
Activity of Essential Oils and Plant Extracts as Biofungicides for Suppression of Soil-Borne Fungi Associated with Root Rot and Wilt of Marigold (Calendula officinalis L.)

 ,
,  ,
,  ,
,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Isolation and Frequency of Fungi Associated with Root Rot and Wilt of Marigold
2.2. Pathogenicity Test of Root Rot and Wilt Fungi towards Potted Marigold Seedlings
2.3. Antifungal Activity of Essential Oils against the Growth of Pathogenic Fungi in an In Vitro Assay
2.4. Antifungal Activity of Plant Extracts against the Growth of Pathogenic Fungi in an In Vitro Assay
2.5. Efficiency of Essential Oils, Plant Extracts, and Fungicides in Reducing Root Rot and Wilt Diseases of Potted Marigold in Artificially Infested Soil
2.6. Management of Marigold Root Rot and Wilt Diseases under Open Field Conditions
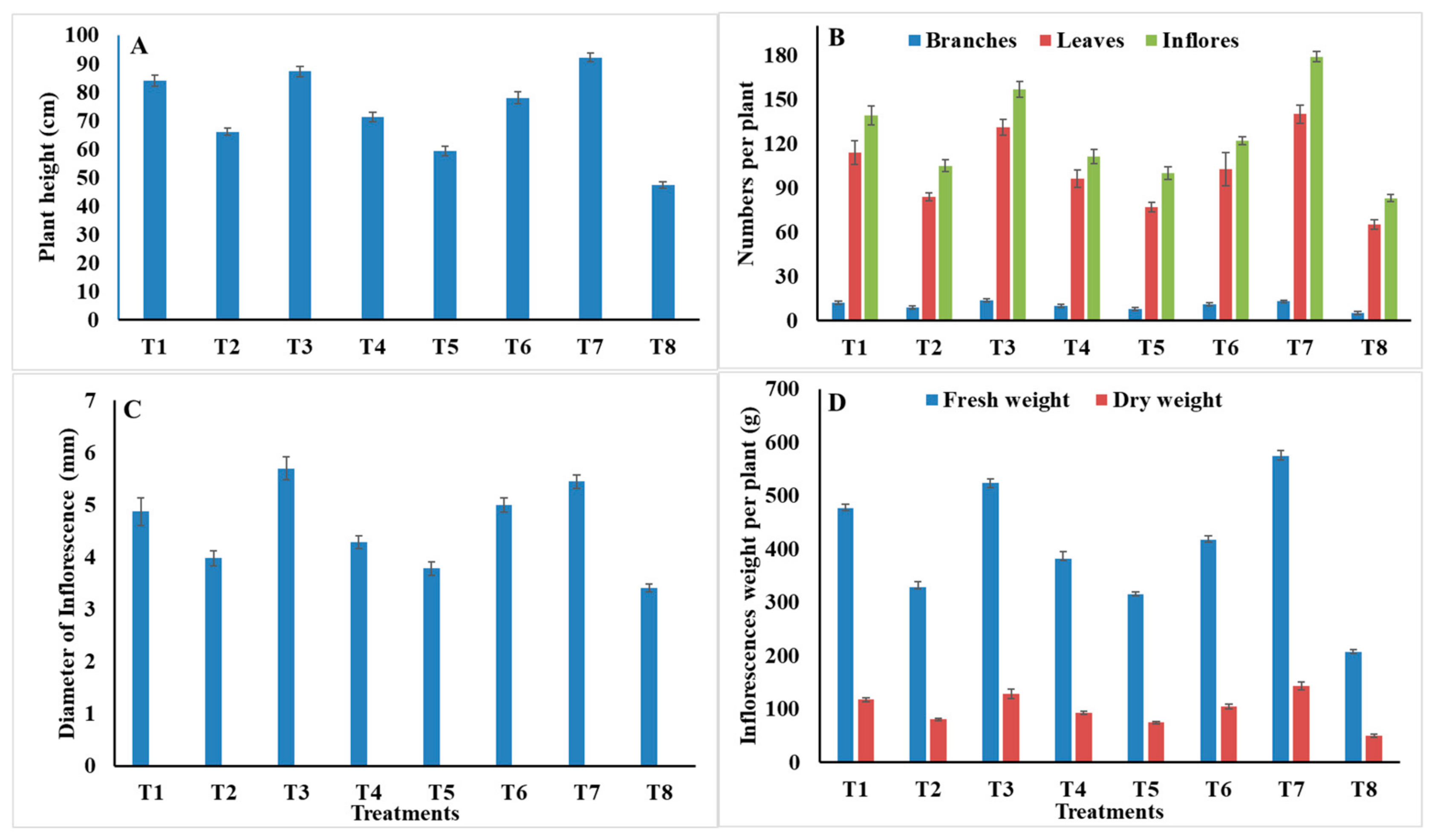
2.7. Morphological and Yield Characteristics
