
Exogenous Application of Melatonin Alleviates Drought Stress in Ranunculus asiaticus by Improving Its Morphophysiological and Biochemical Attributes



Abstract
:1. Introduction
2. Materials and Methods
2.1. Material and Conditions for Planting
2.2. Experimental Design and Irrigation Treatments
2.3. Morphological Characteristics
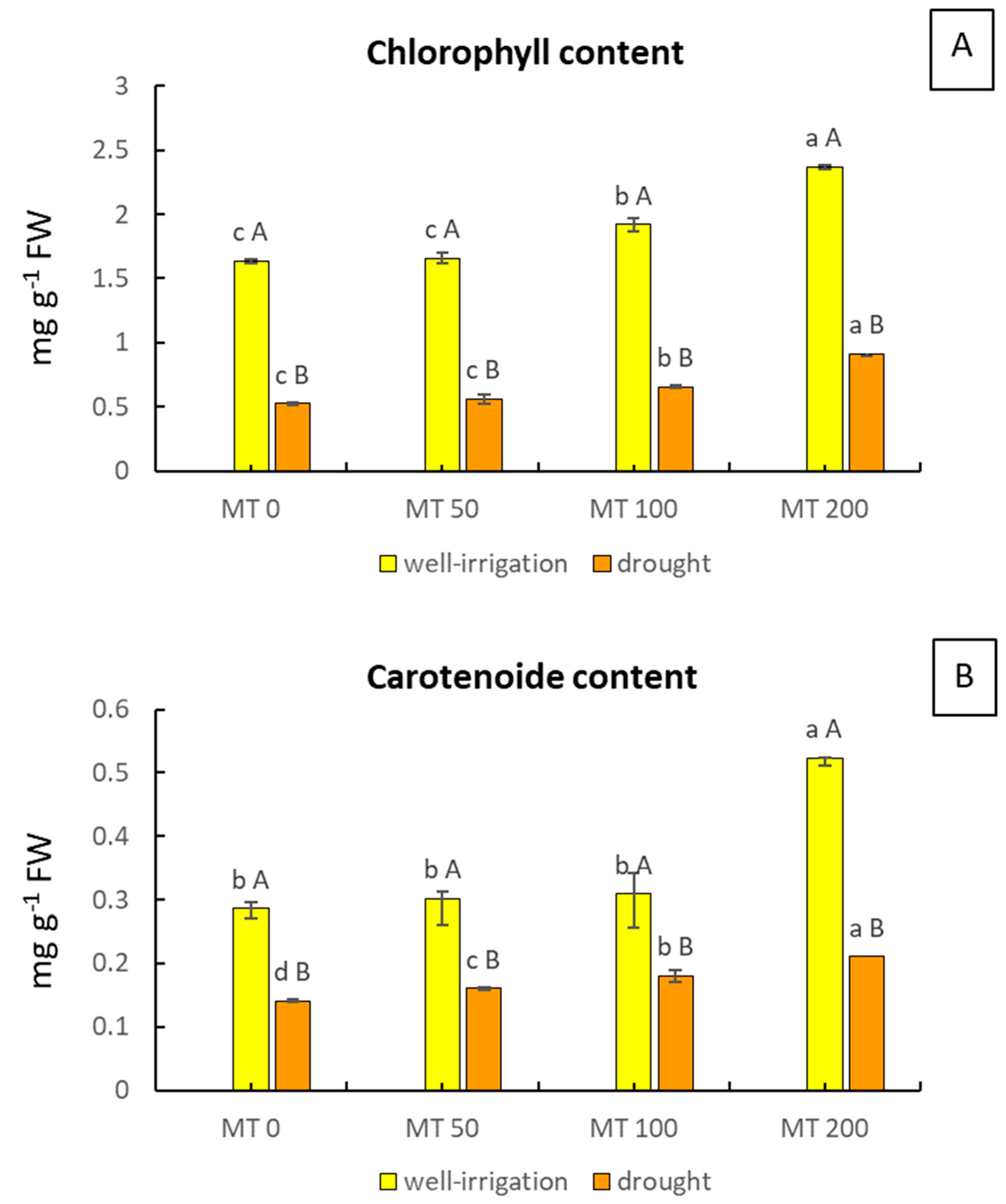
2.4. Photosynthetic Pigments Analysis
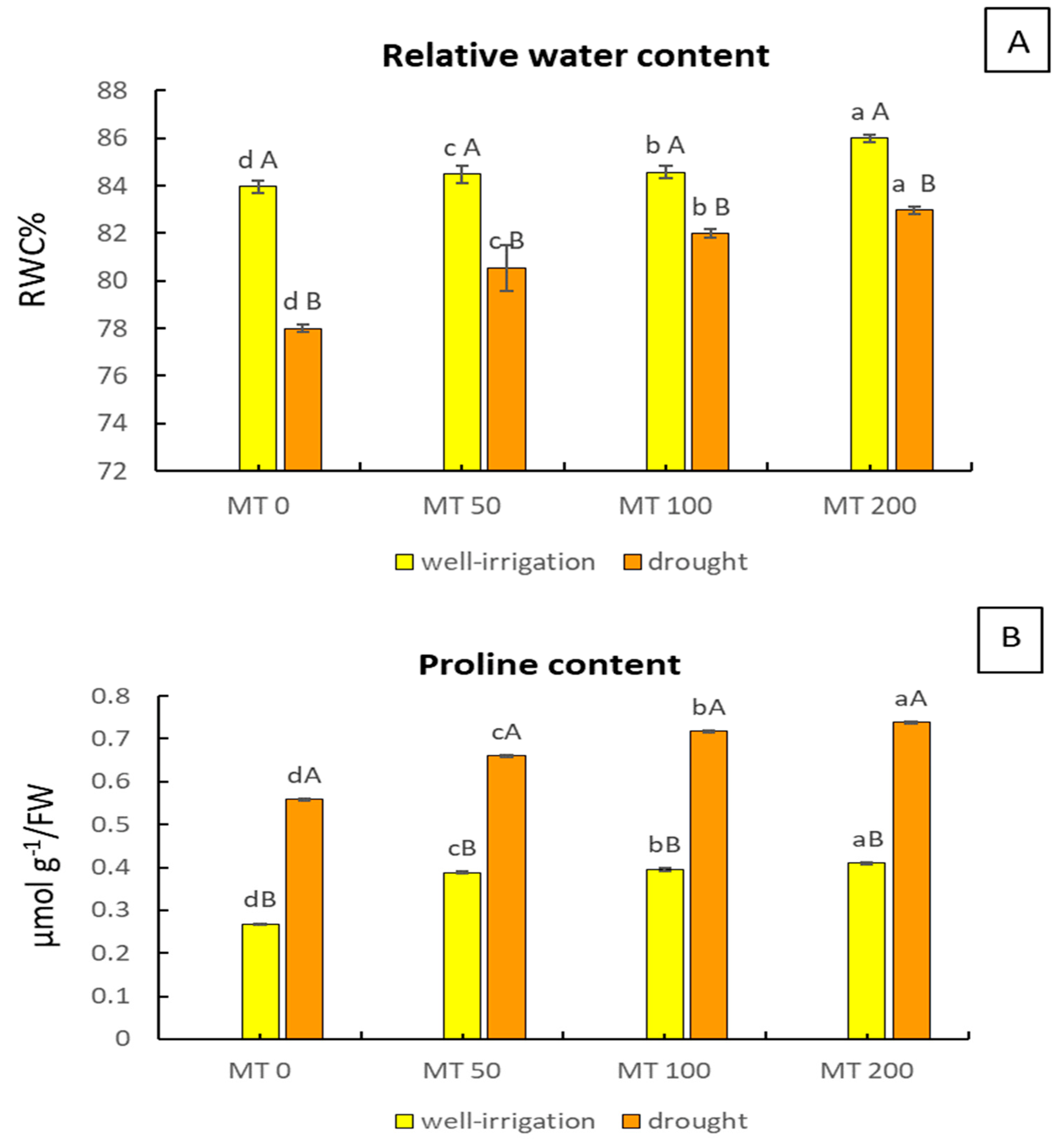
2.5. RWC in Leaves
2.6. Electrolyte Leakage
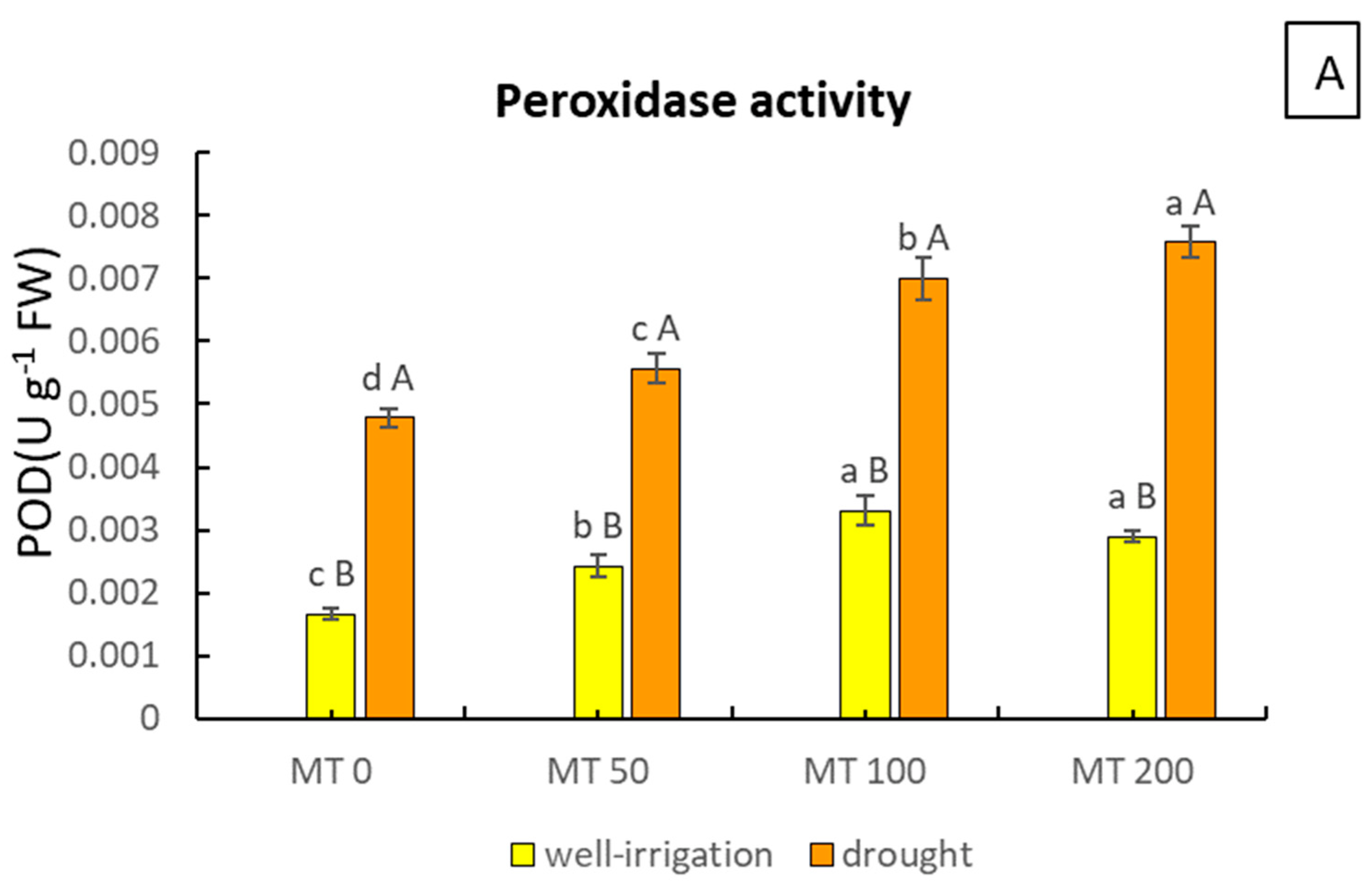
2.7. POD Activity Determination
2.8. Proline Content
2.9. DTI
2.10. Statistical Analyses
3. Results
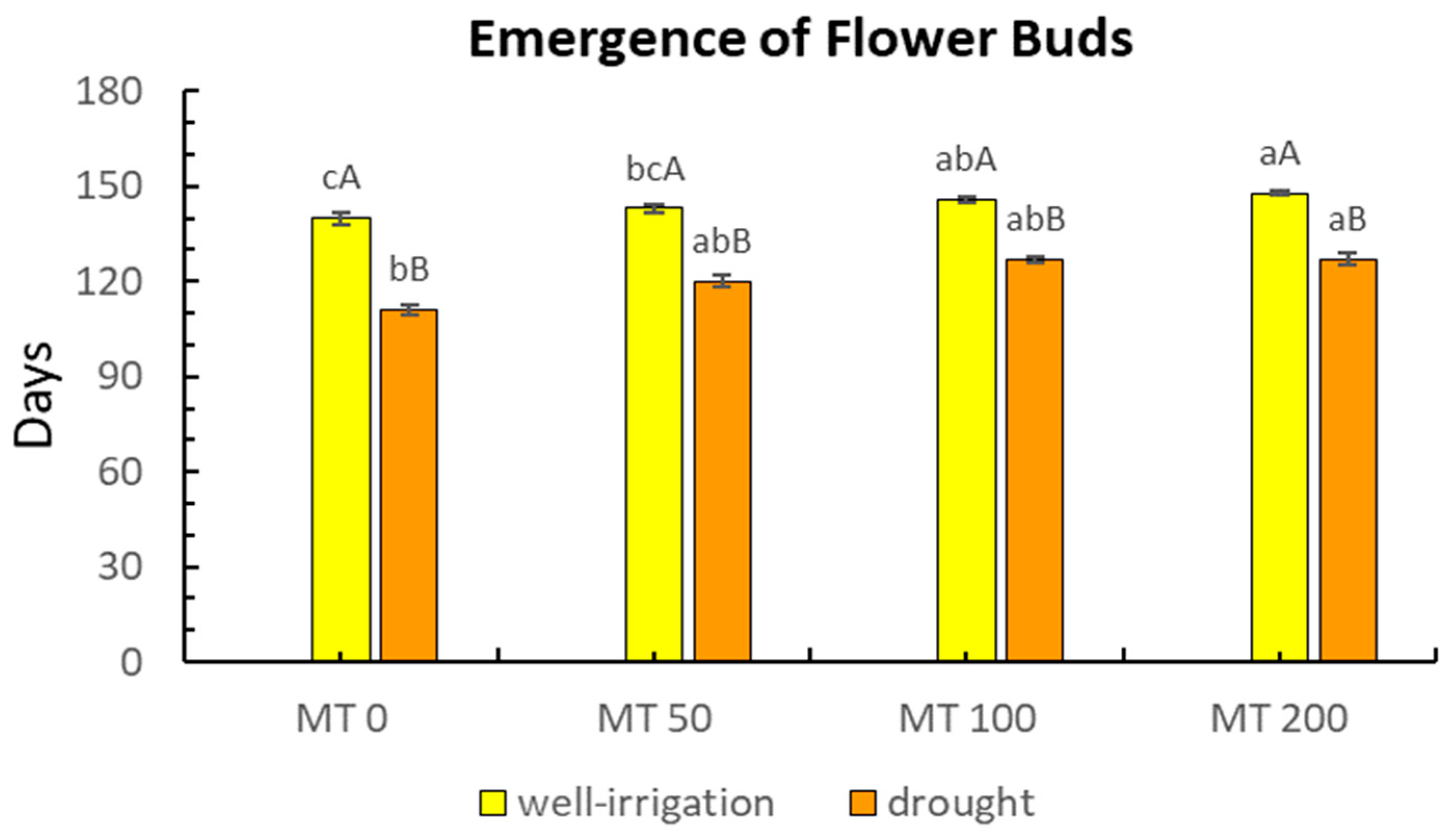
3.1. Impact of MT on Morphology under Drought Conditions
3.2. Changes in Photosynthetic Pigments
3.3. Exogenous MT Alters the RWC and Proline Content under Drought Stress
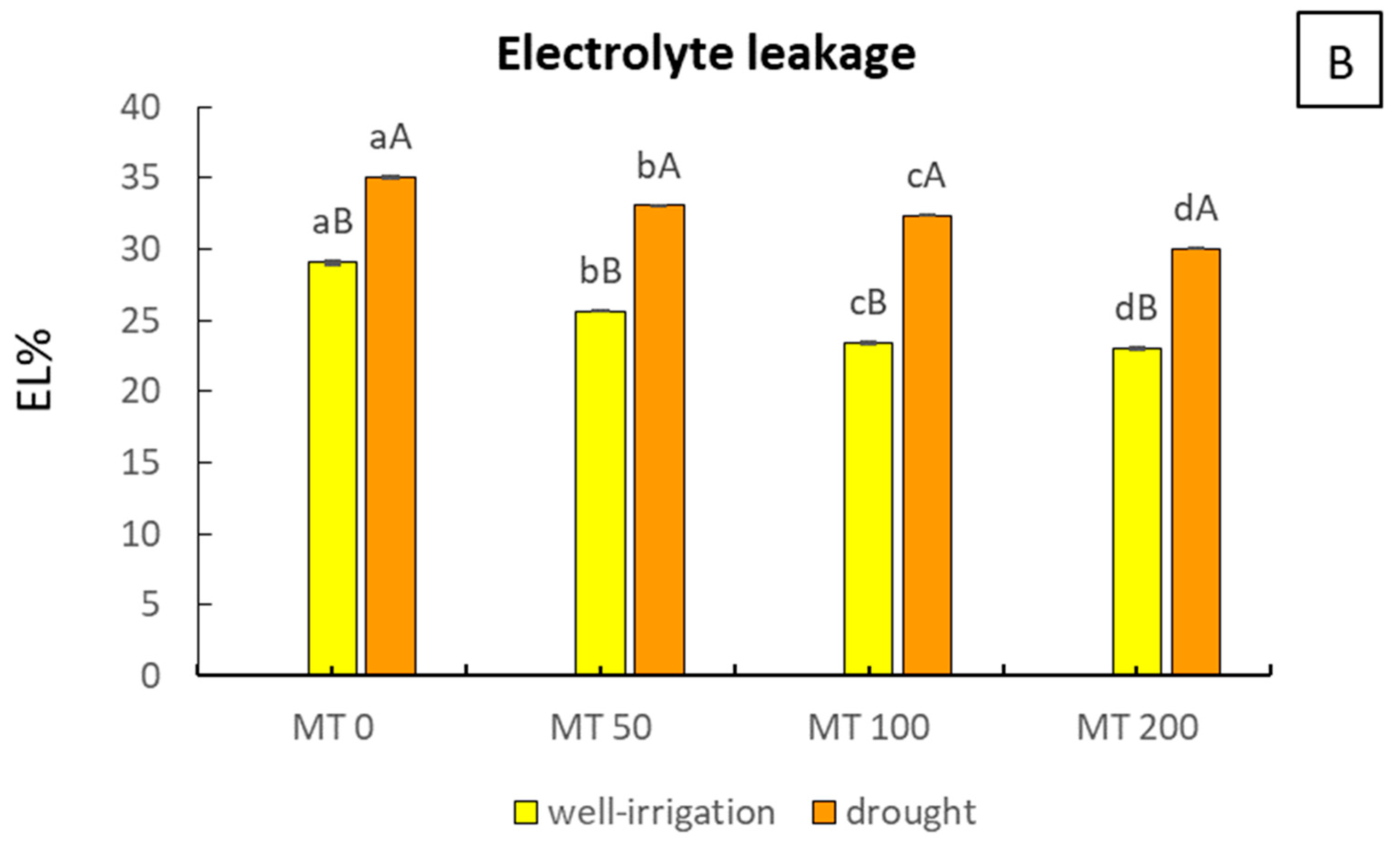
3.4. MT Modulates the Activity of Peroxidase Enzymes and Checks EL
3.5. DTI
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Karlsson, M. Producing Ravishing Ranunculus; Greenhouse Product News: Sparta, MI, USA, 2003; pp. 44–48. [Google Scholar]
- Margherita, B.; Giampiero, C.; Pierre, D. Field performance of tissue-cultured plants of Ranunculus asiaticus L. Sci. Hortic. 1996, 66, 229–239. [Google Scholar] [CrossRef]
- Beruto, M.; Rabaglio, M.; Viglione, S.; van Labeke, M.-C.; Dhooghe, E. Ranunculus. In Ornamental Crops; Springer: Berlin, Germany, 2018; pp. 649–671. [Google Scholar]
- Hashemi, G.S.E.; Mostafa, Z.B.; Heidarzadeh, M. Estimation of water requirement about some of the most dominant green areas in isfahan, using lysimeter. In Proceedings of the Third National Conference on Green Spaces and Urban Landscape, Kish Island, Municipality and Degradation Organization of the Country; Elsevier: Amsterdam, The Netherlands, 2007. (In Persian) [Google Scholar]
- Khan, F.; Upreti, P.; Singh, R.; Shukla, P.K.; Shirke, P.A. Physiological performance of two contrasting rice varieties under water stress. Physiol. Mol. Biol. Plants 2016, 23, 85–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tombesi, S.; Frioni, T.; Poni, S.; Palliotti, A. Effect of water stress “memory” on plant behavior during subsequent drought stress. Environ. Exp. Bot. 2018, 150, 106–114. [Google Scholar] [CrossRef]
- Caser, M.; Chitarra, W.; D’Angiolillo, F.; Perrone, I.; Demasi, S.; Lovisolo, C.; Pistelli, L.; Pistelli, L.; Scariot, V. Drought stress adaptation modulates plant secondary metabolite production in Salvia dolomitica Codd. Ind. Crop. Prod. 2019, 129, 85–96. [Google Scholar] [CrossRef]
- Bidabadi, S.S.; VanderWeide, J.; Sabbatini, P. Exogenous melatonin improves glutathione content, redox state and increases essential oil production in two Salvia species under drought stress. Sci. Rep. 2020, 10, 6883. [Google Scholar] [CrossRef] [Green Version]
- Rafi, Z.N.; Kazemi, F.; Tehranifar, A. Morpho-physiological and biochemical responses of four ornamental herbaceous species to water stress. Acta Physiol. Plant. 2018, 41, 7. [Google Scholar] [CrossRef]
