

Thidiazuron Induced In Vitro Plant Regeneration, Phenolic Contents, Antioxidant Potential, GC-MS Profiles and Nuclear Genome Stability of Plectranthus amboinicus (Lour.) Spreng




Abstract
:1. Introduction
2. Materials and Methods
2.1. Starting Materials and Aseptic Cultures
2.2. Thidiazuron (TDZ) Treatment and Shoot Regeneration
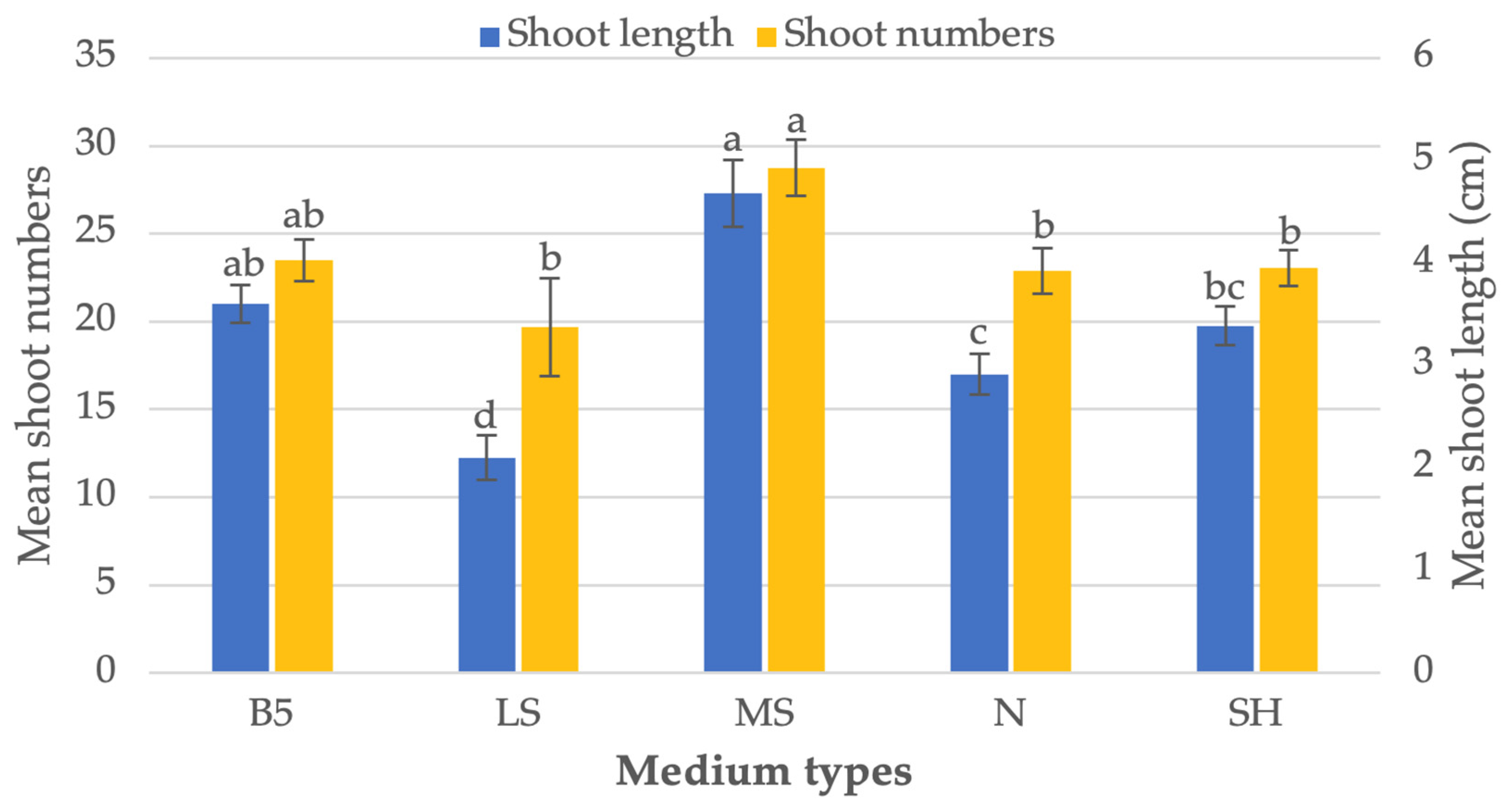
2.3. Effect of Basal Media on Shoot Proliferation
2.4. Rooting and Acclimatization
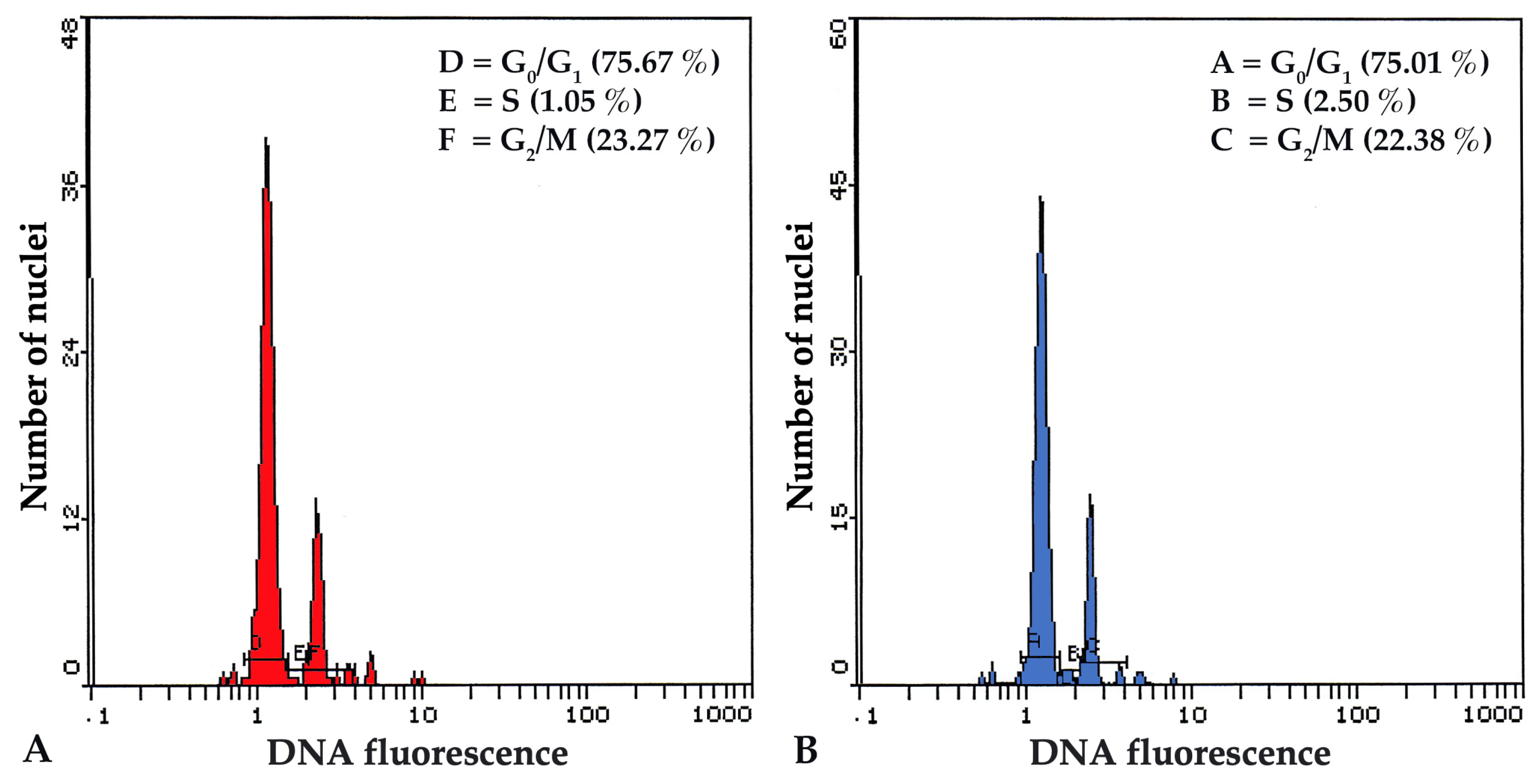
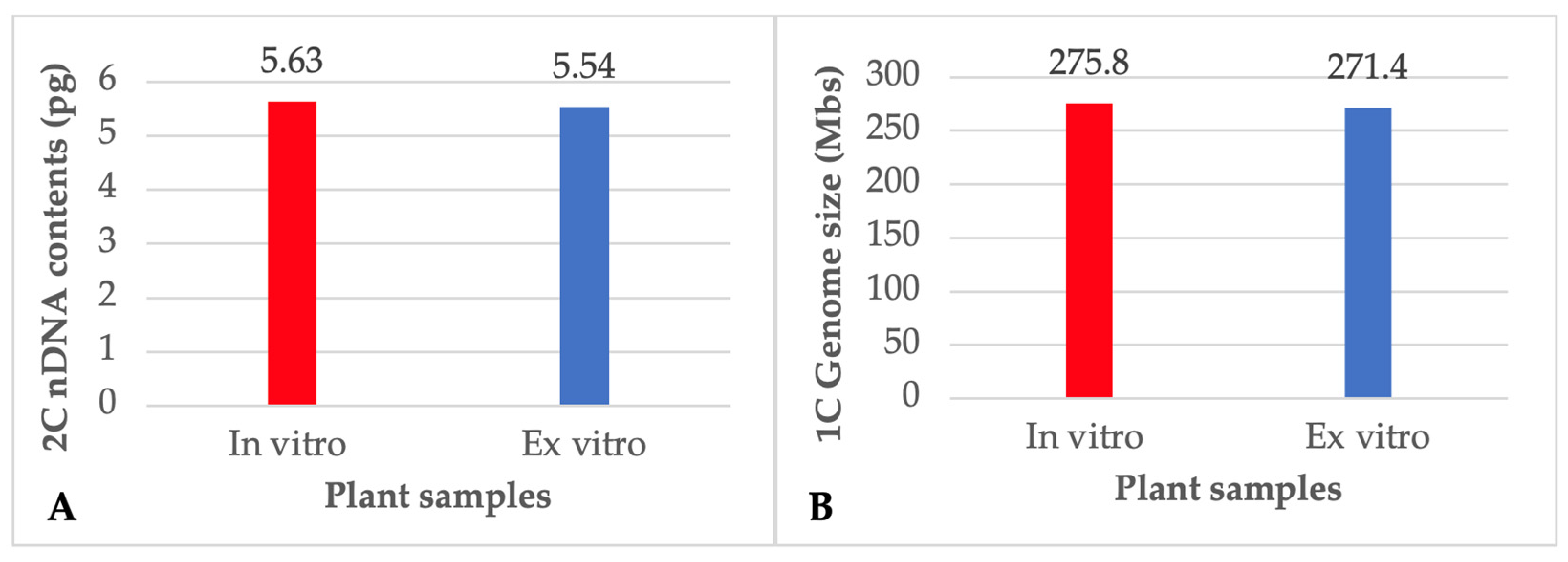
2.5. Flow Cytometry (FCM) and Nuclear DNA Content
2.6. Preparation of Plant Extracts
2.6.1. Determination of Total Phenolics
2.6.2. Determination of Total Tannins
2.6.3. Determination of Total Flavonoid
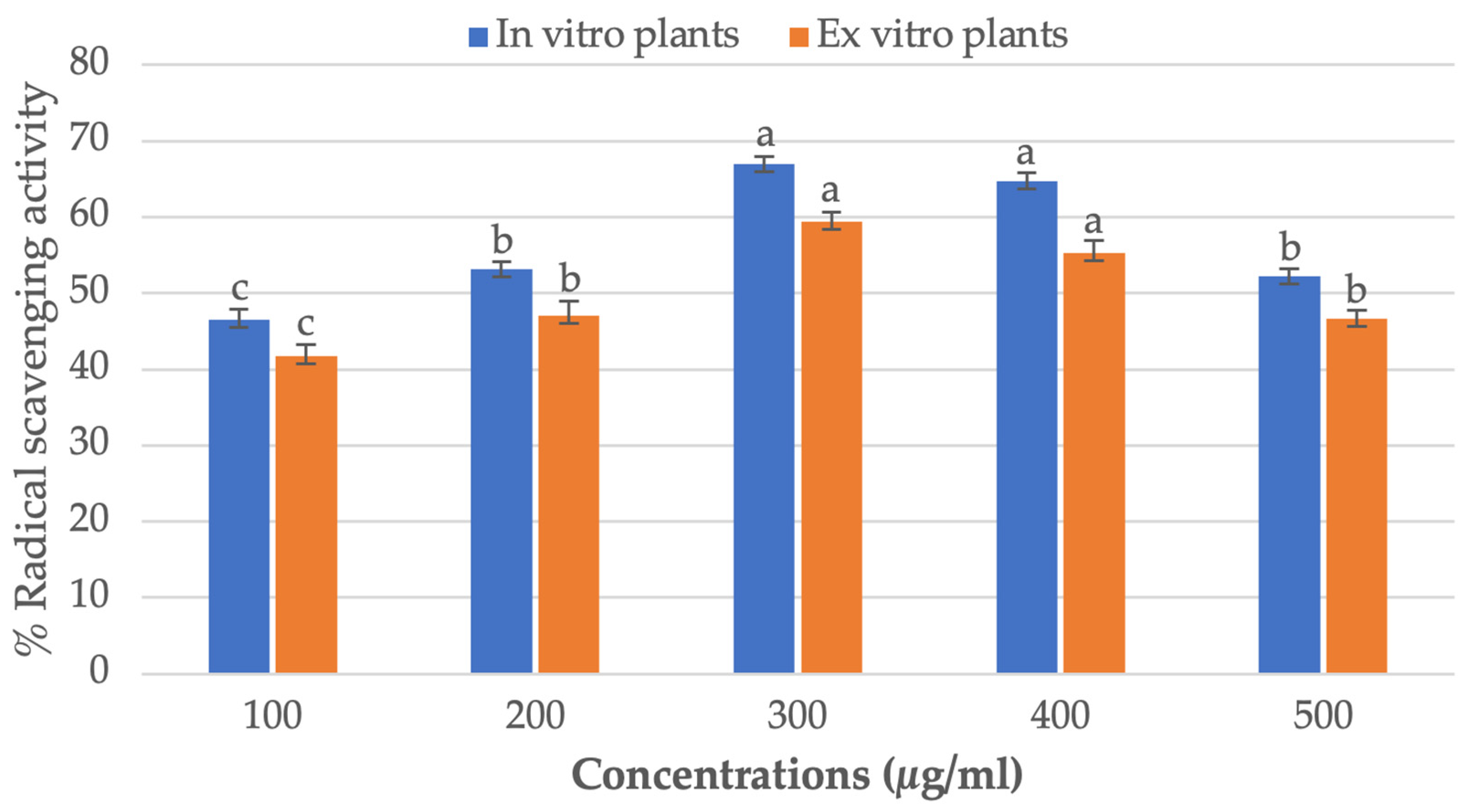
2.6.4. Antioxidant and 2,2-Diphenyl-1-picrylhydrazyl (DPPH) Assay
2.6.5. Gas Chromatography-Mass Spectrometry (GC-MS)
2.7. Statistical Analysis
3. Results and Discussion
3.1. Effect of TDZ Pre-Treatment and Shoot Regeneration
3.2. Effect of Basal Media on Shoot Regeneration
3.3. Rooting and Acclimatization
3.4. Flow Cytometry (FCM) and Nuclear DNA Content
3.5. Phytochemical Analysis
3.6. Antioxidant and DPPH Radical Scavenging Assay
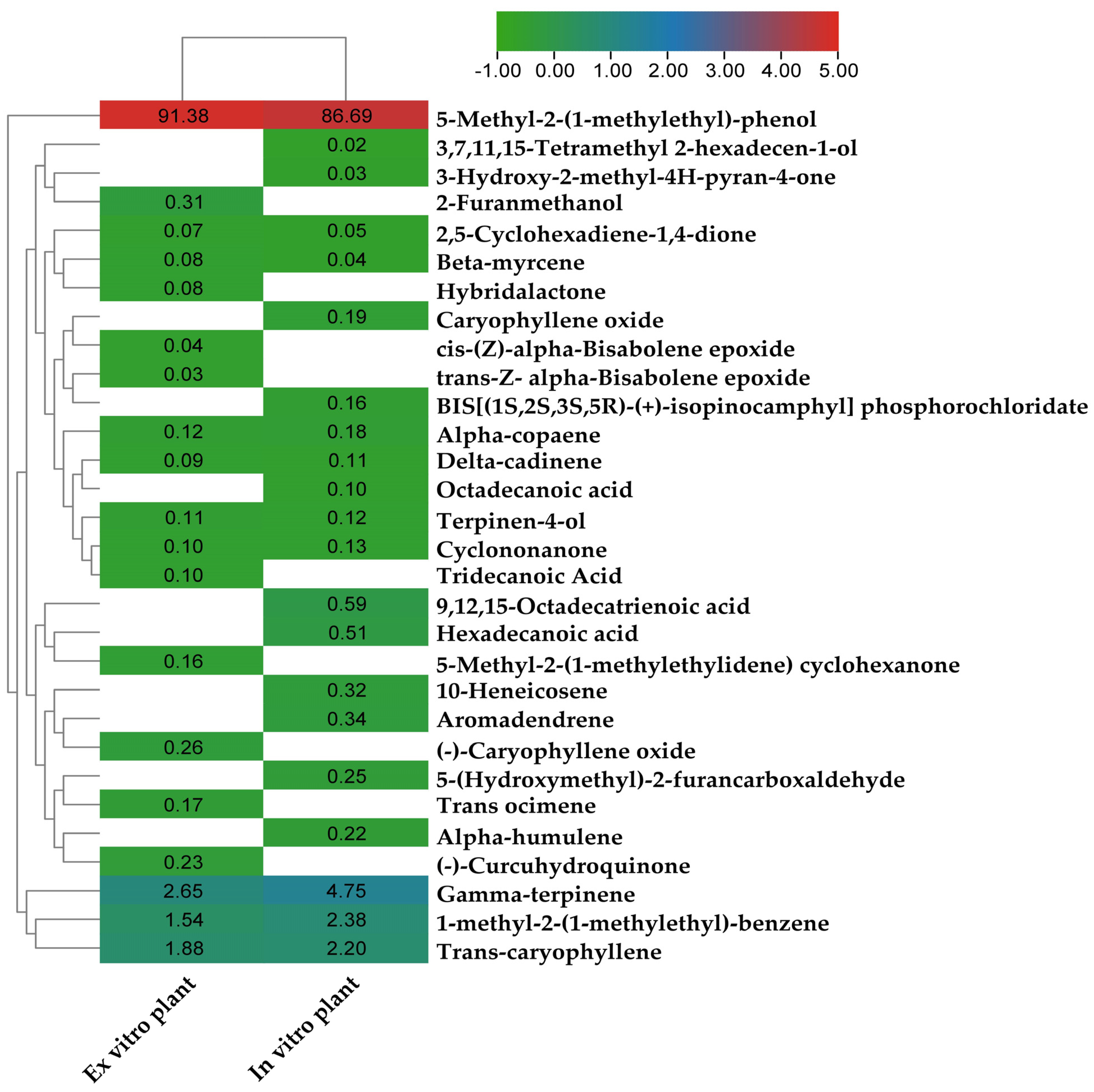
3.7. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Arumugam, G.; Swamy, M.K.; Sinniah, U.R. Plectranthus amboinicus (Lour.) Spreng: Botanical, Phytochemical, Pharmacological and Nutritional Significance. Molecules 2016, 21, 369. [Google Scholar] [CrossRef] [Green Version]
- Ashaari, N.S.; Mohamad, N.E.; Afzinizam, A.H.; Ab. Rahim, M.-H.; Lai, K.S.; Ong Abdullah, J. Chemical Composition of Hexane-Extracted Plectranthus amboinicus Leaf Essential Oil: Maximizing Contents on Harvested Plant Materials. Appl. Sci. 2021, 11, 10838. [Google Scholar] [CrossRef]
- Bhatt, P.; Joseph, G.S.; Negi, P.S.; Varadaraj, M.C. Chemical Composition and Nutraceutical Potential of Indian Borage (Plectranthus amboinicus) Stem Extract. J. Chem. 2013, 2013, 320329. [Google Scholar] [CrossRef] [Green Version]
- Vasconcelos, S.E.C.B.; Melo, H.M.; Cavalcante, T.T.A.; Júnior, F.E.A.C.; de Carvalho, M.G.; Menezes, F.G.R.; de Sousa, O.V.; Costa, R.A. Plectranthus amboinicus essential oil and carvacrol bioactive against planktonic and biofilm of oxacillin- and vancomycin-resistant Staphylococcus aureus. BMC Complement. Altern. Med. 2017, 17, 462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sindhu, M.; Poonkothai, M. Phytochemical and antimicrobial analyses of Plectranthus amboinicus leaf extracts. Res. J. Pharm. Technol. 2021, 14, 6379–6384. [Google Scholar] [CrossRef]
- Gurgel, A.P.A.D.; da Silva, J.G.; Grangeiro, A.R.S.; Oliveira, D.C.; Lima, C.M.P.; da Silva, A.C.P.; Oliveira, R.A.G.; Souza, I.A. In vivo study of the anti-inflammatory and antitumor activities of leaves from Plectranthus amboinicus (Lour.) Spreng (Lamiaceae). J. Ethnopharmacol. 2009, 125, 361–363. [Google Scholar] [CrossRef]