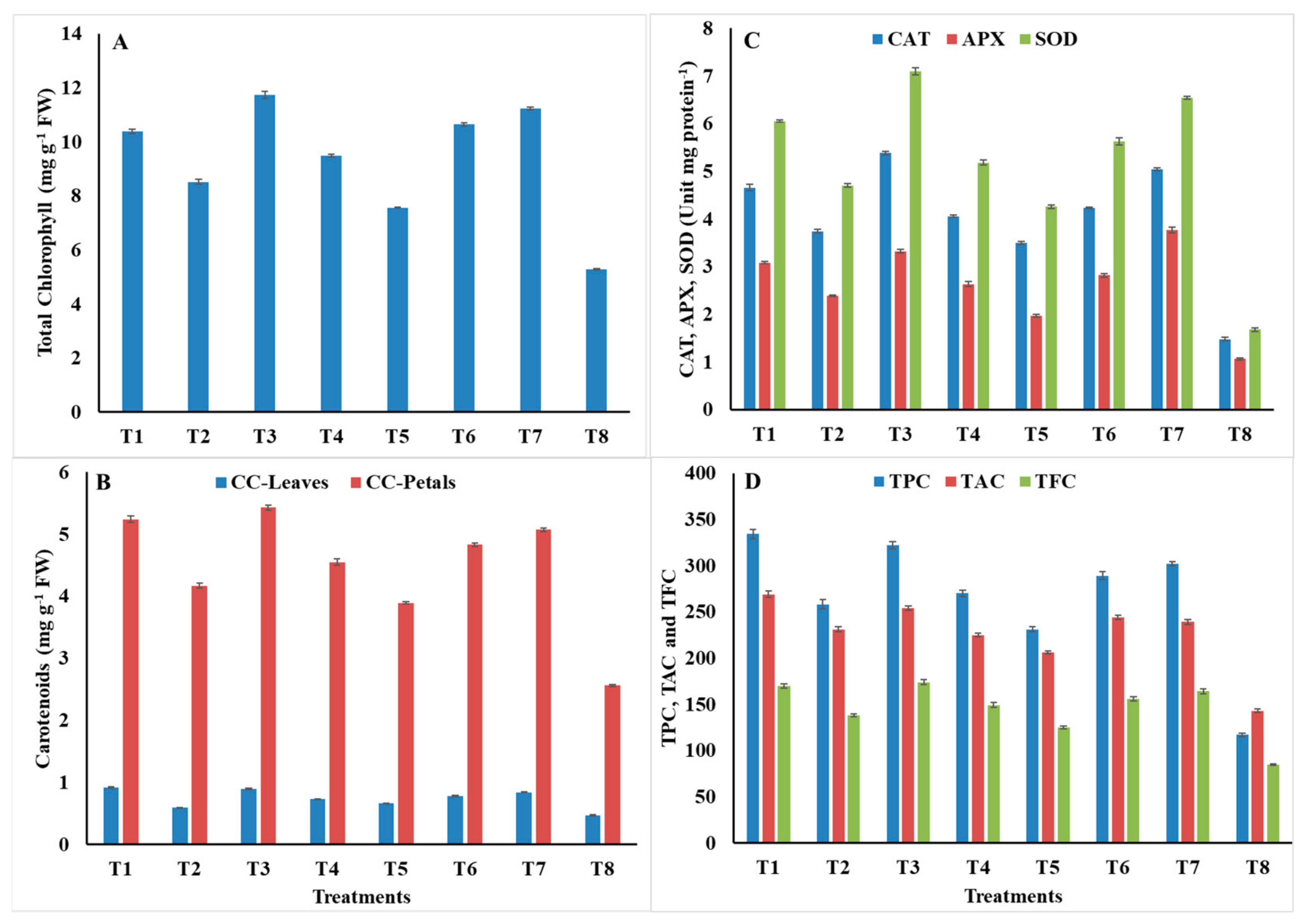
2.8. Measurement of Phytochemical Parameters
2.9. Activity of Defense-Related Antioxidant Enzymes
3. Discussion
4. Materials and Methods
4.1. Plant Material, Treatment Details, and Experimental Site Conditions
4.2. Extraction of Essential Oils and Plant Extracts
4.2.1. Plant Samples Collection, Drying, and Grinding
4.2.2. Essential Oils Extraction and Used Concentrations
4.2.3. Preparation of Methanol Plant Extracts and Required Concentrations
4.3. In Vitro Assays
4.3.1. Culture Medium, Plant Sampling, and Pathogen Isolation
4.3.2. Evaluation of the Antifungal Activity of Essential Oils and Plant Extracts against the Growth of Pathogenic Fungi
4.4. Greenhouse Evaluations
4.4.1. Soil Infestation and Pathogenicity Test
4.4.2. Pot Experimental Layout and Disease Control Approach
4.5. Field Trial Design and Management Strategy
4.5.1. In Vivo Measurements
Disease Assessments
- A.