- Altaf, M.A.; Shahid, R.; Ren, M.-X.; Naz, S.; Altaf, M.M.; Khan, L.U.; Tiwari, R.K.; Lal, M.K.; Shahid, M.A.; Kumar, R.; et al. Melatonin improves drought stress tolerance of tomato by modulating plant growth, root architecture, photosynthesis, and antioxidant defense system. Antioxidants 2022, 11, 309. [Google Scholar] [CrossRef]
- Anjum, S.A.; Tanveer, M.; Ashraf, U.; Hussain, S.; Shahzad, B.; Khan, I.; Wang, L. Effect of progressive drought stress on growth, leaf gas exchange, and antioxidant production in two maize cultivars. Environ. Sci. Pollut. Res. 2016, 23, 17132–17141. [Google Scholar] [CrossRef]
- Sadak, M.S.; Abdalla, A.M.; Elhamid, E.M.A.; Ezzo, M.I. Role of melatonin in improving growth, yield quantity and quality of Moringa oleifera L. plant under drought stress. Bull. Natl. Res. Cent. 2020, 44, 18. [Google Scholar] [CrossRef] [Green Version]
- Abdul Jaleel, C.; Manivannan, P.; Wahid, A.; Farooq, M.; Al-Juburi, H.J.; Somasundaram, R.; Panneerselvam, R. Drought stress in plants: A review on morphological characteristics and pigments composition. Int. J. Agric. Biol. 2009, 11, 100–105. [Google Scholar]
- Munné-Bosch, S.; Mueller, M.; Schwarz, K.; Alegre, L. Diterpenes and antioxidative protection in drought-stressed Salvia officinalis plants. J. Plant Physiol. 2001, 158, 1431–1437. [Google Scholar] [CrossRef]
- Fan, J.; Xie, Y.; Zhang, Z.; Chen, L. Melatonin: A multifunctional factor in plants. Int. J. Mol. Sci. 2018, 19, 1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maksup, S.; Roytrakul, S.; Supaibulwatana, K. Physiological and comparative proteomic analyses of and two check cultivars in response to drought stress. J. Plant Interact. 2012, 9, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Jafari, S.; Garmdareh, S.E.H.; Azadegan, B. Effects of drought stress on morphological, physiological, and biochemical characteristics of stock plant (Matthiola incana L.). Sci. Hortic. 2019, 253, 128–133. [Google Scholar] [CrossRef]
- Oraee, A.; Tehranifar, A. Evaluating the potential drought tolerance of pansy through its physiological and biochemical responses to drought and recovery periods. Sci. Hortic. 2020, 265, 109225. [Google Scholar] [CrossRef]
- Tan, D.-X.; Manchester, L.C.; Liu, X.; Rosales-Corral, S.A.; Acuna-Castroviejo, D.; Reiter, R.J. Mitochondria and chloroplasts as the original sites of melatonin synthesis: A hypothesis related to melatonin’s primary function and evolution in eukaryotes. J. Pineal Res. 2012, 54, 127–138. [Google Scholar] [CrossRef]
- Sarropoulou, V.N.; Therios, I.N.; Dimassi-Theriou, K.N. Melatonin promotes adventitious root regeneration in in vitro shoot tip explants of the commercial sweet cherry rootstocks CAB-6P (Prunus cerasus L.), Gisela 6 (P. cerasus × P. canescens), and MxM 60 (P. avium × P. mahaleb). J. Pineal Res. 2011, 52, 38–46. [Google Scholar] [CrossRef]
- Shi, H.; Jiang, C.; Ye, T.; Tan, D.-X.; Reiter, R.J.; Zhang, H.; Liu, R.; Chan, Z. Comparative physiological, metabolomic, and transcriptomic analyses reveal mechanisms of improved abiotic stress resistance in bermudagrass [Cynodon dactylon (L). Pers.] by exogenous melatonin. J. Exp. Bot. 2014, 66, 681–694. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zeng, L.; Cheng, Y.; Lu, G.; Fu, G.; Ma, H.; Liu, Q.; Zhang, X.; Zou, X.; Li, C. Exogenous melatonin alleviates damage from drought stress in Brassica napus L. (rapeseed) seedlings. Acta Physiol. Plant. 2018, 40, 43. [Google Scholar] [CrossRef]
