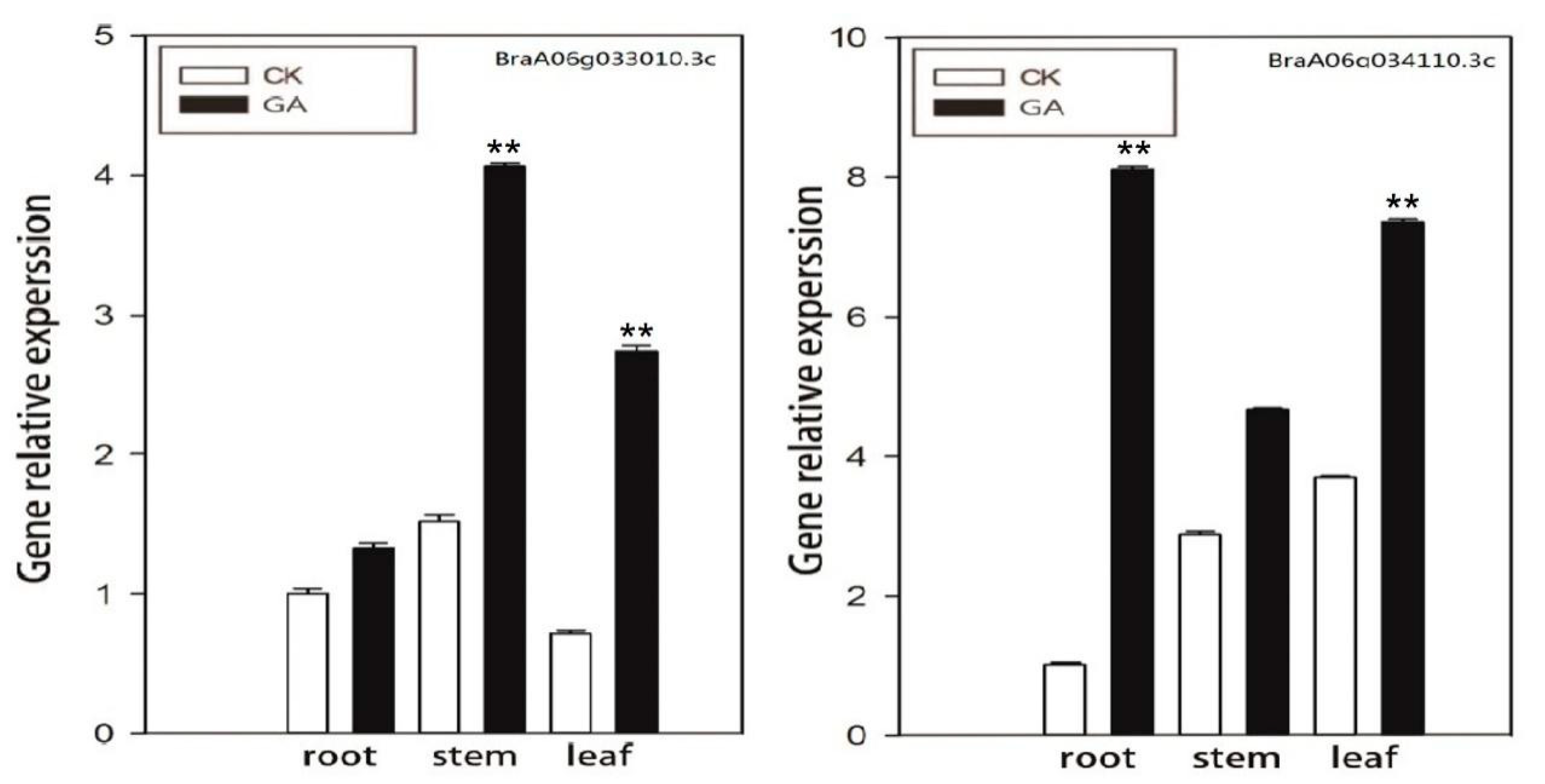
3.7. Expression Analysis of GA-Related Genes
3.8. Sequence Variation Analysis of Candidate Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Ethical Statement
References
- Carpio, D.P.; Basnet, P.K.; De Vos, R.C.H.; Maliepaard, C.; Visser, C.; Bonnema, G. The patterns of population differentiation in a Brassica rapa core collection. Theor. Appl. Genet. 2011, 122, 1105–1118. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.; Zhang, L.; Tang, M.; Cai, Y.; Liu, J.; Liu, H.; Liu, J.; Terzaghi, W.; Wang, H.; Hua, W.; et al. CRISPR/Cas9-targeted mutagenesis of the BnaA03.BP gene confers semi-dwarf and compact architecture to rapeseed (Brassica napus L.). Plant Biotechnol. J. 2021, 19, 2383–2385. [Google Scholar] [CrossRef] [PubMed]
- Muangprom, A.; Thomas, S.G.; Sun, T.P.; Osborn, T.C. A novel dwarfing mutation in a green revolution gene from Brassica rapa. Plant Physio. 2005, 137, 931–938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Zhao, J.; Lu, W.; Deng, D. Gibberellin in plant height control: Old player, new story. Plant Cell. Rep. 2017, 36, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Castorina, G.; Consonni, G. The role of Brassinosteroids in controlling plant height in Poaceae: A genetic perspective. Int. J. Mol. Sci. 2020, 11, 1191. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, S. Gibberellin metabolism and its regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef]
- Davière, J.M.; Achard, P. Gibberellin signaling in plants. Development 2013, 140, 1147–1151. [Google Scholar] [CrossRef] [Green Version]
- Hartweck, L.M. Gibberellin signaling. Planta 2008, 229, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Hedden, P. The genes of the Green Revolution. Trends Genet. 2003, 19, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Tang, T.; Botwright Acuña, T.; Spielmeyer, W.; Richards, R.A. Effect of gibberellin-sensitive Rht18 and gibberellin-insensitive Rht-D1b dwarfing genes on vegetative and reproductive growth in bread wheat. J. Exp. Bot. 2021, 2, 445–458. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Richards, D.E.; Hartley, N.M.; Murphy, G.P.; Devos, K.M.; Flintham, J.E.; Beales, J.; Fish, L.J.; Worland, A.J.; Pelica, F.; et al. ‘Green revolution’ genes encode mutant gibberellin response modulators. Nature 1999, 400, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Tomlinson, L.; Yang, Y.; Emenecker, R.; Smoker, M.; Taylor, J.; Perkins, S.; Smith, J.; MacLean, D.; Olszewski, N.E.; Jones, J.D.G. Using CRISPR/Cas9 genome editing in tomato to create a gibberellin-responsive dominant dwarf DELLA allele. Plant Biotechnol. J. 2019, 17, 132–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, M.; Zhang, L.; Tang, M.; Liu, J.; Liu, H.; Yang, H.; Fan, S.; Terzaghi, W.; Wang, H.; Hua, W. Knockout of two BnaMAX1 homologs by CRISPR/Cas9-targeted mutagenesis improves plant architecture and increases yield in rapeseed (Brassica napus L.). Plant Biotechnol. J. 2020, 18, 644–654. [Google Scholar] [CrossRef] [Green Version]
