
RNA-Seq Based Transcriptomic Analysis of Bud Sport Skin Color in Grape Berries
Abstract
:1. Introduction
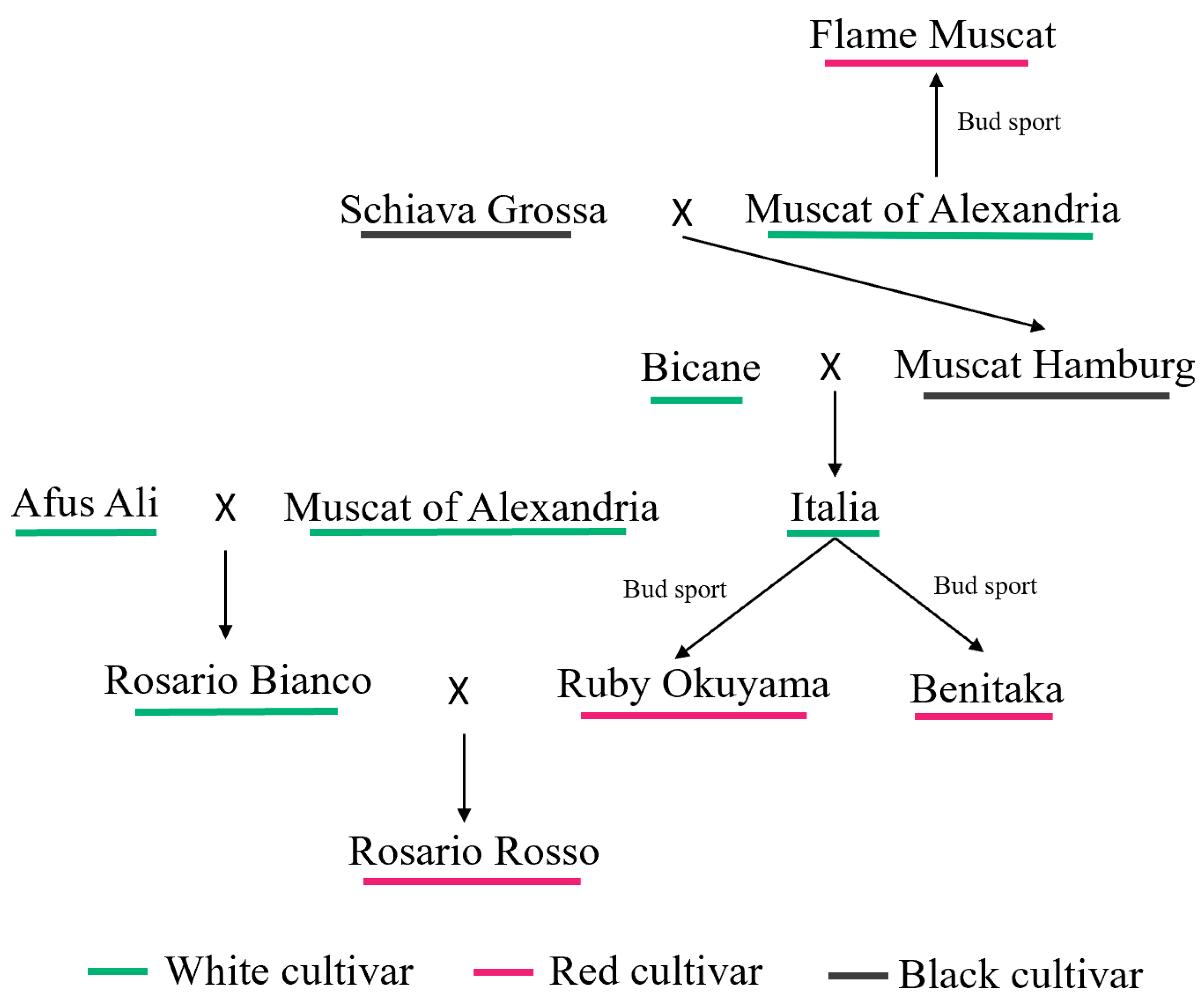
1.1. Bud Sport
1.2. Fruit Color
1.3. Grape Bud Sport
1.4. Transcriptome Sequencing
2. Materials and Methods
2.1. Plant Materials
2.2. RNA Extraction and RNA-Seq
2.3. Transcriptome Sequencing and Analysis
2.4. Statistical Analysis
3. Results
3.1. Quality Control Data Statistics
3.2. Differentially Expressed Gene (DEG) Analysis
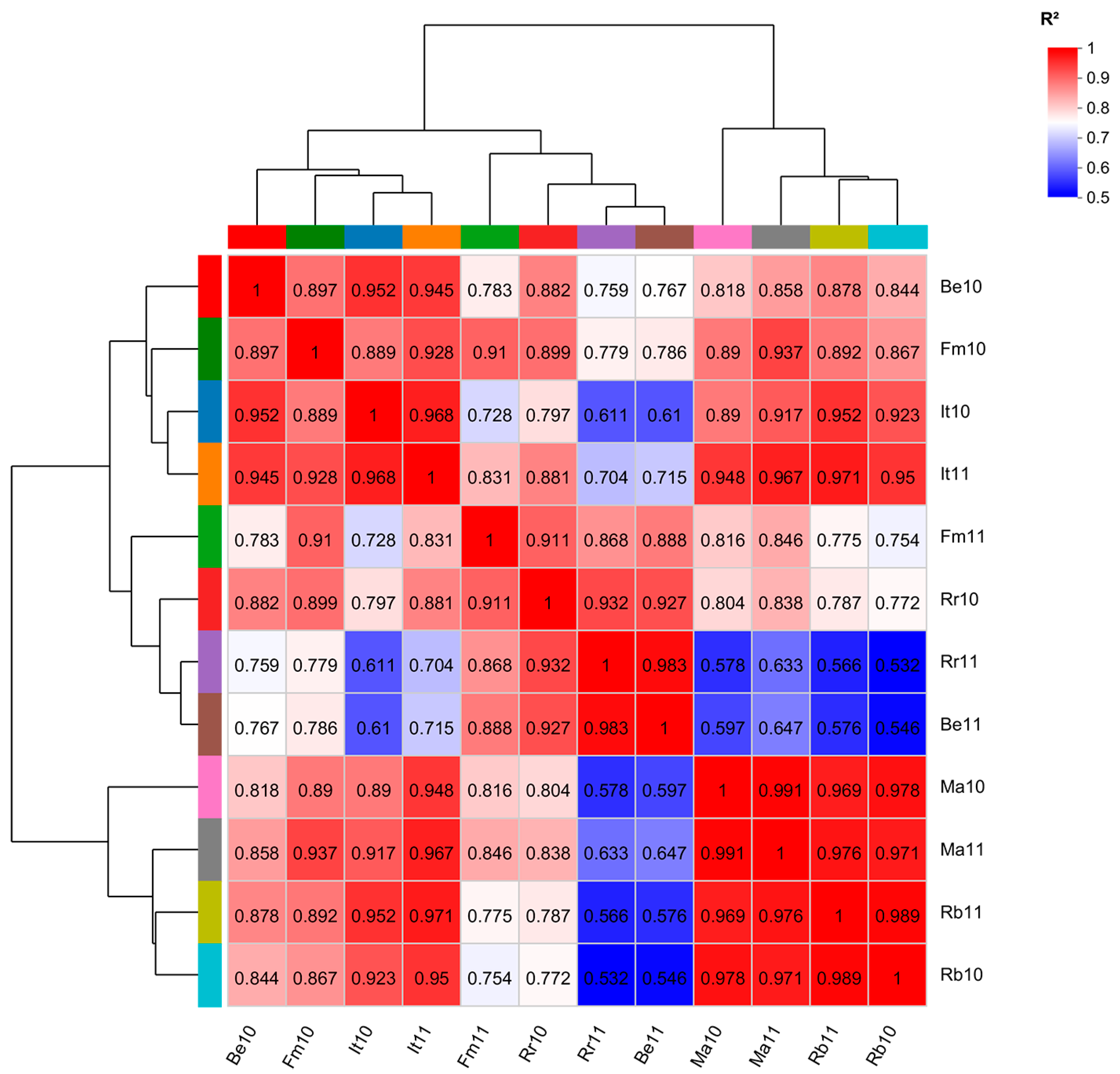
3.3. Correlation Analysis among Each Sample
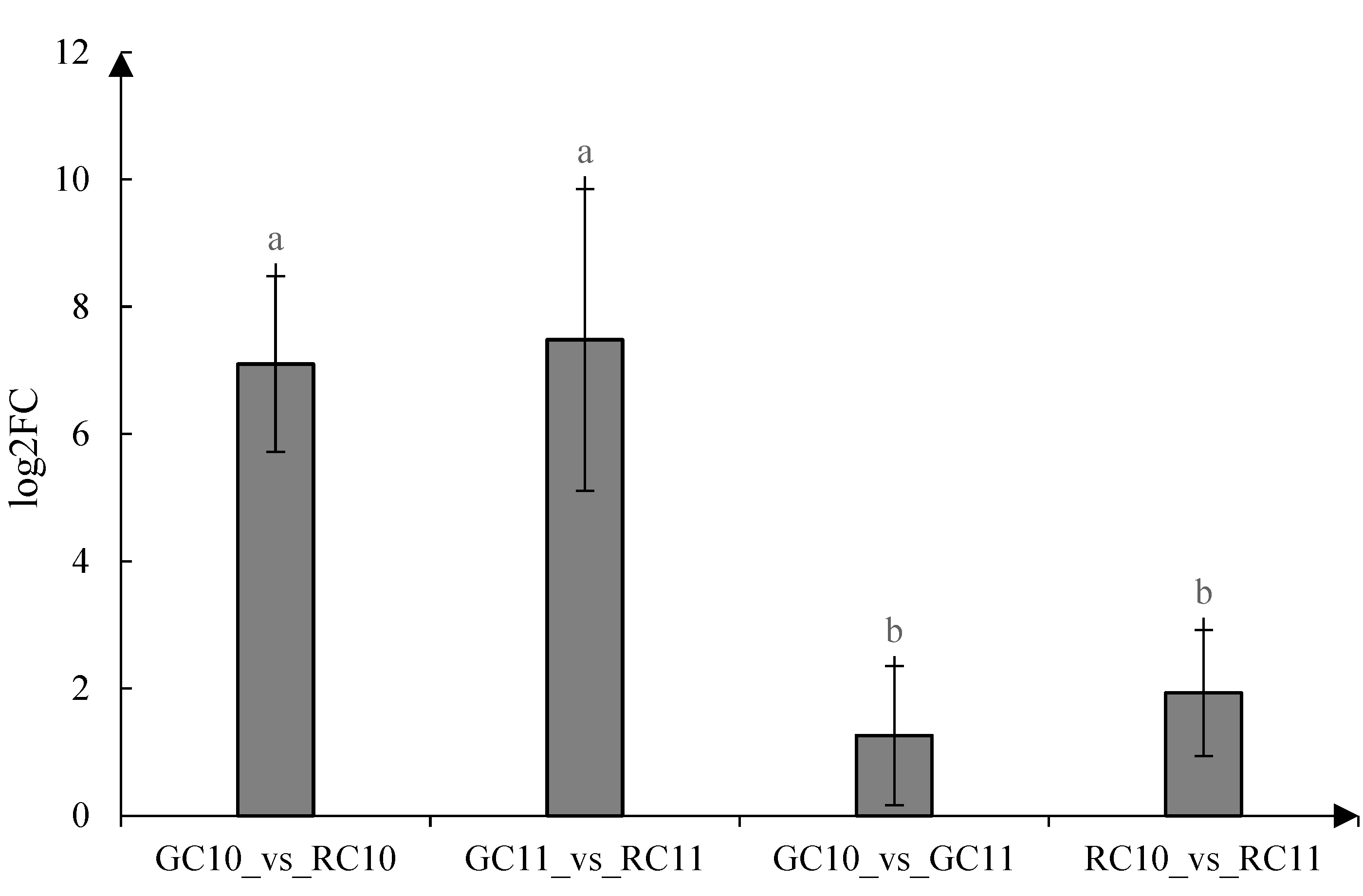
3.4. Gene Expression Level of VvMYBA1 in Berry Skins
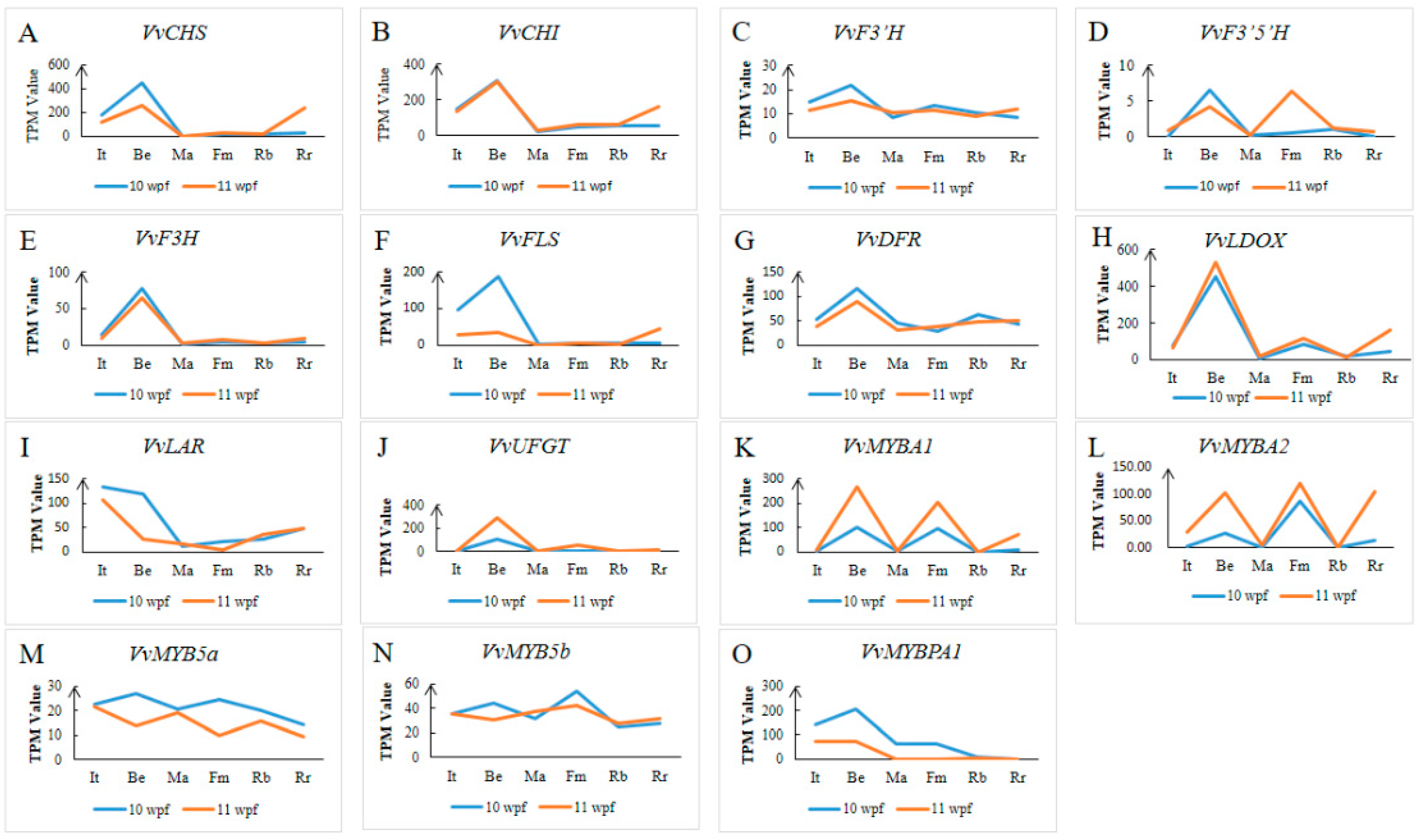
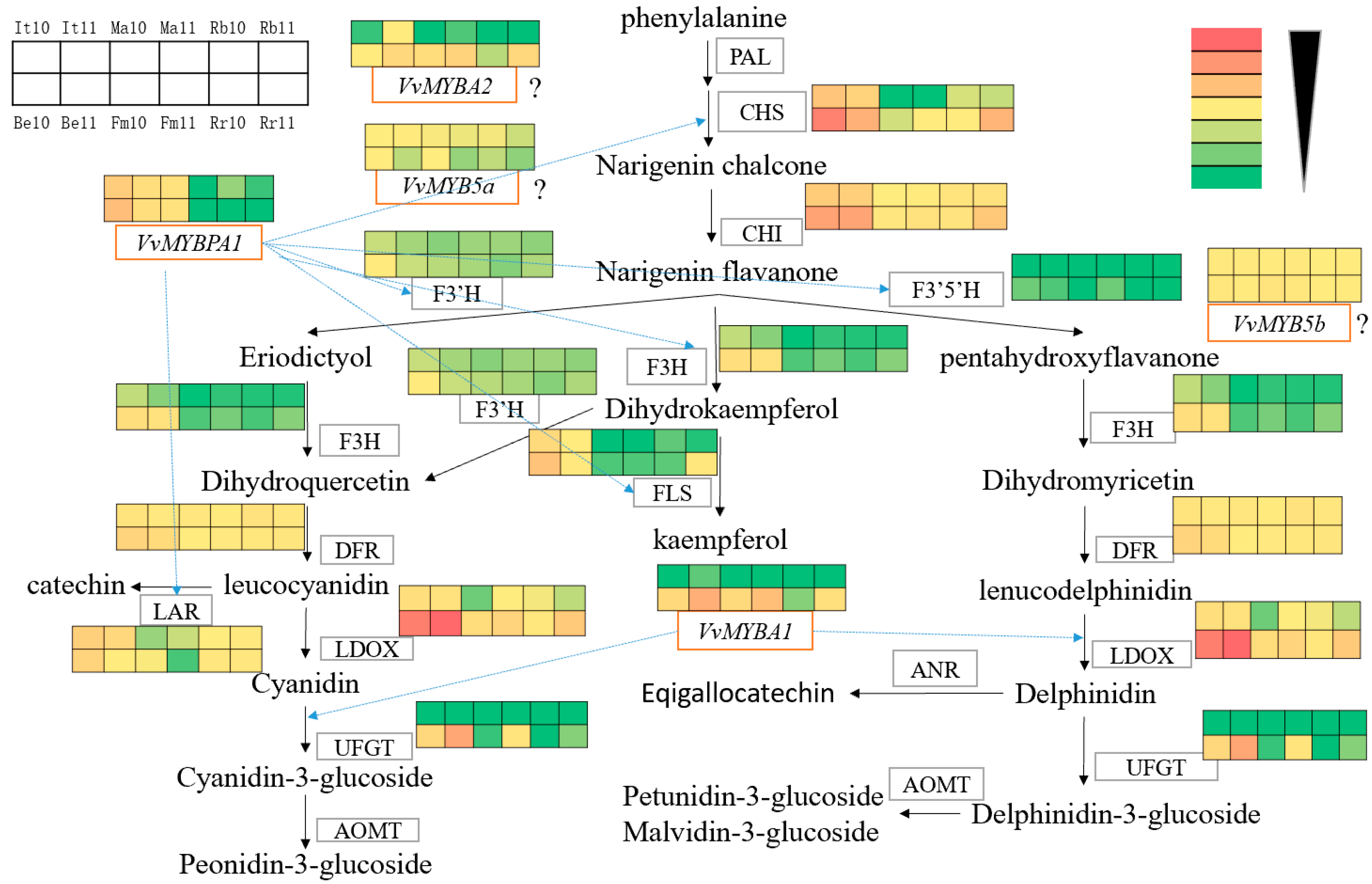
3.5. Anthocyanin-Synthesis-Related Gene Expression Analysis
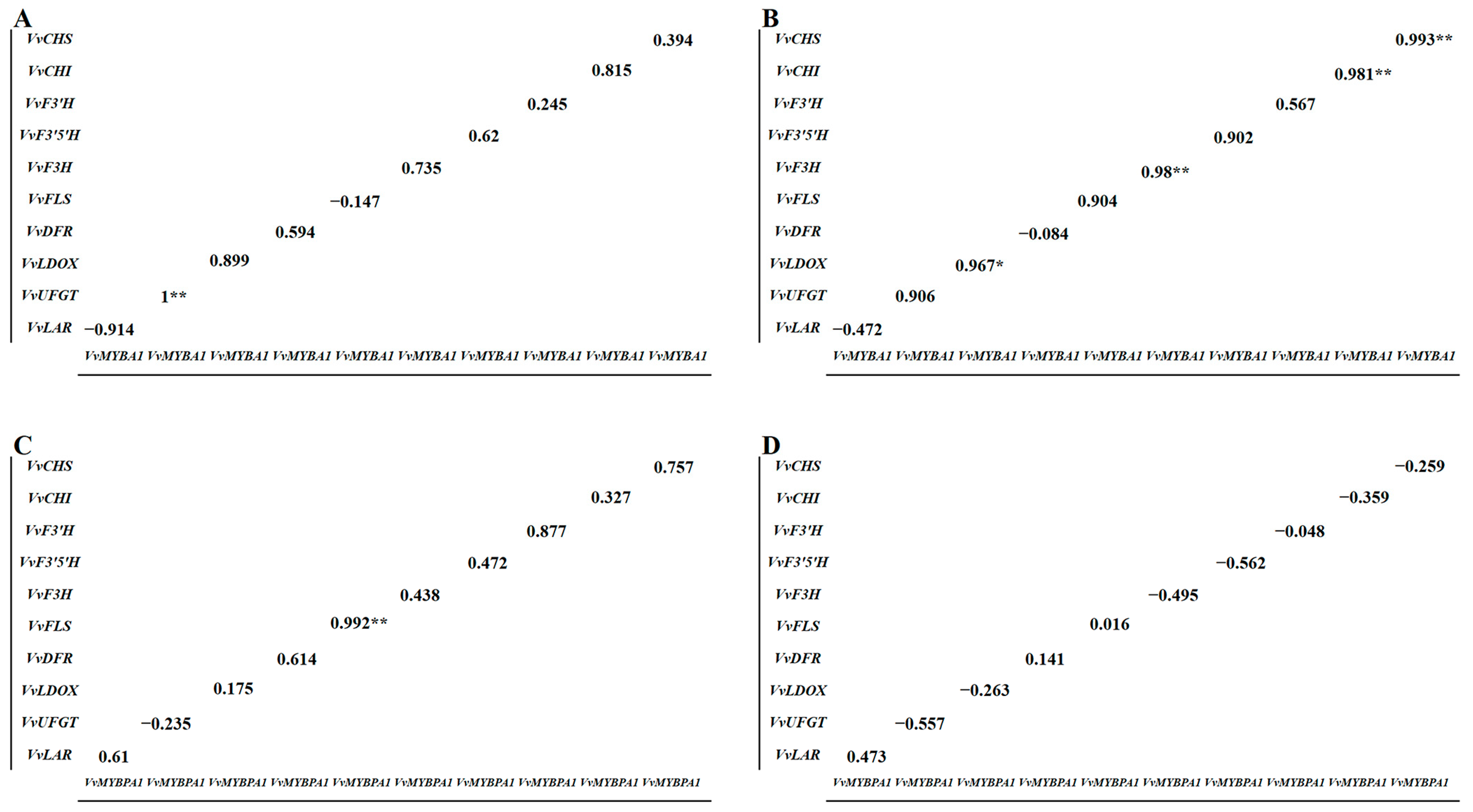
3.6. Correlation Analysis between Anthocyanin-Synthesis-Related Structural Genes and VvMYBA1 and VvMYBPA1 Regulatory Genes among Bud Sport Varieties
3.7. Correlation Analysis of MYB-Related Regulatory Genes and Anthocyanin Synthesis Structure
3.8. Screening of Genes Involved in the Regulation of Metal Ion Binding
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Hansche, P.E.; Beres, W. Genetic remodeling of fruit and nut trees to facilitate cultivar improvement. HortScience 1980, 15, 710–715. [Google Scholar] [CrossRef]
- Massonnet, M.; Cochetel, N.; Minio, A.; Vondras, A.M.; Lin, J.; Muyle, A.; Garcia, J.F.; Zhou, Y.; Delledonne, M.; Riaz, S.; et al. The genetic basis of sex determination in grapes. Nat. Commun. 2020, 11, 2902. [Google Scholar] [CrossRef] [PubMed]
- Foster, T.M.; Aranzana, M.J. Attention sports fans! The far-reaching contributions of bud sport mutants to horticulture and plant biology. Hortic. Res. 2018, 5, 44. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Li, N.; Deng, Z.; Luo, F.; Duan, Y. Selection and evaluation of a thornless and HLB-tolerant bud-sport of pummelo citrus with an emphasis on molecular mechanisms. Front. Plant Sci. 2021, 12, 739108. [Google Scholar] [CrossRef]