- Eisa, E.A.; Honfi, P.; Tilly-Mándy, A.; Mirmazloum, I. Exogenous melatonin application induced morpho-physiological and biochemical regulations conferring salt tolerance in Ranunculus asiaticus L. Horticulturae 2023, 9, 228. [Google Scholar] [CrossRef]
- Wang, F.; Zeng, B.; Sun, Z.; Zhu, C. Relationship between proline and Hg2+-induced oxidative stress in a tolerant rice mutant. Arch. Environ. Contam. Toxicol. 2008, 56, 723–731. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Llorca, M.; Muñoz, P.; Müller, M.; Munné-Bosch, S. Biosynthesis, metabolism and function of auxin, salicylic acid and melatonin in climacteric and non-climacteric fruits. Front. Plant Sci. 2019, 10, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, C.; Zheng, G.; Li, W.; Wang, Y.; Hu, B.; Wang, H.; Wu, H.; Qian, Y.; Zhu, X.-G.; Tan, D.-X.; et al. Melatonin delays leaf senescence and enhances salt stress tolerance in rice. J. Pineal Res. 2015, 59, 91–101. [Google Scholar] [CrossRef]
- Gul, N.; Haq, Z.U.; Ali, H.; Munsif, F.; Hassan, S.S.U.; Bungau, S. Melatonin pretreatment alleviated inhibitory effects of drought stress by enhancing anti-oxidant activities and accumulation of higher proline and plant pigments and improving maize productivity. Agronomy 2022, 12, 2398. [Google Scholar] [CrossRef]
- Mushtaq, N.; Iqbal, S.; Hayat, F.; Raziq, A.; Ayaz, A.; Zaman, W. Melatonin in micro-tom tomato: Improved drought tolerance via the regulation of the photosynthetic apparatus, membrane stability, osmoprotectants, and root system. Life 2022, 12, 1922. [Google Scholar] [CrossRef]
- Langaroudi, I.K.; Piri, S.; Chaeikar, S.S.; Salehi, B. Evaluating drought stress tolerance in different Camellia sinensis L. cultivars and effect of melatonin on strengthening antioxidant system. Sci. Hortic. 2023, 307, 111517. [Google Scholar] [CrossRef]
- Zhang, Y.P.; Yang, S.J.; Chen, Y.Y. Effects of melatonin on photosynthetic performance and antioxidants in melon during cold and recovery. Biol. Plant. 2017, 61, 571–578. [Google Scholar] [CrossRef]
- Li, H.; Chang, J.; Chen, H.; Wang, Z.; Gu, X.; Wei, C.; Zhang, Y.; Ma, J.; Yang, J.; Zhang, X. Exogenous melatonin confers salt stress tolerance to watermelon by improving photosynthesis and redox homeostasis. Front. Plant Sci. 2017, 8, 295. [Google Scholar] [CrossRef] [Green Version]
- Naz, M.; Hussain, S.; Ashraf, I.; Farooq, M. Exogenous application of proline and phosphorus help improving maize performance under salt stress. J. Plant Nutr. 2022, 1–9. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Analysis 1983, 11, 591–592. [Google Scholar] [CrossRef] [Green Version]
- Turk, H.; Erdal, S. Melatonin alleviates cold-induced oxidative damage in maize seedlings by up-regulating mineral elements and enhancing antioxidant activity. J. Plant Nutr. Soil Sci. 2015, 178, 433–439. [Google Scholar] [CrossRef]
- Reddy, A.R.; Chaitanya, K.V.; Vivekanandan, M. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J. Plant Physiol. 2004, 161, 1189–1202. [Google Scholar] [CrossRef] [PubMed]