- The incidence of damping-off at pre- and post-emergence stages was recorded 15 and 35 days after sowing, respectively, according to the following equations;
- B.
- The disease incidence of root rot/wilt was assessed 90 days after sowing according to the equation proposed by Chavdarov [107]:
- C.
- The disease incidence data was re-used to calculate the percentage disease reduction for each treatment as the following equation:
- D.
- The survived plants at the end of the experiment were calculated as the ratio of infected plants to the total number of plants as in the following equation:
Morphological Parameters
4.6. Biochemical Assays
4.6.1. Phytochemical Parameters
Preparation of Plant Material Extract
Total Phenolic Compounds (TPC)
Total Flavonoid Content (TFC)
Total Anthocyanin Content (TAC)
4.6.2. Chlorophyll and Carotenoids Content
4.6.3. Antioxidant Enzyme Activities
Enzyme Extraction
Catalase (CAT) Activity
Ascorbate Peroxidase (APX) Activity
Superoxide Dismutase (SOD) Activity
4.7. Statistical Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Gazim, Z.C.; Rezende, C.M.; Fraga, S.R.; Svidzinski, T.I.E.; Cortez, D.A.G. Antifungal activity of the essential oil from Calendula officinalis L. (Asteraceae) growing in Brazil. Braz. J. Microbiol. 2008, 39, 613. [Google Scholar] [CrossRef]
- Golestani, M.; Dolatkhahi, A.; Kazemi, F. Effect of planting dates on flowering period of Calendula officinalis, Bellis perennis and Viola sp. Adv. Crop Sci. 2013, 3, 563–567. [Google Scholar]
- Verma, P.K.; Raina, R.; Agarwal, S.; Kour, H. Phytochemical ingredients and pharmacological potential of Calendula officinalis Linn. Pharm. Biomed. Res. 2018, 4, 1–17. [Google Scholar] [CrossRef]
- Gunasekaran, S.; Nayagam, A.A.J.; Natarajan, R. Wound healing potentials of herbal ointment containing Calendula officinalis Linn. on the alteration of immunological markers and biochemical parameters in excision wounded animals. Clin. Phytosci. 2020, 77, 2–8. [Google Scholar] [CrossRef]
- Escher, G.B.; Borges, L.D.C.; Santos, J.S.; Cruz, T.M.; Marques, M.P.; do Carmo, M.A.V.; Azevedo, L.; Furtado, M.M.; Sant’Ana, A.S.; Wen, M.; et al. From the field to the pot: Phytochemical and functional analyses of Calendula officinalis L. flower for incorporation in an organic yogurt. Antioxidants 2019, 8, 559. [Google Scholar] [CrossRef]
- Sohi, H.S. Personal communication on disease of marigold; IIHR: Bangalore, India, 1983.
- Heibertshausen, D.; KortekampInfection, A. Cycle and biological control of Podosphaera xanthii, the powdery mildew of marigold (Calendula officinalis). Gesunde Pflanz. 2004, 56, 201–207. [Google Scholar] [CrossRef]
- Garibaldi, A.; Minuto, A.; Gullino, M.L. First report of Sclerotinia sclerotiorum on Calendula officinalis in Italy. Dis. Notes 2007, 85, 446. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, Y.; Kumar, P. Diseases of ornamental plants and their management. In Eco-Friendly Innovative Approaches in Plant Disease Management; Singh, V.K., Singh, Y., Singh, A., Eds.; Int. Book Distributors and Publisher: New Delhi, India, 2012; pp. 543–572. [Google Scholar]
- Abdel-Wahed, G.A. Application of some fungicides alternatives for management root rot and wilt fungal diseases of marigold (Calendula officinalis L.). Sci. J. Agric. Sci. 2020, 2, 31–41. [Google Scholar] [CrossRef]
- Kumar, V. Marigold Diseases and Its Control. Agropedia. 2012. Available online: http://agropedia.iitk.ac.in/content/marigold-diseases-its-control (accessed on 15 June 2022).