- Du, X.; Wang, Y.; Liu, M.; Liu, X.; Jiang, Z.; Zhao, L.; Tang, Y.; Sun, Y. The assessment of epigenetic diversity, differentiation, and structure in the ‘Fuji’ mutation line implicates roles of epigenetic modification in the occurrence of different mutant groups as well as spontaneous mutants. PLoS ONE 2020, 15, e0235073. [Google Scholar] [CrossRef]
- Azuma, A.; Kobayashi, S.; Goto-Yamamoto, N.; Shiraishi, M.; Mitani, N.; Yakushji, H.; Koshita, Y. Color recovery in berries of grape (Vitis vinifera L.) ‘Benitaka’, a bud sport of ‘Italia’, is caused by a novel allele at the VvmybA1 locus. Plant Sci. 2009, 176, 470–478. [Google Scholar] [CrossRef]
- Szymkowiak, E.J.; Sussex, I.M. What chimeras can tell us about plant development. Annu. Rev. Plant Biol. 1996, 47, 351–376. [Google Scholar] [CrossRef]
- Stewart, R.N.; Meyer, F.G.; Dermen, H. Camellia ‘Daisy Eagleson’ a graft chimera of Camellia sasanqua and C. japonica. Am. J. Bot. 1972, 59, 515–524. [Google Scholar] [CrossRef]
- Darwin, C. The Variation of Animals and Plants under Domestication (Cambridge Library Collection—Darwin, Evolution and Genetics, pp. I–Ii); Cambridge University Press: Cambridge, UK, 2010. [Google Scholar] [CrossRef]
- Franks, T.; Botta, R.; Thomas, M.R.; Franks, J. Chimerism in grapevines: Implications for cultivar identity, ancestry and genetic improvement. Theor. Appl. Genet. 2002, 104, 192–199. [Google Scholar] [CrossRef]
- Liu, G.; Li, B.; Hu, P.; Zhou, G. Analysis of differences in mineral element content of early maturation of navel orange and its parent seedlings. Trop. Agric. Sci. 2017, 37, 21–25. [Google Scholar]
- Carolina, R.; Rafael, T.P.; Nuria, M.; Nieves, D.; José, A.V.; Cécile, M.; Thierry, L.; Javier, I.; Manuel, T.; Juan, V.; et al. The major origin of seedless grapes is associated with a 4 missense mutation in the MADS-box gene VviAGL11. Plant Physiol. 2018, 177, 1234–1253. [Google Scholar]