- He, J.; Ren, Y.; Chen, X.; Chen, H. Protective roles of nitric oxide on seed germination and seedling growth of rice (Oryza sativa L.) under cadmium stress. Ecotoxicol. Environ. Saf. 2014, 108, 114–119. [Google Scholar] [CrossRef] [PubMed]
- Ábrahám, E.; Hourton-Cabassa, C.; Erdei, L.; Szabados, L. Methods for determination of proline in plants. Plant Stress Toler. Methods Protoc. 2010, 639, 317–331. [Google Scholar] [CrossRef]
- Sbei, H.; Shehzad, T.; Harrabi, M.; Okuno, K. Salinity tolerance evaluation of Asian barley accessions (Hordeum vulgare L.) at the early vegetative stage. J. Arid Land Stud. 2014, 24, 183–186. [Google Scholar]
- West, S.G.; Finch, J.F.; Curran, P.J. Structural equation models with nonnormal variables: Problems and remedies. In Structural Equation Modeling: Concepts, Issues, and Applications; Hoyle, R.H., Ed.; Sage Publications, Inc.: Thousand Oaks, CA, USA, 1995; pp. 56–75. [Google Scholar]
- Brown, M.B.; Forsythe, A.B. Robust tests for the equality of variances. J. Am. Stat. Assoc. 1974, 69, 364–367. [Google Scholar] [CrossRef]
- Garson, G.D. Testing Statistical Assumptions; Statistical Associates Publishing: Asheboro, NC, USA, 2012. [Google Scholar]
- Tabachnick, B.; Fidell, L. IBM SPSS statistics for windows. In Using Multivariate Statistics; Pearson: Boston, MA, USA, 2020. [Google Scholar]
- Armonk, N.Y. IBM Corp. Released 2020. IBM SPSS Statistics for Windows, Version 27.0; IBM Corp.: Armonk, NY, USA, 2020. [Google Scholar]
- Olson, C.L. On choosing a test statistic in multivariate analysis of variance. Psychol. Bull. 1976, 83, 579. [Google Scholar] [CrossRef]
- Barbara, G.T.; Linda, S.F. Using Multivariate Statistics, 7th ed.; Pearson: New York, NY, USA, 2013; ISBN 9780135350904. [Google Scholar]
- Sahin, U.; Ekinci, M.; Ors, S.; Turan, M.; Yildiz, S.; Yildirim, E. Effects of individual and combined effects of salinity and drought on physiological, nutritional and biochemical properties of cabbage (Brassica oleracea var. capitata). Sci. Hortic. 2018, 240, 196–204. [Google Scholar] [CrossRef]
- Zhang, H.-J.; Zhang, N.; Yang, R.-C.; Wang, L.; Sun, Q.-Q.; Li, D.-B.; Cao, Y.-Y.; Weeda, S.; Zhao, B.; Ren, S.; et al. Melatonin promotes seed germination under high salinity by regulating antioxidant systems, ABA and GA4 interaction in cucumber (Cucumis sativus L.). J. Pineal Res. 2014, 57, 269–279. [Google Scholar] [CrossRef]
- Zhang, N.; Sun, Q.; Zhang, H.; Cao, Y.; Weeda, S.; Ren, S.; Guo, Y.-D. Roles of melatonin in abiotic stress resistance in plants. J. Exp. Bot. 2014, 66, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Liu, M.; Li, M.; Liu, K.; Sui, N. Effects of drought stress on seed germination and seedling growth of different maize varieties. J. Agric. Sci. 2015, 7, 231. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Wang, Z.; Kim, W.S. Effect of drought stress on shoot growth and physiological response in the cut rose ‘charming black’ at different developmental stages. Hortic. Environ. Biotechnol. 2018, 60, 1–8. [Google Scholar] [CrossRef]
- Elewa, T.A.; Sadak, M.S.; Saad, A.M. Proline treatment improves physiological responses in quinoa plants under drought stress. Biosci. Res. 2017, 14, 21–33. [Google Scholar]