- Akinbo, D.B.; Onyeaghala, A.A.; Emomidue, J.O.; Ogbhemhe, S.O.; Okpoli, H.C. Phytochemical and anti-inflammatory activities of aqueous leaf extract of Indian borage (oregano) on rats induced with inflammation. Cancer Biomark. 2018, 22, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.-M.; Cheng, C.-M.; Hung, L.-M.; Chung, Y.-S.; Wu, R.-Y. Potential Use of Plectranthus amboinicus in the Treatment of Rheumatoid Arthritis. Evid.-Based Complement. Altern. Med. 2010, 7, 174726. [Google Scholar] [CrossRef] [Green Version]
- Manjamalai, A.; Grace, V.M.B. The Chemotherapeutic Effect of Essential Oil of Plectranthus amboinicus (Lour) on Lung Metastasis Developed by B16F-10 Cell Line in C57BL/6 Mice. Cancer Investig. 2013, 31, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Kumaran, A.; Joel Karunakaran, R. Antioxidant and free radical scavenging activity of an aqueous extract of Coleus aromaticus. Food Chem. 2006, 97, 109–114. [Google Scholar] [CrossRef]
- Bhatt, P.; Negi, P.S. Antioxidant and antibacterial activities in the leaf extracts of Indian borage (Plectranthus amboinicus). Food Nutr. Sci. 2012, 2012, 146–152. [Google Scholar]
- Kozłowska, M.; Ścibisz, I.; Przybył, J.; Ziarno, M.; Żbikowska, A.; Majewska, E. Phenolic Contents and Antioxidant Activity of Extracts of Selected Fresh and Dried Herbal Materials. Pol. J. Food Nutr. Sci. 2021, 71, 269–278. [Google Scholar] [CrossRef]
- Viswanathaswamy, A.; Koti, B.; Gore, A.; Thippeswamy, A.; Kulkarni, R. Antihyperglycemic and antihyperlipidemic activity of Plectranthus amboinicus on normal and alloxan-induced diabetic rats. Indian J. Pharm. Sci. 2011, 73, 139. [Google Scholar] [PubMed] [Green Version]
- Patel, R.D.; Mahobia, N.K.; Singh, M.P.; Singh, A.; Sheikh, N.W.; Alam, G.; Singh, S.K. Antioxidant potential of leaves of Plectranthus amboinicus (Lour) Spreng. Pharm. Lett. 2010, 2, 240–245. [Google Scholar]
- Varshney, A.; Anis, M. Trees: Propagation and Conservation: Biotechnological Approaches for Propagation of a Multipurpose Tree, Balanites Aegyptiaca Del; Springer Science & Business Media: New Delhi, India, 2014. [Google Scholar]
- Ahmad, N.; Javed, S.B.; Khan, M.I.; Anis, M. Rapid plant regeneration and analysis of genetic fidelity in micropropagated plants of Vitex trifolia: An important medicinal plant. Acta Physiol. Plant. 2013, 35, 2493–2500. [Google Scholar] [CrossRef]
- Ab Rahman, Z.; Noor, E.S.M.; Ali, M.S.M.; Mirad, R.; Othman, A.N. In vitro micropropagation of a valuable medicinal plant, Plectranthus amboinicus. Am. J. Plant Sci. 2015, 6, 1091–1097. [Google Scholar] [CrossRef] [Green Version]
- Arumugam, G.; Sinniah, U.R.; Swamy, M.K.; Lynch, P.T. Micropropagation and essential oil characterization of Plectranthus amboinicus (Lour.) Sprengel, an aromatic medicinal plant. In Vitro Cell. Dev. Biol.-Plant 2020, 56, 491–503. [Google Scholar] [CrossRef]
- Faisal, M.; Alatar, A.A. Establishment of an Efficient In Vitro Propagation Method for a Sustainable Supply of Plectranthus amboinicus (Lour.) and Genetic Homogeneity Using Flow Cytometry and SPAR Markers. Horticulturae 2022, 8, 693. [Google Scholar] [CrossRef]