- Tetali, S.; Karkamkar, S.P.; Phalake, S.V. Mutation breeding for inducing seedlessness in grape variety ARI 516. Int. J. Minor Fruits Med. Aromat. Plants 2020, 6, 67–71. [Google Scholar]
- Visser, T.; Verhaegh, J.J.; De Vries, D.P. Pre-selection of compact mutants induced by X-ray treatment in apple and pear. Euphytica 1971, 20, 195–207. [Google Scholar] [CrossRef]
- Kuksova, V.B.; Piven, N.M.; Gleba, Y.Y. Somaclonal variation and in vitro induced mutagenesis in grapevine. Plant Cell Tissue Organ Cult. 1997, 49, 17–27. [Google Scholar] [CrossRef]
- Hamill, S.D.; Smith, M.K.; Dodd, W.A. In vitro induction of banana autotetraploids by colchicine treatment of micropropagated diploids. Aust. J. Bot. 1992, 40, 887–896. [Google Scholar] [CrossRef]
- Predieri, S.; Gatti, E. Effects of gamma radiation on microcuttings of plum (Prunus salicina Lindl.)‘Shiro’. Adv. Hortic. Sci. 2000, 14, 7–11. [Google Scholar]
- Hashmi, G.P.; Hammerschlag, F.A.; Huettel, R.N. Growth, development, and response of peach somaclones to the root-knot nematode, Meloidogyne incognita. J. Am. Soc. Hortic. Sci. 1995, 120, 932–937. [Google Scholar] [CrossRef]
- Wu, J.H.; Ferguson, A.R.; Murray, B.G. Manipulation of ploidy for kiwifruit breeding: In vitro chromosome doubling in diploid Actinidia chinensis Planch. Plant Cell Tissue Organ Cult. 2011, 106, 503–511. [Google Scholar] [CrossRef]
- Takahashi, H. Breeding of strawberry resistant to Alternaria black spot of strawberry varieties (Alternaria althernata strawberry pathotype). Bul. Akita Pref. Coll. Agri. 1993, 19, 1–44. [Google Scholar]
- Ge, H.; Li, Y.; Fu, H.; Fu, H.; Luo, L.; Li, R.; Deng, Z. Production of sweet orange somaclones tolerant to citrus canker disease by in vitro mutagenesis with EMS. Plant Cell Tissue Organ Cult. 2015, 123, 29–38. [Google Scholar] [CrossRef]
- Shamel, A.D.; Pomeroy, C.S. Bud mutations in horticultural crops. J. Hered. 1936, 27, 487–494. [Google Scholar] [CrossRef]