- Zhao, B.; Li, H.; Li, J.; Wang, B.; Dai, C.; Wang, J.; Liu, K. Brassica napus DS-3, encoding a DELLA protein, negatively regulates stem elongation through gibberellin signaling pathway. Theor. Appl. Genet. 2017, 130, 727–741. [Google Scholar] [CrossRef] [PubMed]
- Muangprom, A.; Osborn, T.C. Characterization of a dwarf gene in Brassica rapa, including the identification of a candidate gene. Theor. Appl. Genet. 2004, 108, 1378–1384. [Google Scholar] [CrossRef]
- Yan, J.; Liao, X.; He, R.; Zhong, M.; Feng, P.; Li, X.; Tang, D.; Liu, X.; Zhao, X. Ectopic expression of GA 2-oxidase 6 from rapeseed (Brassica napus L.) causes dwarfism, late flowering and enhanced chlorophyll accumulation in Arabidopsis thaliana. Plant Physiol. Biochem. 2017, 111, 10–19. [Google Scholar] [CrossRef]
- Zhao, B.; Wang, B.; Li, Z.; Guo, T.; Zhao, J.; Guan, Z.; Liu, K. Identification and characterization of a new dwarf locus DS-4 encoding an Aux/IAA7 protein in Brassica napus. Theor. Appl. Genet. 2019, 132, 1435–1449. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Zhang, L.; Zhu, R.; Jiang, X.; Yue, C.; Su, Y.; Ren, H.; Wang, M. A Gain-of-Function Mutant of IAA7 Inhibits Stem Elongation by Transcriptional Repression of EXPA5 Genes in Brassica napus. Int. J. Mol. Sci. 2021, 22, 9018. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, S.; Liu, Z.; Tang, X.; Feng, H. Changes in endogenous phytohormones regulated by microRNA-target mRNAs contribute to the development of Dwarf Autotetraploid Chinese Cabbage (Brassica rapa L. ssp. pekinensis). Mol. Genet. Genom. 2018, 293, 1535–1546. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, X.; Ji, X.; Zhang, L.; Liu, Y.; Lv, X.; Feng, H. Identification and validation of a major QTL controlling the presence/absence of leaf lobes in Brassica rapa L. Euphytica 2015, 205, 761–771. [Google Scholar] [CrossRef]
- Raj, J.D.; Nelson, J.A.; Rao, K.S. A study on the effects of some reinforcers to improve performance of employees in a retail industry. Behav. Modif. 2006, 30, 848–866. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Huang, S.; Liu, Z.; Hou, L.; Feng, H. Mutation in EMB1923 gene promoter is associated with chlorophyll deficiency in Chinese cabbage (Brassica campestris ssp. pekinensis). Physiol. Plant. 2019, 166, 909–920. [Google Scholar] [CrossRef]
- Arnon, D.I. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiol. 1949, 24, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, C.; Liu, Z.; Huang, S.; Feng, H. Mapping of the male sterile mutant gene ftms in Brassica rapa L. ssp. pekinensis via BSR-Seq combined with whole-genome resequencing. Theor. Appl. Genet. 2018, 132, 355–370. [Google Scholar] [CrossRef]
- Qu, G.; Gao, Y.; Wang, X.; Fu, W.; Sun, Y.; Gao, X.; Wang, W.; Hao, C.; Feng, H.; Wang, Y. Fine mapping and analysis of candidate genes for qFT7.1, a major quantitative trait locus controlling flowering time in Brassica rapa L. Theor. Appl. Genet. 2022, 135, 2233–2246. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.; Cai, X.; Wu, J.; Liu, M.; Grob, S.; Cheng, F.; Liang, J.; Cai, C.; Liu, Z.; Liu, B.; et al. Improved Brassica rapa reference genome by single-molecule sequencing and chromo-some conformation capture technologies. Hortic Res. 2018, 5, 50. [Google Scholar] [CrossRef] [Green Version]