- Sadiq, M.; Akram, N.A.; Ashraf, M. Impact of exogenously applied tocopherol on some key physio-biochemical and yield attributes in mungbean [Vigna radiata (L.) Wilczek] under limited irrigation regimes. Acta Physiol. Plant. 2018, 40, 131. [Google Scholar] [CrossRef]
- Dawood, M.G.; El-Awadi, M.E.; Sadak, M.S.; El-Lethy, S.R. Comparison between the physiological role of carrot root extract and β-carotene in inducing Helianthus annuus L. drought tolerance. Asian J. Biol. Sci 2019, 12, 231–241. [Google Scholar]
- Dawood, M.G.; Sadak, M.S. Physiological role of glycinebetaine in alleviating the deleterious effects of drought stress on canola plants (Brassica napus L.). Middle East J. Agric. Res. 2014, 3, 943–954. [Google Scholar]
- Bañon, S.; Ochoa, J.; Franco, J.; Alarcón, J.; Sánchez-Blanco, M. Hardening of oleander seedlings by deficit irrigation and low air humidity. Environ. Exp. Bot. 2006, 56, 36–43. [Google Scholar] [CrossRef]
- Bakry, B.A.; El-Hariri, D.M.; Sadak, M.S.; El-Bassiouny, H.M.S. Drought stress mitigation by foliar application of salicylic acid in two linseed varieties grown under newly reclaimed sandy soil. J. Appl. Sci. Res. 2012, 8, 3503–3514. [Google Scholar]
- Alam, M.; Nahar, K.; Hasanuzzaman, M.; Fujita, M. Trehalose-induced drought stress tolerance: A comparative study among different Brassica species. Plant Omics 2014, 7, 271–283. [Google Scholar]
- Naeem, M.; Ahmad, R.; Ahmad, R.; Ashraf, M.Y.; Ihsan, M.Z.; Nawaz, F.; Athar, H.-U.; Abbas, H.T.; Abdullah, M. Improving drought tolerance in maize by foliar application of boron: Water status, antioxidative defense and photosynthetic capacity. Arch. Agron. Soil Sci. 2017, 64, 626–639. [Google Scholar] [CrossRef]
- Szafrańska, K.; Reiter, R.J.; Posmyk, M.M. Melatonin application to Pisum sativum L. seeds positively influences the function of the photosynthetic apparatus in growing seedlings during paraquat-induced oxidative stress. Front. Plant Sci. 2016, 7, 1663. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Q.; Huang, B.; Ding, C.; Zhang, Z.; Chen, Y.; Hu, C.; Zhou, L.; Huang, Y.; Liao, J.; Yuan, S.; et al. Osystem II in cold-stressed rice seedlings. Front. Plant Sci. 2017, 8, 785. [Google Scholar] [CrossRef] [PubMed]
- Kamiab, F. Exogenous melatonin mitigates the salinity damages and improves the growth of pistachio under salinity stress. J. Plant Nutr. 2020, 43, 1468–1484. [Google Scholar] [CrossRef]
- Imran, M.; Khan, A.L.; Shahzad, R.; Khan, M.A.; Bilal, S.; Khan, A.; Kang, S.-M.; Lee, I.-J. Exogenous melatonin induces drought stress tolerance by promoting plant growth and antioxidant defence system of soybean plants. AoB Plants 2021, 13, plab026. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Li, Q.-T.; Chu, Y.-N.; Reiter, R.J.; Yu, X.-M.; Zhu, D.-H.; Zhang, W.-K.; Ma, B.; Lin, Q.; Zhang, J.-S.; et al. Melatonin enhances plant growth and abiotic stress tolerance in soybean plants. J. Exp. Bot. 2015, 66, 695–707. [Google Scholar] [CrossRef] [Green Version]
- El-Awadi, M.; Sadak, M.; Dawood, M.; Khater, M.; Elashtokhy, M. Amelioration the adverse effects of salinity stress by using γ-radiation in faba bean plants. Bull. NRC 2017, 41, 293–310. [Google Scholar]