- Lamo, K.; Bhat, D.J.; Kour, K.; Pratap, S. Mutation studies in fruit crops: A review. Int. J. Curr. Microbiol. Appl. Sci. 2017, 6, 3620–3633. [Google Scholar] [CrossRef]
- Allan, A.C.; Hellens, R.P.; Laing, W.A. MYB transcription factors that colour our fruit. Trends Plant Sci. 2008, 13, 99–102. [Google Scholar] [CrossRef] [PubMed]
- Winkel-Shirley, B. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology, and biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar] [CrossRef] [PubMed]
- Grotewold, E. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol. 2006, 57, 761–780. [Google Scholar] [CrossRef]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef]
- Tohge, T.; de Souza, L.P.; Fernie, A.R. Current understanding of the pathways of flavonoid biosynthesis in model and crop plants. J. Exp. Bot. 2017, 68, 4013–4028. [Google Scholar] [CrossRef]
- Jun, J.H.; Xiao, X.; Rao, X.; Dixon, R. Proanthocyanidin subunit composition determined by functionally diverged dioxygenases. Nat. Plants 2018, 4, 1034–1043. [Google Scholar] [CrossRef]
- Gu, C.; Liao, L.; Zhou, H.; Wang, L.; Deng, X.; Han, Y. Constitutive activation of an anthocyanin regulatory gene PcMYB10. 6 is related to red coloration in purple-foliage plum. PLoS ONE 2015, 10, e0135159. [Google Scholar] [CrossRef]
- Sun, B.; Zhu, Z.; Cao, P.; Chen, H.; Chen, C.; Zhou, X.; Mao, Y.; Lei, J. Purple foliage coloration in tea (Camellia sinensis L.) arises from activation of the R2R3-MYB transcription factor CsAN1. Sci. Rep. 2016, 6, srep32534. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, H.; Lin-Wang, K.; Vimolmangkang, S.; Espley, R.; Wang, L.; Allan, A. Transcriptome analysis and transient transformation suggest an ancient duplicated MYB transcription factor as a candidate gene for leaf red coloration in peach. BMC Plant Biol. 2014, 14, 388. [Google Scholar] [CrossRef] [PubMed]