- Nakajima, M.; Shimada, A.; Takashi, Y.; Kim, Y.-C.; Park, S.-H.; Ueguchi-Tanaka, M.; Suzuki, H.; Katoh, E.; Iuchi, S.; Kobayashi, M.; et al. Identification and characterization of Arabidopsis gibberellin receptors. Plant J. 2006, 46, 880–889. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [Green Version]
- Su, A.; Song, W.; Xing, J.; Zhao, Y.; Zhang, R.; Li, C.; Duan, M.; Luo, M.; Shi, Z.; Zhao, J. Identification of Genes Potentially Associated with the Fertility Instability of S-Type Cytoplasmic Male Sterility in Maize via Bulked Segregant RNA-Seq. PLOS ONE 2016, 11, e0163489. [Google Scholar] [CrossRef] [Green Version]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucl. Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, Y.; Huang, S.; Qu, G.; Fu, W.; Zhang, M.; Liu, Z.; Feng, H. The mutation of ent-kaurene synthase, a key enzyme involved in gibberellin biosynthesis, confers a non-heading phenotype to Chinese cabbage (Brassica rapa L. ssp pekinensis). Hortic. Res. 2020, 1, 178. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402-8. [Google Scholar] [CrossRef]
- Livak Narasimhan, V.; Danecek, P.; Scally, A.; Xue, Y.; Tyler-Smith, C.; Durbin, R. BCFtools/RoH: A hidden Markov model approach for detecting autozygosity from next-generation sequencing data. Bioinformatics 2016, 32, 1749–1751. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.N.; Liu, Z.Y.; Yao, R.P.; Li, D.Y.; Feng, H. Comparative transcriptome analysis of the petal degeneration mutant pdm in Chinese cabbage (Brassica campestris ssp pekinensis) using RNA-seq. Mol. Genet. Genom. 2015, 290, 1833–1847. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Lu, Y.; Yang, H.; He, J.; Hu, Z.; Hu, X.; Luan, M.; Zhang, L.; Fan, Y.; Wang, L. ZmGRF, a GA regulatory factor from maize, promotes flowering and plant growth in Arabidopsis. Plant Mol. Biol. 2015, 87, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M.; Zeng, B.; Tang, D.; Yang, J.; Qu, L.; Yan, J.; Wang, X.; Li, X.; Liu, X.; Zhao, X. The blue light receptor CRY1 interacts with GID1 and DELLA proteins to repress GA signaling during photomorphogenesis in Arabidopsis. Mol. Plant 2021, 2, 1328–1342. [Google Scholar] [CrossRef]
- Fu, W.; Huang, S.; Gao, Y.; Zhang, M.; Qu, G.; Wang, N.; Liu, Z.; Feng, H. Role of BrSDG8 on bolting in Chinese cabbage (Brassica rapa). Appl Genet. 2020, 133, 2937–2948. [Google Scholar] [CrossRef]
- Wang, S.; Wang, Y. Harnessing hormone gibberellin knowledge for plant height regulation. Plant Cell. Rep. 2022, 41, 1945–1953. [Google Scholar] [CrossRef]
- Guizhen, L.; Dongbing, L.; Shaoshan, L.; Hongpeng, H. STO and GA negatively regulate UV-B-induced Arabidopsis root growth inhibition. Plant Signal. Behav. 2019, 14, 1675471. [Google Scholar] [CrossRef]
- Griffiths, J.; Murase, K.; Rieu, I.; Zentella, R.; Zhang, Z.L.; Powers, S.J.; Gong, F.; Phillips, A.L.; Hedden, P.; Sun, T.P.; et al. Genetic characterization and functional analysis of the GID1 gibberellin receptors in Arabidopsis. Plant Cell. 2006, 18, 3399–3414. [Google Scholar] [CrossRef] [Green Version]