- Sreedevi, E.; Anuradha, M.; Pullaiah, T. Plant regeneration from leaf-derived callus in Plectranthus barbatus Andr. [Syn.: Coleus forskohlii (Wild.) Briq.]. Afr. J. Biotechnol. 2013, 12, 2441–2448. [Google Scholar]
- Tsegaw, M.; Feyissa, T. Micropropagation of Plectranthus edulis (Vatke) Agnew from meristem cultur. Afr. J. Biotechnol. 2014, 13, 3682–3688. [Google Scholar] [CrossRef] [Green Version]
- Kebede, B.; Abera, B. Micropropagation of Plectranthus edulis (Vatke) Agnew from shoot tip and nodal explants. Afr. J. Agric. Res. 2015, 10, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Thaniarasu, R.; Kumar, T.S.; Rao, M. In vitro Propagation of Plectranthus bourneae Gamble-An Endemic Red Listed Plant. Plant Tissue Cult. Biotechnol. 2015, 25, 273–284. [Google Scholar] [CrossRef] [Green Version]
- Thaniarasu, R.; Senthil Kumar, T.; Rao, M.V. Mass propagation of Plectranthus bourneae Gamble through indirect organogenesis from leaf and internode explants. Physiol. Mol. Biol. Plants 2016, 22, 143–151. [Google Scholar] [CrossRef] [Green Version]
- Fonseka, D.; Wickramaarachchi, W.; Situge, C. Mass production of Plectranthus zeylanicus-A valuable medicinal and aromatic plant with a future value. Int. J. Minor Fruits Med. Aromat. Plants 2019, 5, 15–20. [Google Scholar]
- Mahmoud, D.S.; Sayed, L.M.; Diab, M.; Fahmy, E.M. In vitro propagation of Plectranthus barbatus andrews as important medicinal plant. Arab. Univ. J. Agric. Sci. 2019, 27, 511–517. [Google Scholar] [CrossRef] [Green Version]
- Kujeke, G.T.; Chitendera, T.C.; Masekesa, R.T.; Mazarura, U.; Ngadze, E.; Rugare, J.T.; Matikiti, A. Micropropagation of Livingstone Potato (Plectranthus esculentus N.E.Br). Adv. Agric. 2020, 2020, 8364153. [Google Scholar] [CrossRef]
- Hussain, S.A.; Ahmad, N.; Anis, M.; Hakeem, K.R. Development of an efficient micropropagation system for Tecoma stans (L.) Juss. ex Kunth using thidiazuron and effects on phytochemical constitution. In Vitro Cell. Dev. Biol.-Plant 2019, 55, 442–453. [Google Scholar] [CrossRef]
- Ahmad, N.; Faisal, M.; Ahmad, A.; Alatar, A.A.; Qahtan, A.A.; Alok, A. Thidiazuron Induced In Vitro Clonal Propagation of Lagerstroemia speciosa (L.) Pers.—An Important Avenue Tree. Horticulturae 2022, 8, 359. [Google Scholar] [CrossRef]
- Gharari, Z.; Bagheri, K.; Sharafi, A.; Danafar, H. Thidiazuron induced efficient in vitro organogenesis and regeneration of Scutellaria bornmuelleri: An important medicinal plant. In Vitro Cell. Dev. Biol.-Plant 2019, 55, 133–138. [Google Scholar] [CrossRef]
- Mok, M.; Mok, D.; Armstrong, D.; Shudo, K.; Isogai, Y.; Okamoto, T. Cytokinin activity of N-phenyl-N′-1, 2, 3-thiadiazol-5-ylurea (thidiazuron). Phytochemistry 1982, 21, 1509–1511. [Google Scholar] [CrossRef]
- Murthy, B.N.S.; Murch, S.J.; Saxena, P.K. Thidiazuron: A potent regulator of in vitro plant morphogenesis. In Vitro Cell. Dev. Biol.-Plant 1998, 34, 267. [Google Scholar] [CrossRef]