- Matus, J.T.; Aquea, F.; Arce-Johnson, P. Analysis of the grape MYB R2R3 subfamily reveals expanded wine quality-related clades and conserved gene structure organization across Vitis and Arabidopsis genomes. BMC Plant Biol. 2008, 8, 83. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Hu, J.; Han, X.; Li, J.; Gao, Y.; Zhang, G.; Tian, Y. A high-quality apple genome assembly reveals the association of a retrotransposon and red fruit colour. Nat. Commun. 2019, 10, 1494. [Google Scholar] [CrossRef] [PubMed]
- Boss, P.K.; Davies, C.; Robinson, S.P. Expression of anthocyanin biosynthesis pathway genes in red and white grapes. Plant Mol. Biol. 1996, 32, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, S.; Goto-Yamamoto, N.; Hirochika, H. Retrotransposon-induced mutations in grape skin color. Science 2004, 304, 982. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.R.; Lee, E.; Bogs, J.; McDavid, D.; Robinson, S.; Thomas, M. White grapes arose through the mutation of two similar and adjacent regulatory genes. Plant J. 2007, 49, 772–785. [Google Scholar] [CrossRef] [PubMed]
- Lijavetzky, D.; Ruiz-García, L.; Cabezas, J.A.; Andres, M.T.; Bravo, G.; Lbanez, A.; Cabello, F. Molecular genetics of berry colour variation in table grape. Mol. Genet. Genom. 2006, 276, 427–435. [Google Scholar] [CrossRef]
- Walker, A.R.; Lee, E.; Robinson, S.P. Two new grape varieties, bud sports of Cabernet Sauvignon bearing pale-coloured berries, are the result of deletion of two regulatory genes of the berry colour locus. Plant Mol. Biol. 2006, 62, 623–635. [Google Scholar] [CrossRef]
- This, P.; Lacombe, T.; Cadle-Davidson, M.; Owens, C.L. Wine grape (Vitis vinifera L.) color associates with allelic variation in the domestication gene VvmybA1. Theor. Appl. Genet. 2007, 114, 723–730. [Google Scholar] [CrossRef]
- Granhall, I. Spontaneous and induced bud mutations in fruit trees. Acta Agric. Scand. 1954, 4, 594–600. [Google Scholar] [CrossRef]
- Alexander, V. Breeding for Ornamentals: Classical and Molecular Approaches; Springer Science & Business Media: Rehovot, Israel, 2002. [Google Scholar]