- Uchi, S.; Suzuki, H.; Kim, Y.C.; Iuchi, A.; Kuromori, T.; Ueguchi-Tanaka, M.; Asami, T.; Yamaguchi, I.; Matsuoka, M.; Kobayashi, M.; et al. Multiple loss-of-function of Arabidopsis gibberellin receptor AtGID1s completely shuts down a gibberellin signal. Plant J. 2007, 50, 958–966. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z.; Liu, X.; Zhang, B.; Yuan, X.; Xing, Y.; Liu, H.; Luo, L.; Chen, G.; Xiong, L. Genetic analyses of lodging resistance and yield provide insights into post-Green-Revolution breeding in rice. Plant Biotechnol. J. 2021, 19, 814–829. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Yang, L.; Tang, M.; Liu, L.; Huang, J.; Tong, C.; Xiang, Y.; Liu, S.; Cheng, X.; Xie, M. Genome-wide association study reveals a GLYCOGEN SYNTHASE KINASE 3 gene regulating plant height in Brassica napus. Front. Plant Sci. 2022, 2, 1061196. [Google Scholar] [CrossRef] [PubMed]
- Shearman, J.R.; Vejchasarn, P.; Naktang, C.; Phansenee, Y.; Jomchai, N.; Lanceras-Siangliw, J.; Tangphatsornruang, S.; Toojinda, T. Rice height QTLs in KDML105 chromosome segment substitution lines. Genomics 2022, 114, 482–487. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.S.; Park, S.; Kang, K.; Lee, K.; Back, K. Tyramine accumulation in rice cells caused a dwarf phenotype via reduced cell division. Planta 2011, 233, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Guo, M.; Yang, F.; Duncan, K.; Jackson, D.; Rafalski, A.; Wang, S.; Li, B. Mutations in an AP2 transcription factor-like gene affect internode length and leaf shape in maize. PLoS ONE 2012, 7, e37040. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Photosynthetic Rate (μmol m−2s−1) | Intercellular CO2 Concentration (μmol mol−1) | Transpiration Rate (mmol m−2s−1) | Stomatal Conductance (mmol m−2s−1) | Chlorophyll a (mg g−1 FW) | Chlorophyll b (mg g−1 FW) | Total Chlorophyll (mg g−1 FW) | Carotenoid (mg g−1 FW) | |
|---|---|---|---|---|---|---|---|---|
| YS | 20.44 ± 1.49 | 404 ± 4.05 | 6.17 ± 0.30 | 6.17 ± 16.15 | 1.61 ± 0.01 | 0.47 ± 0.03 | 2.08 ± 0.04 | 0.52 ± 0.03 |
| Brcd1YS | 25.91 ± 0.74 * | 390 ± 3.31 * | 5.04 ± 0.28 * | 7.69 ± 25.0 * | 1.67 ± 0.01 * | 0.55 ± 0.03 * | 2.20 ± 0.02 * | 0.46 ± 0.02 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Hu, J.; Fang, B.; Gao, X.; Hao, C.; Mu, Y.; Feng, H.; Qu, G.; Wang, Y. Brcd1 Is Associated with Plant Height through the Gibberellin Pathway in Brassica rapa L. Horticulturae 2023, 9, 282. https://doi.org/10.3390/horticulturae9020282
Wang W, Hu J, Fang B, Gao X, Hao C, Mu Y, Feng H, Qu G, Wang Y. Brcd1 Is Associated with Plant Height through the Gibberellin Pathway in Brassica rapa L. Horticulturae. 2023; 9(2):282. https://doi.org/10.3390/horticulturae9020282
Chicago/Turabian StyleWang, Wei, Junyu Hu, Bing Fang, Xu Gao, Chunming Hao, Yizhuo Mu, Hui Feng, Gaoyang Qu, and Yugang Wang. 2023. "Brcd1 Is Associated with Plant Height through the Gibberellin Pathway in Brassica rapa L." Horticulturae 9, no. 2: 282. https://doi.org/10.3390/horticulturae9020282