- Ċosiċ, T.; Motyka, V.; Raspor, M.; Savić, J.; Cingel, A.; Vinterhalter, B.; Vinterhalter, D.; Trávníčková, A.; Dobrev, P.I.; Bohanec, B.; et al. In vitro shoot organogenesis and comparative analysis of endogenous phytohormones in kohlrabi (Brassica oleracea var. gongylodes): Effects of genotype, explant type and applied cytokinins. Plant Cell Tissue Organ Cult. 2015, 121, 741–760. [Google Scholar] [CrossRef]
- Guo, B.; Abbasi, B.H.; Zeb, A.; Xu, L.; Wei, Y. Thidiazuron: A multi-dimensional plant growth regulator. Afr. J. Biotechnol. 2011, 10, 8984–9000. [Google Scholar] [CrossRef] [Green Version]
- Khan, I.; Khan, M.A.; Shehzad, M.A.; Ali, A.; Mohammad, S.; Ali, H.; Alyemeni, M.N.; Ahmad, P. Micropropagation and Production of Health Promoting Lignans in Linum usitatissimum. Plants 2020, 9, 728. [Google Scholar] [PubMed]
- Baskaran, P.; Ncube, B.; Van Staden, J. In vitro propagation and secondary product production by Merwilla plumbea (Lindl.) Speta. Plant Growth Regul. 2012, 67, 235–245. [Google Scholar] [CrossRef]
- Bhattacharyya, P.; Kumaria, S.; Diengdoh, R.; Tandon, P. Genetic stability and phytochemical analysis of the in vitro regenerated plants of Dendrobium nobile Lindl., an endangered medicinal orchid. Meta Gene 2014, 2, 489–504. [Google Scholar] [CrossRef]
- Moradi, A.; Zarinkamar, F.; Caretto, S.; Azadi, P. Influence of thidiazuron on callus induction and crocin production in corm and style explants of Crocus sativus L. Acta Physiol. Plant. 2018, 40, 185. [Google Scholar] [CrossRef]
- Anjum, S.; Abbasi, B.H. Thidiazuron-enhanced biosynthesis and antimicrobial efficacy of silver nanoparticles via improving phytochemical reducing potential in callus culture of Linum usitatissimum L. Int. J. Nanomed. 2016, 11, 715. [Google Scholar] [CrossRef] [Green Version]
- Huetteman, C.A.; Preece, J.E. Thidiazuron: A potent cytokinin for woody plant tissue culture. Plant Cell Tissue Organ Cult. 1993, 33, 105–119. [Google Scholar] [CrossRef]
- Faisal, M.; Ahmad, N.; Anis, M. Shoot multiplication in Rauvolfia tetraphylla L. using thidiazuron. Plant Cell Tissue Organ Cult. 2005, 80, 187–190. [Google Scholar] [CrossRef]
- Dewir, Y.H.; Nurmansyah; Naidoo, Y.; Teixeira da Silva, J.A. Thidiazuron-induced abnormalities in plant tissue cultures. Plant Cell Rep. 2018, 37, 1451–1470. [Google Scholar] [CrossRef]
- Akasaka, Y.; Daimon, H.; Mii, M. Improved plant regeneration from cultured leaf segments in peanut (Arachis hypogaea L.) by limited exposure to thidiazuron. Plant Sci. 2000, 156, 169–175. [Google Scholar] [CrossRef]
- Ahmad, N.; Srivastava, R.; Anis, M. Improvement in carnation shoot multiplication using thidiazuron in vitro. Propag. Ornam. Plants 2006, 6, 109–113. [Google Scholar]
- Kadota, M.; Niimi, Y. Effects of cytokinin types and their concentrations on shoot proliferation and hyperhydricity in in vitro pear cultivar shoots. Plant Cell Tissue Organ Cult. 2003, 72, 261–265. [Google Scholar] [CrossRef]
- Ivanova, M.; van Staden, J. Effect of ammonium ions and cytokinins on hyperhydricity and multiplication rate of in vitro regenerated shoots of Aloe polyphylla. Plant Cell Tissue Organ Cult. 2008, 92, 227–231. [Google Scholar] [CrossRef]