- Ahmad, S.; Cui, W.; Kamran, M.; Ahmad, I.; Meng, X.; Wu, X.; Su, W.; Javed, T.; El-Serehy, H.A.; Jia, Z.; et al. Exogenous application of melatonin induces tolerance to salt stress by improving the photosynthetic efficiency and antioxidant defense system of maize seedling. J. Plant Growth Regul. 2021, 40, 1270–1283. [Google Scholar] [CrossRef]
- Sadak, M.S.; Ramadan, A.A.E.-M. Impact of melatonin and tryptophan on water stress tolerance in white lupine (Lupinus termis L.). Physiol. Mol. Biol. Plants 2021, 27, 469–481. [Google Scholar] [CrossRef]
- Hernández-Ruiz, J.; Cano, A.; Arnao, M.B. Melatonin: A growth-stimulating compound present in lupin tissues. Planta 2004, 220, 140–144. [Google Scholar] [CrossRef]
- Weeda, S.; Na Zhang, N.; Zhao, X.; Ndip, G.; Guo, Y.; Buck, G.A.; Fu, C.; Ren, S. Arabidopsis transcriptome analysis reveals key roles of melatonin in plant defense systems. PLoS ONE 2014, 9, e93462. [Google Scholar] [CrossRef] [Green Version]
- Sharp, R.; Else, M.; Cameron, R.; Davies, W. Water deficits promote flowering in Rhododendron via regulation of pre and post initiation development. Sci. Hortic. 2009, 120, 511–517. [Google Scholar] [CrossRef]
- Wada, K.C.; Takeno, K. Stress-induced flowering. Plant Signal. Behav. 2010, 5, 944–947. [Google Scholar] [CrossRef] [PubMed]
- Chimonidou-Pavlidou, D. Effect of irrigation and shading at the stage of flower bud appearance. Acta Hortic. 2001, 547, 245–251. [Google Scholar] [CrossRef]
- Shi, H.; Wei, Y.; Wang, Q.; Reiter, R.J.; He, C. Melatonin mediates the stabilization of DELLA proteins to repress the floral transition in Arabidopsis. J. Pineal Res. 2016, 60, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Hu, Q.; Liu, Y.; Cheng, P.; Cheng, H.; Liu, W.; Xing, X.; Guan, Z.; Fang, W.; Chen, S.; et al. Strigolactone represses the synthesis of melatonin, thereby inducing floral transition in Arabidopsis thaliana in an FLC-dependent manner. J. Pineal Res. 2019, 67, e12582. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Melatonin in flowering, fruit set and fruit ripening. Plant Reprod. 2020, 33, 77–87. [Google Scholar] [CrossRef]
- Arnao, M.B.; Hernández-Ruiz, J. Protective effect of melatonin against chlorophyll degradation during the senescence of barley leaves. J. Pineal Res. 2008, 46, 58–63. [Google Scholar] [CrossRef]
- Din, J.; Khan, S.U.; Ali, I.; Gurmani, A.R. Physiological and agronomic response of canola varieties to drought stress. J. Anim. Plant Sci. 2011, 21, 78–82. [Google Scholar]
- Pandey, H.; Baig, M.; Bhatt, R. Effect of moisture stress on chlorophyll accumulation and nitrate reductase activity at vegetative and flowering stage in Avena species. Agric. Sci. Res. J. 2012, 2, 111–118. [Google Scholar]
- Emiliani, J.; D’Andrea, L.; Ferreyra, M.L.F.; Maulión, E.; Rodríguez, E.J.; Rodriguez-Concepción, M.; Casati, P. A role for β,β-xanthophylls in Arabidopsis UV-B photoprotection. J. Exp. Bot. 2018, 69, 4921–4933. [Google Scholar] [CrossRef]
- Luis Castanares, J.; Alberto Bouzo, C. Effect of Exogenous Melatonin on Seed Germination and Seedling Growth in Melon (Cucumis melo L.) Under Salt Stress. Hortic. Plant J. 2019, 5, 79–87. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, X.; Chen, J.; Chen, A.; Wang, L.; Guo, X.; Niu, Y.; Liu, S.; Mi, G.; Gao, Q. Reducing basal nitrogen rate to improve maize seedling growth, water and nitrogen use efficiencies under drought stress by optimizing root morphology and distribution. Agric. Water Manag. 2018, 212, 328–337. [Google Scholar] [CrossRef]