- Kobayashi, S.; Ishimaru, M.; Hiraoka, K.; Honda, C. Myb-related genes of the Kyoho grape (Vitis labruscana) regulate anthocyanin biosynthesis. Planta 2002, 215, 924–933. [Google Scholar]
- He, P.; Li, L.; Wang, H.; Chang, Y. An RNA-Seq analysis of the peach transcriptome with a focus on genes associated with skin colour. Czech J. Genet. Plant Breed. 2019, 55, 166–169. [Google Scholar] [CrossRef]
- Azuma, A.; Kono, A.; Sato, A. Simple DNA marker system reveals genetic diversity of MYB genotypes that determine skin color in grape genetic resources. Tree Genet. Genomes 2020, 16, 29. [Google Scholar] [CrossRef]
- Bogs, J.; Jaffé, F.W.; Takos, A.M.; Walker, A.R.; Robinson, S.P. The grapevine transcription factor VvMYBPA1 regulates proanthocyanidin synthesis during fruit development. Plant Physiol. 2007, 143, 1347–1361. [Google Scholar] [CrossRef]
- Wang, N.; Qu, C.; Jiang, S.; Chen, Z.; Xu, H.; Fang, H.; Su, M.; Zhang, J.; Wang, Y.; Liu, W.; et al. The proanthocyanidin-specific transcription factor MdMYBPA1 initiates anthocyanin synthesis under low-temperature conditions in red-fleshed apples. Plant J. 2018, 96, 39–55. [Google Scholar] [CrossRef] [PubMed]
- Primetta, A.K.; Karppinen, K.; Riihinen, K.R.; Jaakola, L. Metabolic and molecular analyses of white mutant Vaccinium berries show down-regulation of MYBPA1-type R2R3 MYB regulatory factor. Planta 2015, 242, 631–643. [Google Scholar] [CrossRef] [PubMed]
- Günther, C.S.; Dare, A.P.; McGhie, T.K.; Deng, C.; Lafferty, D.J.; Plunkett, B.J.; Grierson, E.R.; Turner, J.L.; Jaakola, L.; Albert, N.W.; et al. Spatiotemporal modulation of flavonoid metabolism in blueberries. Front. Plant Sci. 2020, 11, 545. [Google Scholar] [CrossRef] [PubMed]
- Karppinen, K.; Lafferty, D.J.; Albert, N.W.; Mikkola, N.; McGhie, T.; Allan, A.C.; Afzal, B.M.; Häggman, H.; Espley, R.V.; Jaakola, L. MYBA and MYBPA transcription factors co-regulate anthocyanin biosynthesis in blue-coloured berries. New Phytol. 2021, 232, 1350–1367. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shi, Z.; Maximova, S.N.; Payne, M.J.; Guiltinan, M.J. Tc-MYBPA is an Arabidopsis TT2-like transcription factor and functions in the regulation of proanthocyanidin synthesis in Theobroma cacao. BMC Plant Biol. 2015, 15, 160. [Google Scholar] [CrossRef]