- Magioli, C.; Rocha, A.P.M.; de Oliveira, D.E.; Mansur, E. Efficient shoot organogenesis of eggplant (Solanum melongena L.) induced by thidiazuron. Plant Cell Rep. 1998, 17, 661–663. [Google Scholar] [CrossRef] [PubMed]
- Khoddamzadeh, A.A.; Sinniah, U.R.; Kadir, M.A.; Kadzimin, S.; Mahmood, M.; Sreeramanan, S. Detection of somaclonal variation by random amplified polymorphic DNA analysis during micropropagation of Phalaenopsis bellina (Rchb. f.) Christenson. Afr. J. Biotechnol. 2010, 9, 6632–6639. [Google Scholar]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Gamborg, O.L.; Miller, R.A.; Ojima, K. Nutrient requirements of suspension cultures of soybean root cells. Exp. Cell Res. 1968, 50, 151–158. [Google Scholar]
- Linsmaier, E.M.; Skoog, F. Organic growth factor requirements of tobacco tissue cultures. Physiol. Plant. 1965, 18, 100–127. [Google Scholar]
- Nitsch, J.P.; Nitsch, C. Haploid Plants from Pollen Grains. Science 1969, 163, 85–87. [Google Scholar] [CrossRef]
- Schenk, R.U.; Hildebrandt, A. Medium and techniques for induction and growth of monocotyledonous and dicotyledonous plant cell cultures. Can. J. Bot. 1972, 50, 199–204. [Google Scholar] [CrossRef]
- Galbraith, D.W.; Harkins, K.R.; Maddox, J.M.; Ayres, N.M.; Sharma, D.P.; Firoozabady, E. Rapid flow cytometric analysis of the cell cycle in intact plant tissues. Science 1983, 220, 1049–1051. [Google Scholar] [CrossRef]
- Doležel, J.; Bartoš, J. Plant DNA flow cytometry and estimation of nuclear genome size. Ann. Bot. 2005, 95, 99–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doležel, J.; Bartoš, J.; Voglmayr, H.; Greilhuber, J. Nuclear DNA Content and Genome Size of Trout and Human. Cytom. Part A 2003, 51, 127–128. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of folin-ciocalteu reagent. In Methods in Enzymology; Academic Press: Cambridge, MA, USA, 1999; Volume 299, pp. 152–178. [Google Scholar]
- Chandran, K.C.; Indira, G. Quantitative estimation of total phenolic, flavonoids, tannin and chlorophyll content of leaves of Strobilanthes Kunthiana (Neelakurinji). J. Med. Plants Stud. 2016, 4, 282–286. [Google Scholar]
- Ordoñez, A.A.L.; Gomez, J.D.; Vattuone, M.A.; lsla, M.I. Antioxidant activities of Sechium edule (Jacq.) Swartz extracts. Food Chem. 2006, 97, 452–458. [Google Scholar] [CrossRef]
- Liyana-Pathirana, C.M.; Shahidi, F. Antioxidant activity of commercial soft and hard wheat (Triticum aestivum L.) as affected by gastric pH conditions. J. Agric. Food Chem. 2005, 53, 2433–2440. [Google Scholar] [PubMed]
- Thomas, T.D. Pretreatment in thidiazuron improves the in vitro shoot induction from leaves in Curculigo orchioides Gaertn., an endangered medicinal plant. Acta Physiol. Plant. 2007, 29, 455–461. [Google Scholar] [CrossRef]
- Kereša, S.; Mihovilović, A.; Barić, M.; Jerčić, I.H. Efficient plant regeneration of Begonia semperflorens and Begonia spp. from petiole and leaf explants. J. Food Agric. Environ. 2011, 9, 240–244. [Google Scholar]