- Seleiman, M.F.; Santanen, A.; Mäkelä, P. Recycling sludge on cropland as fertilizer-Advantages and risks. Resour. Conserv. Recycl. 2020, 155, 104647. [Google Scholar] [CrossRef]
- Abdelmoteleb, A.; Moreno-Ramírez, L.; Valdez-Salas, B.; Seleiman, M.F.; El-Hendawy, S.; Aldhuwaib, K.J.; Alotaibi, M.; González-Mendoza, D. New Bacillus subtilis Strains Isolated from Prosopis glandulosa Rhizosphere for Suppressing Fusarium Spp. and Enhancing Growth of Gossypium hirsutum L. Biology 2023, 12, 73. [Google Scholar] [CrossRef]
- Seepe, H.A.; Nxumalo, W.; Amoo, S.O. Natural products from medicinal plants against phytopathogenic fusarium species: Current research endeavors, challenges and prospects. Molecules 2021, 26, 6539. [Google Scholar] [CrossRef]
- Yan, Y.; Liu, Q.; Jacobsen, S.E.; Tang, Y. The impact and prospect of natural product discovery in agriculture. EMBO Rep. 2018, 19, e46824. [Google Scholar] [CrossRef]
- Valdivieso-Ugarte, M.; Gomez-Llorente, C.; Plaza-Díaz, J.; Gil, Á. Antimicrobial, antioxidant, and immunomodulatory properties of essential oils: A systematic review. Nutrients 2019, 11, 2786. [Google Scholar] [CrossRef]
- Sultana, S.; Akhtar, N.; Asif, H.M. Phytochemical screening and antipyretic effects of hydro-methanol extract of Melia azedarach leaves in rabbits. Bangladesh J. Pharmacol. 2013, 8, 214–217. [Google Scholar] [CrossRef]
- Sharma, A.; Rajendran, S.; Srivastava, A.; Sharma, S.; Kundu, B. Antifungal activities of selected essential oils against Fusarium oxysporum f. sp. lycopersici 1322, with emphasis on Syzygium aromaticum essential oil. J. Biosci. Bioeng. 2017, 123, 308–313. [Google Scholar] [CrossRef]
- Císarová, M.; Hleba, L.; Medo, J.; Tančinová, D.; Mašková, Z.; Čuboň, J.; Kováčik, A.; Foltinová, D.; Božik, M.; Klouček, P. The in vitro and in situ effect of selected essential oils in vapor phase against bread spoilage toxicogenic aspergilla. Food Control 2020, 110, 107007. [Google Scholar] [CrossRef]
- Moumni, M.; Allagui, M.B.; Mezrioui, K.; Ben Amara, H.; Romanazzi, G. Evaluation of seven essential oils as seed treatments against seed borne fungal pathogens of Cucurbita maxima. Molecules 2021, 26, 2354. [Google Scholar] [CrossRef]
- Passos, J.L.P.; Barbosa, L.C.A.; Demuner, A.J.; Alvarenga, E.S.; da Silva, C.M.; Barreto, R.W. Chemical characterization of volatile compounds of Lantana camara L. and L. radula Sw. and their antifungal activity. Molecules 2012, 17, 11447–11455. [Google Scholar] [CrossRef]
- Seepe, H.A.; Amoo, S.O.; Nxumalo, W.; Adeleke, R.A. Sustainable use of thirteen south African medicinal plants for the management of crop diseases caused by Fusarium species–An in vitro study. S. Afr. J. Bot. 2020, 130, 456–464. [Google Scholar] [CrossRef]
- Khaledi, N.; Taheri, P.; Tarighi, S. Antifungal activity of various essential oils against Rhizoctonia solani and Macrophomina phaseolina as major bean pathogens. J. Appl. Microbiol. 2015, 118, 704–717. [Google Scholar] [CrossRef]
- Zabka, M.; Pavela, R.; Slezakova, L. Antifungal effect of Pimenta dioica essential oil against dangerous pathogenic and toxinogenic fungi. Ind. Crop Prod. 2009, 30, 250–253. [Google Scholar] [CrossRef]
- Ibáñeza, M.D.; López-Gresab, M.P.; Lisónb, P.; Rodrigob, I.; Bellésb, J.M.; González-Masa, M.C.; Blázqueza, M.A. Essential oils as natural antimicrobial and antioxidant products in the agrifood industry. MOL2NET 2019, 5, 2624–5078. [Google Scholar]
- Mohamed, A.B.; El-Sheikh, A.M.M.; El-Sharkawy, R.M.I. Antifungal activity of bioagents and plant extracts against certain fungal diseases of potatoes. J. Phytopathol. Pest Manag. 2021, 8, 29–45. [Google Scholar]
- Nuzhat, T.; Vidyasagar, G.M. Antifungal investigations on plant essential oils. A review. Int. J. Pharm. Pharm. Sci. 2013, 5, 19–28. [Google Scholar]
- United Nation Industrial Development Organisation FAO. Herbs, Spices and Essential Oils Post-Harvest Operations in Developing Countries; United Nation Industrial Development Organisation: Vienna, Austria, 2005; p. 9.