- Soltys-Kalina, D.; Plich, J.; Strzelczyk-Żyta, D.; Śliwka, J.; Marczewski, W. The effect of drought stress on the leaf relative water content and tuber yield of a half-sib family of ‘Katahdin’-derived potato cultivars. Breed. Sci. 2016, 66, 328–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keyvan, S. The effects of drought stress on yield, relative water content, proline, soluble carbohydrates and chlorophyll of bread wheat cultivars. J. Anim. Plant Sci. 2010, 8, 1051–1060. [Google Scholar]
- Hu, W.; Zhang, J.; Wu, Z.; Loka, D.A.; Zhao, W.; Chen, B.; Wang, Y.; Meng, Y.; Zhou, Z.; Gao, L. Effects of single and combined exogenous application of abscisic acid and melatonin on cotton carbohydrate metabolism and yield under drought stress. Ind. Crop. Prod. 2021, 176, 114302. [Google Scholar] [CrossRef]
- Liu, J.; Wang, W.; Wang, L.; Sun, Y. Exogenous melatonin improves seedling health index and drought tolerance in tomato. Plant Growth Regul. 2015, 77, 317–326. [Google Scholar] [CrossRef]
- Seki, M.; Umezawa, T.; Urano, K.; Shinozaki, K. Regulatory metabolic networks in drought stress responses. Curr. Opin. Plant Biol. 2007, 10, 296–302. [Google Scholar] [CrossRef]
- Chegah, S.; Chehrazi, M.; Albaji, M. Effects of drought stress on growth and development frankinia plant (Frankinia leavis). Bulg. J. Agric. Sci. 2013, 19, 659–665. [Google Scholar]
- Zhang, P.; Liu, L.; Wang, X.; Wang, Z.; Zhang, H.; Chen, J.; Liu, X.; Wang, Y.; Li, C. Beneficial effects of exogenous melatonin on overcoming salt stress in sugar beets (Beta vulgaris L.). Plants 2021, 10, 886. [Google Scholar] [CrossRef]
- Jiang, D.; Lu, B.; Liu, L.; Duan, W.; Meng, Y.; Li, J.; Zhang, K.; Sun, H.; Zhang, Y.; Dong, H.; et al. Exogenous melatonin improves the salt tolerance of cotton by removing active oxygen and protecting photosynthetic organs. BMC Plant Biol. 2021, 21, 331. [Google Scholar] [CrossRef]
- Varghese, N.; Alyammahi, O.; Nasreddine, S.; Alhassani, A.; Gururani, M.A. Melatonin positively influences the photosynthetic machinery and antioxidant system of Avena sativa during salinity stress. Plants 2019, 8, 610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Chen, Y.; Tan, D.-X.; Reiter, R.J.; Chan, Z.; He, C. Melatonin induces nitric oxide and the potential mechanisms relate to innate immunity against bacterial pathogen infection in Arabidopsis. J. Pineal Res. 2015, 59, 102–108. [Google Scholar] [CrossRef] [PubMed]
- Saneoka, H.; Moghaieb, R.E.; Premachandra, G.S.; Fujita, K. Nitrogen nutrition and water stress effects on cell membrane stability and leaf water relations in Agrostis palustris Huds. Environ. Exp. Bot. 2004, 52, 131–138. [Google Scholar] [CrossRef]
- Parkash, V.; Singh, S. A review on potential plant-based water stress indicators for vegetable crops. Sustainability 2020, 12, 3945. [Google Scholar] [CrossRef]