- Terrier, N.; Torregrosa, L.; Ageorges, A.; Vialet, S.; Verriès, C.; Cheynier, V.; Romieu, C. Ectopic expression of VvMybPA2 promotes proanthocyanidin biosynthesis in grapevine and suggests additional targets in the pathway. Plant Physiol. 2009, 149, 1028–1041. [Google Scholar] [CrossRef] [PubMed]
- Boss, P.K.; Davies, C.; Robinson, S.P. Anthocyanin composition and anthocyanin pathway gene expression in grapevine sports differing in berry skin colour. Aust. J. Grape Wine Res. 1996, 2, 163–170. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample Name | Raw Reads | Clean Reads | Error Rate (%) | Q20 (%) | Q30 (%) | GC Content (%) | Total Mapped |
|---|---|---|---|---|---|---|---|
| Be10 | 42,647,782 | 42,347,510 | 0.0245 | 98.26 | 94.66 | 46.27 | 34,536,653 (81.56%) |
| Be11 | 45,278,732 | 44,781,572 | 0.0254 | 97.85 | 93.73 | 47.01 | 35,050,465 (78.27%) |
| Fm10 | 45,595,408 | 45,132,012 | 0.0252 | 97.96 | 93.93 | 46.53 | 41,466,312 (91.88%) |
| Fm11 | 42,127,462 | 41,834,580 | 0.0248 | 98.11 | 94.32 | 46.74 | 38,000,195 (90.83%) |
| It10 | 41,804,432 | 41,371,026 | 0.0247 | 98.14 | 94.45 | 46.51 | 38,519,065 (93.11%) |
| It11 | 48,476,130 | 48,148,376 | 0.025 | 98.06 | 94.16 | 45.96 | 44,559,329 (92.55%) |
| Ma10 | 46,362,636 | 46,058,214 | 0.0248 | 98.1 | 94.28 | 46.4 | 41,828,638 (90.82%) |
| Ma11 | 47,352,198 | 46,978,274 | 0.0251 | 97.98 | 94 | 46.36 | 42,739,595 (90.98%) |
| Rb10 | 41,748,704 | 41,378,660 | 0.0252 | 97.95 | 93.95 | 46.35 | 37,569,526 (90.79%) |
| Rb11 | 43,902,394 | 43,576,488 | 0.0249 | 98.06 | 94.18 | 46.04 | 40,136,365 (92.11%) |
| Rr10 | 43,522,364 | 43,177,256 | 0.0249 | 98.06 | 94.22 | 46.48 | 39,821,925 (92.23%) |
| Rr11 | 42,739,842 | 42,458,930 | 0.0246 | 98.19 | 94.51 | 46.57 | 38,767,709 (91.31%) |