- Yu, J.; Su, J.; Li, F.; Gao, J.; Li, B.; Pang, M.; Lv, G.; Chen, S. Identification and quantification of pine needle essential oil from different habitats and species of China by GC-MS and GC method. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 1–9. [Google Scholar] [CrossRef]
- Ojaghian, M.R.; Sun, X.; Zhang, L.; Li, X.; Xie, G.L.; Zhang, J.; Wang, L. Effect of E-Cinnamaldehyde against Sclerotinia sclerotiorum on potato and induction of glutathione s-transferase genes. Physiol. Mol. Plant Pathol. 2015, 91, 66–71. [Google Scholar] [CrossRef]
- Císarová, M.; Tančinová, D.; Medo, J.; Kačániová, M. The in vitro effect of selected essential oils on the growth and mycotoxin production of Aspergillus species. J. Environ. Sci. Health Part B 2016, 51, 668–674. [Google Scholar] [CrossRef]
- Yeole, G.J.; Kotkar, H.M.; Teli, N.P.; Mendki, P.S. Herbal fungicide to control fusarium wilt in tomato plants. Biopestic. Int. 2016, 12, 25–35. [Google Scholar]
- Desam, N.R.; Al-Rajab, A.J.; Sharma, M.; Mylabathula, M.M.; Gowkanapalli, R.R.; Albratty, M. Chemical constituents, in vitro antibacterial and antifungal activity of Mentha piperita L. (peppermint) essential oils. J. King Saud Univ. Sci. 2019, 31, 528–533. [Google Scholar] [CrossRef]
- Xing, Y.; Li, X.; Xu, Q.; Yun, J.; Lu, Y. Antifungal activities of cinnamon oil against Rhizopus nigricans, Aspergillus flavus and Penicillium expansum in vitro and in vivo fruit test. Int. J. Food Sci. Technol. 2010, 45, 1837–1842. [Google Scholar] [CrossRef]
- Sempere-Ferre, F.; Asamar, J.; Castell, V.; Roselló, J.; Santamarina, M.P. Evaluating the antifungal potential of botanical compounds to control Botryotinia fuckeliana and Rhizoctonia solani. Molecules 2021, 26, 2472. [Google Scholar] [CrossRef]
- Draz, I.S.; Elkwaga, A.A.; Elzaawely, A.A.; El-Zahaby, H.M.; Ismail, A.W.A. Application of plant extracts as inducers to challenge leaf rust of wheat. Egypt. J. Biol. Pest Cont. 2019, 29, 6. [Google Scholar] [CrossRef]
- Lagrouh, F.; Dakka, N.; Bakri, Y. The antifungal activity of Moroccan plants and the mechanism of action of secondary metabolites from plants. J. Mycol. Méd. 2017, 27, 303–311. [Google Scholar] [CrossRef]
- Tarkowski, Ł.P.; Van De Poel, B.; Höfte, M.; Ende, W.V.D. Sweet immunity: Inulin boosts resistance of lettuce (Lactuca sativa) against grey mold (Botrytis cinerea) in an ethylene-dependent manner. Int. J. Mol. Sci. 2019, 20, 1052. [Google Scholar] [CrossRef]
- De Pinto, M.C.; De Gara, L. Changes in the ascorbate metabolism of apoplastic and symplastic spaces are associated with cell differentiation. J. Exp. Bot. 2004, 55, 2559–2569. [Google Scholar] [CrossRef]
- Walter, S.; Nicholson, P.; Doohan, F. Action and reaction of host and pathogen during Fusarium head blight disease. New Phytol. 2009, 185, 54–66. [Google Scholar] [CrossRef]
- Barna, B.; Fodor, J.; Harrach, B.; Pogany, M.; Király, Z. The Janus face of reactive oxygen species in resistance and susceptibility of plants to necrotrophic and biotrophic pathogens. Plant Physiol. Biochem. 2012, 59, 37–43. [Google Scholar] [CrossRef]
- Hewedy, O.A.; Abdel Lateif, K.S.; Seleiman, M.F.; Shami, A.; Albarakaty, F.M.; El-Meihy, M.R. Phylogenetic Diversity of Trichoderma Strains and Their Antagonistic Potential against Soil-Borne Pathogens under Stress Conditions. Biology 2020, 9, 189. [Google Scholar] [CrossRef]
- Hillocks, R. Farming with fewer pesticides: EU pesticide review and resulting challenges for UK agriculture. Crop. Prot. 2012, 31, 85–93. [Google Scholar] [CrossRef]