- Jahan, A.A.; Anis, M.; Aref, I.M. Preconditioning of Axillary Buds in Thidiazuron-Supplemented Liquid Media Improves In Vitro Shoot Multiplication in Nyctanthes arbor-tristis L. Appl. Biochem. Biotechnol. 2011, 163, 851–859. [Google Scholar] [CrossRef] [PubMed]
- Ivarson, E.; Ahlman, A.; Li, X.; Zhu, L.-H. Development of an efficient regeneration and transformation method for the new potential oilseed crop Lepidium campestre. BMC Plant Biol. 2013, 13, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faisal, M.; Alatar, A.A.; Hegazy, A.K.; Alharbi, S.A.; El-Sheikh, M.; Okla, M.K. Thidiazuron induced in vitro multiplication of Mentha arvensis and evaluation of genetic stability by flow cytometry and molecular markers. Ind. Crops Prod. 2014, 62, 100–106. [Google Scholar] [CrossRef]
- Faisal, M.; Alatar, A.A.; El-Sheikh, M.A.; Abdel-Salam, E.M.; Qahtan, A.A. Thidiazuron induced in vitro morphogenesis for sustainable supply of genetically true quality plantlets of Brahmi. Ind. Crops Prod. 2018, 118, 173–179. [Google Scholar] [CrossRef]
- Novikova, T.I.; Asbaganov, S.V.; Ambros, E.V.; Zaytseva, Y.G. TDZ-induced axillary shoot proliferation of Rhododendron mucronulatum Turcz and assessment of clonal fidelity using DNA-based markers and flow cytometry. In Vitro Cell. Dev. Biol.-Plant 2020, 56, 307–317. [Google Scholar] [CrossRef]
- Khalafalla, M.M.; Hattori, K. A combination of thidiazuron and benzyladenine promotes multiple shoot production from cotyledonary node explants of faba bean (Vicia faba L.). Plant Growth Regul. 1999, 27, 145–148. [Google Scholar] [CrossRef]
- Capelle, S.C.; Mok, D.W.; Kirchner, S.C.; Mok, M.C. Effects of thidiazuron on cytokinin autonomy and the metabolism of N 6-(Δ2-isopentenyl)[8-14C] adenosine in callus tissues of Phaseolus lunatus L. Plant Physiol. 1983, 73, 796–802. [Google Scholar] [CrossRef] [Green Version]
- Debergh, P.C.; Maene, L.J. A scheme for commercial propagation of ornamental plants by tissue culture. Sci. Hortic. 1981, 14, 335–345. [Google Scholar] [CrossRef]
- Agretious, T.K.; Martin, K.; Hariharan, M. In vitro clonal multiplication of Alpinia calcarata Rosc. Phytomorphology 1996, 46, 133–138. [Google Scholar]
- Slazak, B.; Sliwinska, E.; Saługa, M.; Ronikier, M.; Bujak, J.; Słomka, A.; Göransson, U.; Kuta, E. Micropropagation of Viola uliginosa (Violaceae) for endangered species conservation and for somaclonal variation-enhanced cyclotide biosynthesis. Plant Cell Tissue Organ Cult. 2015, 120, 179–190. [Google Scholar] [CrossRef] [Green Version]
- Matkowski, A. Plant in vitro culture for the production of antioxidants—A review. Biotechnol. Adv. 2008, 26, 548–560. [Google Scholar] [CrossRef]
- Abbasi, B.H.; Khan, M.A.; Mahmood, T.; Ahmad, M.; Chaudhary, M.F.; Khan, M.A. Shoot regeneration and free-radical scavenging activity in Silybum marianum L. Plant Cell Tissue Organ Cult. 2010, 101, 371–376. [Google Scholar] [CrossRef]