- Rodriguez, C.; Mayo, J.C.; Sainz, R.M.; Antolin, I.; Herrera, F.; Martin, V.; Reiter, R.J. Regulation of antioxidant enzymes: A significant role for melatonin. J. Pineal Res. 2004, 36, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Ahmad, R.; Ashraf, M.Y.; Ashraf, M.; Waraich, E.A. Sunflower (Helianthus annuus L.) response to drought stress at germination and seedling growth stages. Pak. J. Bot 2009, 41, 647–654. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Substrate Type | Peat Moss |
|---|---|
| pH | 6 |
| EC (cm/mmhos) | 0.4–0.5 |
| Humidity % | 50–65 |
| Organic matter (OM)% | >90 |
| Macroelement (1 kg/m3) | |
| N:P:K | 14:10:18 |
| Microelement (mg/kg) | |
| Zn | 32.45 |
| Cu | 15.6 |
| Cd | 0.42 |
| Pb | 15.28 |
| Mo | 0.10 |
| Ni | 8.75 |
| Cr | 0.70 |
| Hg | <0.01 |
| As | 0.112 |
| Co | 1.11 |
| Cl | <0.2 |
| Na | <0.1 |
| Treatments | Shoot Length (cm) | No. of Leaves | Area/Leaf (cm2) | Fresh Weight (g) | Dry Weight (g) | |
|---|---|---|---|---|---|---|
| Effect of MT (µM) under well-irrigated conditions | 0 | 18.87 ± 0.18 bA | 6.67 ± 0.49 bA | 47.97 ± 0.05 dA | 18.19 ± 0.39 cA | 3.12 ± 0.19 bA |
| 50 | 19.07 ± 0.27 bA | 7.33 ± 0.49 aA | 54.49 ± 0.11 cA | 18.15 ± 0.15 cA | 3.12 ± 0.07 bA | |
| 100 | 21.49 ± 0.35 aA | 7.67 ± 0.49 aA | 55.31 ± 0.08 bA | 18.82 ± 0.26 bA | 3.17 ± 0.14 bA | |
| 200 | 21.55 ± 0.34 aA | 7.73 ± 0.46 aA | 55.60 ± 0.18 aA | 21.73 ± 0.27 aA | 3.12 ± 0.19 aA | |
| Effect of MT (µM) under drought conditions | 0 | 13.97 ± 0.47 cB | 4.60 ± 0.51 cB | 18.15 ± 0.20 dB | 11.92 ± 0.17 dB | 1.47 ± 0.06 dB |
| 50 | 15.75 ± 0.37 bB | 5.20 ± 0.56 bB | 21.61 ± 0.16 cB | 13.99 ± 0.09 cB | 1.63 ± 0.09 cB | |
| 100 | 16.78 ± 0.30 aB | 5.73 ± 0.70 abB | 28.78 ± 0.22 bB | 15.92 ± 0.13 bB | 1.85 ± 0.06 bB | |
| 200 | 17.09 ± 0.39 aB | 5.93 ± 0.46 aB | 29.56 ± 0.17 aB | 17.01 ± 0.10 aB | 2.01 ± 0.12 aB | |
| Traits | DTI (%) |
|---|---|
| Shoot length | 74.02 ± 2.83 ef |
| Leaf number | 69.37 ± 9.35 ef |
| Leaf area | 37.84 ± 0.39 gh |
| Shoot fresh weight | 65.53 ± 1.73 f |
| Shoot dry weight | 47.20 ± 3.48 g |
| Time of flower bud emergence | 79.31 ± 2.30 ed |
| Total chlorophyll content | 32.00 ± 1.02 h |
| Carotenoid content | 49.12 ± 0.58 g |
| Relative water content (RWC) | 92.90 ± 0.45 d |
| Proline content | 207.40 ± 0.30 b |
| Peroxidase activity (POD) | 282.35 ± 10.53 a |
| Electrolyte leakage (EL) | 120.49 ± 0.72 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eisa, E.A.; Honfi, P.; Tilly-Mándy, A.; Gururani, M.A. Exogenous Application of Melatonin Alleviates Drought Stress in Ranunculus asiaticus by Improving Its Morphophysiological and Biochemical Attributes. Horticulturae 2023, 9, 262. https://doi.org/10.3390/horticulturae9020262
Eisa EA, Honfi P, Tilly-Mándy A, Gururani MA. Exogenous Application of Melatonin Alleviates Drought Stress in Ranunculus asiaticus by Improving Its Morphophysiological and Biochemical Attributes. Horticulturae. 2023; 9(2):262. https://doi.org/10.3390/horticulturae9020262
Chicago/Turabian StyleEisa, Eman Abdelhakim, Péter Honfi, Andrea Tilly-Mándy, and Mayank Anand Gururani. 2023. "Exogenous Application of Melatonin Alleviates Drought Stress in Ranunculus asiaticus by Improving Its Morphophysiological and Biochemical Attributes" Horticulturae 9, no. 2: 262. https://doi.org/10.3390/horticulturae9020262