- Ribeiro, A.; Romeiras, M.M.; Tavares, J.; Faria, M.T. Ethnobotanical survey in Canhane village, district of Massingir, Mozambique: Medicinal plants and traditional knowledge. J. Ethnobiol. Ethnomed. 2010, 6, 33. [Google Scholar] [CrossRef]
- Ahmad, I.; Beg, A.Z. Antimicrobial and phytochemical studies on 45 Indian medicinal plants against multi-drug resistant human pathogens. J. Ethnopharmacol. 2001, 74, 113–123. [Google Scholar] [CrossRef]
- MartÍnez, J.A. Natural fungicides obtained from plants, fungicides for plant and animal diseases. In Fungicides for Plant and Animal Diseases; Dhanasekaran, D., Ed.; In Tech Open: Shanghai, China, 2012. [Google Scholar]
- Shurigin, V.; Alaylar, B.; Davranov, K.; Wirth, S.; Bellingrath-Kimura, S.D.; Egamberdieva, D. Diversity and biological activity of culturable endophytic bacteria associated with marigold (Calendula officinalis L.). AIMS Microbiol. 2021, 7, 336–353. [Google Scholar] [CrossRef] [PubMed]
- Hancock, R.D.; Hogenhout, S.; Foyer, C.H. Mechanisms of plant-insect interaction. J. Exp. Bot. 2015, 66, 421–424. [Google Scholar] [CrossRef] [PubMed]
- Shaaban, H.A.E.; El-Ghorab, A.H.; Shibamoto, T. Bioactivity of essential oils and their volatile aroma components: Review. J. Essen. Oil Res. 2012, 24, 203–212. [Google Scholar] [CrossRef]
- Zorzi-Tamazoni, G.; Schiavo-Griggio, E.; Pessin-Broilo, R.; Teresinha da silva Ribeiro, G.L.; Goncalvez-Soares; Schwambach, J. Screening for inhibitory activity of essential oils on fungal tomato pathogen Stemphylium solani Weber. Biocatal. Agric. Biotechnol. 2018, 16, 364–372. [Google Scholar] [CrossRef]
- Kishore, G.K.; Pande, S.; Harish, S. Evaluation of essential oils and their components for broadspectrum antifungal activity and control of late leaf spot and crown rot diseases in peanut. Plant Dis. 2007, 91, 375–379. [Google Scholar] [CrossRef]
- Barrera-Necha, L.L.; Garduño-Pizaña, C.; Garcīa-Bārrera, L.J. In vitro antifungal activity of essential oils and their compounds on mycelial growth of Fusarium oxysporum f. sp. gladioli (Massey) Snyder and Hansen. Plant Pathol. J. 2009, 8, 17–21. [Google Scholar] [CrossRef]
- Hyldgaard, M.; Mygind, T.; Meyer, R.L. Essential oils in food preservation: Mode of action, synergies, and interactions with food matrix components. Front. Microbiol. 2012, 3, 12. [Google Scholar] [CrossRef]
- De Jesus, C.; Santos Frazo, G.G.; Blank, A.F.; De Aquino Santana, L.C.L. Myrcia ovata cambessedes essential oils: A proposal for a novel natural antimicrobial against foodborne bacteria. Microb. Pathog. 2016, 99, 142–147. [Google Scholar] [CrossRef]
- Carson, C.F.; Hammer, K.A.; Riley, T.V. Melaleuca alternifolia (tea tree) oil: A review of antimicrobial and other medicinal properties. Clin. Microbiol. Rev. 2006, 19, 50–62. [Google Scholar] [CrossRef]
- Huang, X.; Liu, T.; Zhou, C.; Huang, Y.; Liu, X.; Yuan, H. Antifungal activity of essential oils from three artemisia species against Colletotrichum gloeosporioides of Mango. Antibiotics 2021, 10, 1331. [Google Scholar] [CrossRef]
- Pinto, E.; Vale-Silva, L.; Cavaleiro, C.; Salgueiro, L. Antifungal activity of the clove essential oil from Syzygium aromaticum on Candida, Aspergillus and Dermatophyte species. J. Med. Microbiol. 2009, 58, 1454–1462. [Google Scholar] [CrossRef]
- Bhuiyan, N.; Begum, J.; Nandi, N.; Akter, F. Constituents of the essential oil from leaves and buds of clove (Syzigium caryophyllatum). Afr. J. Plant Sci. 2010, 4, 451–454. [Google Scholar]