- Baskaran, P.; Moyo, M.; Van Staden, J. In vitro plant regeneration, phenolic compound production and pharmacological activities of Coleonema pulchellum. S. Afr. J. Bot. 2014, 90, 74–79. [Google Scholar] [CrossRef] [Green Version]
- Karamać, M. Chelation of Cu(II), Zn(II), and Fe(II) by Tannin Constituents of Selected Edible Nuts. Int. J. Mol. Sci. 2009, 10, 5485–5497. [Google Scholar] [CrossRef] [PubMed]
- Chavan, J.J.; Gaikwad, N.B.; Umdale, S.D.; Kshirsagar, P.R.; Bhat, K.V.; Yadav, S.R. Efficiency of direct and indirect shoot organogenesis, molecular profiling, secondary metabolite production and antioxidant activity of micropropagated Ceropegia santapaui. Plant Growth Regul. 2014, 72, 1–15. [Google Scholar] [CrossRef]
- Thiruvengadam, M.; Chung, I.-M. Phenolic compound production and biological activities from in vitro regenerated plants of gherkin (Cucumis anguria L.). Electron. J. Biotechnol. 2015, 18, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Rawat, J.M.; Bhandari, A.; Mishra, S.; Rawat, B.; Dhakad, A.K.; Thakur, A.; Chandra, A. Genetic stability and phytochemical profiling of the in vitro regenerated plants of Angelica glauca Edgew.: An endangered medicinal plant of Himalaya. Plant Cell Tissue Organ Cult. 2018, 135, 111–118. [Google Scholar] [CrossRef]
- Qahtan, A.A.; Faisal, M.; Alatar, A.A.; Abdel-Salam, E.M. Callus-Mediated High-Frequency Plant Regeneration, Phytochemical Profiling, Antioxidant Activity and Genetic Stability in Ruta chalepensis L. Plants 2022, 11, 1614. [Google Scholar] [CrossRef]
- López-Laredo, A.R.; Ramírez-Flores, F.D.; Sepúlveda-Jiménez, G.; Trejo-Tapia, G. Comparison of metabolite levels in callus of Tecoma stans (L.) Juss. ex Kunth. cultured in photoperiod and darkness. In Vitro Cell. Dev. Biol. Plant 2009, 45, 550–558. [Google Scholar] [CrossRef]
- Upadhyay, A.; Shahzad, A.; Ahmad, Z.; Alatar, A.A.; Guerriero, G.; Faisal, M. High frequency direct organogenesis, genetic homogeneity, chemical characterization and leaf ultra-structural study of regenerants in Diplocyclos palmatus (L.) C. Jeffrey. Agronomy 2021, 11, 2164. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Thidiazuron (μM) | % Regeneration | Number of Shoots Per Explant | Shoot Length (cm) |
|---|---|---|---|
| 0.0 | 23.25 ± 1.25 e | 1.50 ± 0.57 e | 2.87 ± 0.42 cd |
| 0.5 | 46.30 ± 2.50 d | 6.57 ± 0.95 d | 3.55 ± 0.33 bc |
| 5.0 | 68.25 ± 2.36 c | 12.25 ± 2.06 c | 4.02 ± 0.23 ab |
| 25.0 | 97.15 ± 2.90 a | 27.30 ± 1.91 a | 4.93 ± 0.28 a |
| 50.0 | 87.50 ± 2.80 b | 18.25 ± 1.70 b | 3.17 ± 0.19 bcd |
| 100.0 | 65.95 ± 3.55 c | 11.00 ± 1.29 c | 2.50 ± 0.17 d |
| Phytoconstituents | In Vitro Plants | Ex Vitro Plants |
|---|---|---|
| Total phenolic (mg GAE/g DW) | 81.23 ± 0.72 a | 56.64 ± 0.31 b |
| Total Tannin (mg TAE/g DW) | 55.68 ± 0.48 a | 23.41 ± 52 b |
| Total Flavonoid (mg QE/g DW) | 42.77 ± 0.43 a | 40.27 ± 0.39 a |
| Name | Retention Time (min) | Area % | Formula | Molecular Weight (g/mol) | ||
|---|---|---|---|---|---|---|
| Ex Vitro | In Vitro | Ex Vitro | In Vitro | |||
| Beta-myrcene | 5.03 | 5.05 | 0.08 | 0.04 | C10H16 | 136.23 |