| Difference Comparison Group | Total DEG Number | Upregulated DEG Number | Downregulated DEG Number |
|---|---|---|---|
| It10_vs_It11 | 3124 | 1941 | 1183 |
| Be10_vs_Be11 | 2707 | 1114 | 1593 |
| It10_vs_Be10 | 1731 | 1095 | 636 |
| It11_vs_Be11 | 2074 | 925 | 1149 |
| Ma10_vs_Ma11 | 1766 | 1090 | 676 |
| Fm10_vs_Fm11 | 1716 | 505 | 1211 |
| Ma10_vs_Fm10 | 2790 | 1531 | 1259 |
| Ma11_vs_Fm11 | 2000 | 551 | 1449 |
| Rb10_vs_Rb11 | 1640 | 1152 | 488 |
| Rr10_vs_Rr11 | 1579 | 865 | 714 |
| Rb10_vs_Rr10 | 2962 | 931 | 2031 |
| Rb11_vs_Rr11 | 3282 | 936 | 2346 |
| Gene | VvMYB5a | VvMYB5b | VvMYBPA1 | VvMYBA1 | VvMYBA2 |
|---|---|---|---|---|---|
| VvCHS | 0.194 | 0.154 | 0.731 ** | 0.384 | 0.231 |
| VvCHI | −0.109 | 0.075 | 0.665 | 0.544 | 0.306 |
| VvF3’H | 0.459 | 0.487 | 0.839 ** | 0.431 | 0.22 |
| VvF3’5’H | −0.09 | 0.33 | 0.349 | 0.747 ** | 0.481 |
| VvF3H | 0.24 | 0.184 | 0.709 ** | 0.585 | 0.222 |
| VvFLS | 0.468 | 0.263 | 0.873 ** | 0.121 | −0.059 |
| VvDFR | 0.231 | -0.09 | 0.663 | 0.374 | 0.007 |
| VvLDOX | 0.029 | 0.187 | 0.548 | 0.756 ** | 0.459 |
| VvUFGT | −0.152 | 0.007 | 0.304 | 0.831 ** | 0.468 |
| VvLAR | 0.5 | 0.078 | 0.752 ** | −0.246 | −0.295 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, W.; Fang, H.; Yue, L.; Khalil-Ur-Rehman, M.; Huang, Y.; Du, Z.; Yang, G.; Xu, Y. RNA-Seq Based Transcriptomic Analysis of Bud Sport Skin Color in Grape Berries. Horticulturae 2023, 9, 260. https://doi.org/10.3390/horticulturae9020260
Wen W, Fang H, Yue L, Khalil-Ur-Rehman M, Huang Y, Du Z, Yang G, Xu Y. RNA-Seq Based Transcriptomic Analysis of Bud Sport Skin Color in Grape Berries. Horticulturae. 2023; 9(2):260. https://doi.org/10.3390/horticulturae9020260
Chicago/Turabian StyleWen, Wuwu, Haimeng Fang, Lingqi Yue, Muhammad Khalil-Ur-Rehman, Yiqi Huang, Zhaoxuan Du, Guoshun Yang, and Yanshuai Xu. 2023. "RNA-Seq Based Transcriptomic Analysis of Bud Sport Skin Color in Grape Berries" Horticulturae 9, no. 2: 260. https://doi.org/10.3390/horticulturae9020260