- Nowotarska, S.W.; Nowotarski, K.; Grant, I.R.; Elliott, C.T.; Friedman, M.; Situ, C. Mechanisms of antimicrobial action of cinnamon and oregano oils, cinnamaldehyde, carvacrol, 2,5-dihydroxybenzaldehyde, and 2-hydroxy-5-methoxybenzaldehyde against Mycobacterium avium subsp. paratuberculosis (Map). Foods 2017, 6, 72. [Google Scholar] [CrossRef]
- Arnal-Schnebelen, B.; Hadji-Minaglou, F.; Peroteau, J.F.; Ribeyre, F.; de Billerbeck, V.G. Essential oils in infectious gynaecological disease: A statistical study of 658 cases. Int. J. Aromather. 2004, 14, 192–197. [Google Scholar] [CrossRef]
- Raveau, R.; Fontaine, J.; Lounès-Hadj Sahraoui, A. Essential oils as potential alternative biocontrol products against plant pathogens and weeds: A review. Foods 2020, 9, 365. [Google Scholar] [CrossRef]
- Abdel-Kader, M.M.; El-Mougy, N.S.; Lashin, S.M. Essential oils and Trichoderma harzianum as an integrated control measure against Faba bean root rot pathogens. J. Plant Prot. Res. 2011, 51, 306–313. [Google Scholar] [CrossRef]
- Hamad, Y.K.; Fahmi, M.M.; Zaitoun, F.M.; Ziyada, S.M. Role of essential oils in controlling fungi that cause decline disease of guava. Int. J. Pure App. Biosci. 2015, 3, 143–151. [Google Scholar] [CrossRef]
- Saltos-Rezabala, L.A.; Silveira, P.R.D.; Tavares, D.G.; Moreira, S.I.; Magalhães, T.A.; Botelho, D.M.D.S.; Alves, E. Thyme essential oil reduces disease severity and induces resistance against Alternaria linariae in tomato plants. Horticulture 2022, 8, 919. [Google Scholar] [CrossRef]
- Aravena, R.; Besoain, X.; Riquelme, N.; Salinas, A.; Valenzuela, M.; Oyanedel, E.; Barros, W.; Olguin, Y.; Madrid, A.; Alvear, M.; et al. Antifungal nanoformulation for biocontrol of tomato root and crown rot caused by Fusarium oxysporum f. sp. radicis-lycopersici. Antibiotics 2021, 10, 1132. [Google Scholar] [CrossRef]
- Rawal, P.; Adhikari, R.S. Evaluation of antifungal activity of Zingiber officinale against Fusarium oxysporum f. sp. lycopersici. Adv. Appl. Sci. Res. 2016, 7, 5–9. [Google Scholar]
- Gull, I.; Saeed, M.; Shaukat, H.; Aslam, S.M.; Samra, Z.O.; Athar, A.M. Inhibitory effect of Allium sativum and Zingiber officinales extracts on clinically important drug resistant pathogenic bacteria. Ann. Clinic. Microbiol. Antimicrob. 2012, 11, 8. [Google Scholar] [CrossRef] [PubMed]
- Iotsor, B.I.; Iseghohi, F.; Oladoja, O.E.; Raji, O.R.; Yusuf, Z.; Oyewole, O.A. Antimicrobial activities of garlic and ginger extracts on some clinical isolates. Int. J. Biotechnol. 2019, 8, 59–65. [Google Scholar]
- Helal, G.A.; Sarhan, M.M.; Shahla, A.N.K.A.; Abou El-Khair, E.K. Effects of Cymbopogon citratus L. essential oil on the growth, morphogenesis and aflatoxin production of Aspergillus flavus ML2-strain. J. Basic Microbiol. 2007, 47, 5–15. [Google Scholar] [CrossRef] [PubMed]
- Das, L.; Godbole, S. Antifungal and phytochemical analysis of Lantana camara, Citrus limonum (lemon), Azadirachta indica (neem) and Hibiscus rosasinensis (China rose). J. Pharmacy Res. 2015, 9, 476–479. [Google Scholar]
- Rongai, D.; Milano, F.; Scio, E. Inhibitory effect of plant extracts on conidial germination of the phytopathogenic fungus Fusarium oxysporum. Am. J. Plant Sci. 2012, 3, 1693–1698. [Google Scholar] [CrossRef]
- Morsy, K.M.; Abdel-Monaim, M.F.; Mazen, M.M. Use of abiotic and biotic inducers for controlling fungal diseases and improving growth of alfalfa. World J. Agric. Sci. 2011, 7, 566–576. [Google Scholar]
- El-Shahir, A.A.; El-Wakil, D.A.; Latef, A.A.H.A.; Youssef, N.H. Bioactive compounds and antifungal activity of leaves and fruit methanolic extracts of Ziziphus spina-christi L. Plants 2022, 11, 746. [Google Scholar] [CrossRef]