| 1-methyl-2-(1-methylethyl)-benzene | 5.38 | 5.39 | 1.54 | 2.38 | C10H14 | 134.22 |
| Gamma-terpinene | 5.66 | 5.67 | 2.65 | 4.75 | C10H16 | 136.23 |
| 3-Hydroxy-2-methyl-4H-pyran-4-one | - | 6.11 | - | 0.03 | C6H6O3 | 126.11 |
| Cyclononanone | 6.24 | 6.24 | 0.1 | 0.13 | C9H16O | 140.22 |
| Terpinen-4-ol | 6.7 | 6.69 | 0.11 | 0.12 | C10H18O | 154.25 |
| 5-(Hydroxymethyl)-2-furancarboxaldehyde | - | 6.96 | - | 0.25 | C6H6O3 | 126.11 |
| 2-Furanmethanol | 6.98 | - | 0.31 | - | C5H6O2 | 98.1 |
| 2,5-Cyclohexadiene-1,4-dione | 7.2 | 7.18 | 0.07 | 0.05 | C6H4O2 | 108.09 |
| 5-Methyl-2-(1-methylethyl)-phenol | 7.5 | 7.45 | 91.38 | 86.69 | C10H14O | 150.22 |
| Alpha-copaene | 8.2 | 8.14 | 0.12 | 0.18 | C15H24 | 204.36 |
| Trans-caryophyllene | 8.54 | 8.47 | 1.88 | 2.2 | C15H24 | 204.35 |
| Alpha-humulene | - | 8.7 | - | 0.22 | C15H24 | 204.35 |
| Trans ocimene | 8.8 | - | 0.17 | - | C10H16 | 136.23 |
| Aromadendrene | - | 8.91 | - | 0.34 | C15H24 | 204.35 |
| Delta-cadinene | 9.22 | 9.04 | 0.09 | 0.11 | C15H24 | 204.35 |
| (-)-Curcuhydroquinone | 9.49 | - | 0.23 | - | C15H22O2 | 234.16 |
| Caryophyllene oxide | - | 9.51 | - | 0.19 | C15H24O | 220.35 |
| (-)-Caryophyllene oxide | 9.74 | - | 0.26 | - | C15H24O | 220.35 |
| Hybridalactone | 10.14 | - | 0.08 | - | C21H30O3 | 330.5 |
| cis-(Z)-alpha-Bisabolene epoxide | 10.28 | - | 0.04 | - | C15H24O | 220.35 |
| trans-Z- alpha-Bisabolene epoxide | 10.4 | - | 0.03 | - | C15H24O | 220.35 |
| BIS[(1S,2S,3S,5R)-(+)-isopinocamphyl] phosphorochloridate | - | 11.15 | - | 0.16 | C10H18O | 154.25 |
| Hexadecanoic acid | - | 11.4 | - | 0.51 | C16H32O2 | 256.42 |
| 3,7,11,15-Tetramethyl 2-hexadecen-1-ol | - | 11.59 | - | 0.02 | C20H40O | 296.53 |
| 10-Heneicosene | - | 12.24 | - | 0.32 | C21H42 | 294.56 |
| 9,12,15-Octadecatrienoic acid | - | 12.4 | - | 0.59 | C18H30O2 | 278.43 |
| Octadecanoic acid | - | 12.52 | - | 0.1 | C18H36O2 | 284.48 |
| 5-Methyl-2-(1-methylethylidene) cyclohexanone | 12.56 | - | 0.16 | - | C10H16O | 152.23 |
| Tridecanoic Acid | 15.12 | - | 0.1 | - | C13H26O2 | 214.34 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Faisal, M.; Qahtan, A.A.; Alatar, A.A. Thidiazuron Induced In Vitro Plant Regeneration, Phenolic Contents, Antioxidant Potential, GC-MS Profiles and Nuclear Genome Stability of Plectranthus amboinicus (Lour.) Spreng. Horticulturae 2023, 9, 277. https://doi.org/10.3390/horticulturae9020277
Faisal M, Qahtan AA, Alatar AA. Thidiazuron Induced In Vitro Plant Regeneration, Phenolic Contents, Antioxidant Potential, GC-MS Profiles and Nuclear Genome Stability of Plectranthus amboinicus (Lour.) Spreng. Horticulturae. 2023; 9(2):277. https://doi.org/10.3390/horticulturae9020277
Chicago/Turabian StyleFaisal, Mohammad, Ahmed A. Qahtan, and Abdulrahman A. Alatar. 2023. "Thidiazuron Induced In Vitro Plant Regeneration, Phenolic Contents, Antioxidant Potential, GC-MS Profiles and Nuclear Genome Stability of Plectranthus amboinicus (Lour.) Spreng" Horticulturae 9, no. 2: 277. https://doi.org/10.3390/horticulturae9020277