- Sharma, A.; Sharma, N.K.; Srivastava, A.; Kataria, A.; Dubey, S.; Sharma, S.; Kundu, B. Clove and lemongrass oil based non-ionic nanoemulsión for suppressing the growth of plant pathogenic Fusarium oxysporum f. sp. lycopersici. Ind. Crops Prod. 2018, 123, 353–362. [Google Scholar] [CrossRef]
- Hu, L.B.; Zhou, W.; Zhang, T.; Yang, Z.M.; Xu, J.H.; Shi, Z.Q. Mechanism of inhibition to Fusarium moniliforme by antimicrobial peptide Fengycins. Microbiol. China 2010, 37, 251–255. [Google Scholar]
- Kawakami, K.; Inuzuka, H.; Hori, N.; Takahashi, N.; Ishida, K.; Mochizuki, K.; Ohkusu, K.; Muraosa, Y.; Watanabe, A.; Kamei, K. Inhibitory effects of antimicrobial agents against Fusarium species. Med. Mycol. 2015, 53, 603–611. [Google Scholar] [CrossRef]
- Nene, Y.L.; Thapliyal, P.N. Fungicides in Plant Disease Control, 3rd ed.; Oxford and IBH Publishing: New Delhi, India, 1993; pp. 311–348. [Google Scholar]
- Ahmed, H.F.A.; Seleiman, M.F.; Al-Saif, A.M.; Alshiekheid, M.A.; Battaglia, M.L.; Taha, R.S. Biological control of celery powdery mildew disease caused by Erysiphe heraclei DC in vitro and in vivo conditions. Plants 2021, 10, 2342. [Google Scholar] [CrossRef]
- Adil, W.; Ahlam, M.; Muneeb, R.; Seema, A.; Hussain, M. Bee Propolis (Bee’s Glue): A phytochemistry review. J. Crit. Rev. 2017, 4, 9–13. [Google Scholar]
- Goodman, R.N.; Király, Z.; Wood, K.R. The Biochemistry and Physiology of Plant Disease; University of Missouri Press: Columbia, MO, USA, 1986. [Google Scholar]
- Mohamed, M.S.M.; Saleh, A.M.; Abdel-Farid, I.B.; El-Naggar, A.A. Growth, hydrolases and ultrastructure of Fusarium oxysporum as affected by phenolic rich extracts from several xerophytic plants. Pestic. Bioch. Physiol. 2016, 141, 57–64. [Google Scholar] [CrossRef]
- Nicholson, R.L.; Hammerschmidt, R. Phenolic compounds and their role in disease resistance. Ann. Rev. Phytopathol. 1992, 30, 369–389. [Google Scholar] [CrossRef]
- Zaynab, M.; Fatima, M.; Abbas, S.; Sharif, Y.; Umair, M.; Zafar, M.H.; Bahadar, K. Role of secondary metabolites in plant defense against pathogens. Microb. Pathog. 2018, 124, 198–202. [Google Scholar] [CrossRef]
- Özçelik, B.; Deliorman Orhan, D.; Özgen, S.; Ergun, F. Antimicrobial activity of flavonoids against extended-spectrum ß-Lactamase (ES_L)-producing Klebsiella pneumonia. Trop. J. Pharm. Res. 2008, 7, 1151–1157. [Google Scholar] [CrossRef]
- Pastuszak, J.; Szczerba, A.; Dziurka, M.; Hornyák, M.; Kope’c, P.; Szklarczyk, M.; Płażek, A. Physiological and biochemical response to Fusarium culmorum infection in three durum wheat genotypes at seedling and full anthesis stage. Int. J. Mol. Sci. 2021, 22, 7433. [Google Scholar] [CrossRef]
- Dangl, J.; Jones, J.D.G. Plant pathogens and integrated defense responses to pathogens. Nature 2001, 411, 826–834. [Google Scholar] [CrossRef]
- Patykowski, J.; Urbanek, H. Activity of enzymes related to H2O2 generation and metabolism in leaf apoplastic fraction of tomato leaves infected with Botrytis cinerea. J. Phytopathol. 2003, 151, 153–161. [Google Scholar] [CrossRef]
- Niu, J.H.; Cao, Y.; Lin, X.G.; Leng, Q.Y.; Chen, Y.M.; Yin, J.M. Field and laboratory screening of Anthurium cultivars for resistance to foliar bacterial blight and the induced activities of defense-related enzymes. Folia Hort. 2018, 30, 129–137. [Google Scholar] [CrossRef]
- Yang, T.; Poovaiah, B.W. Hydrogen peroxide homeostasis: Activation of plant catalase by calcium/calmodulin. Proc. Natl. Acad. Sci. USA 2002, 99, 4097–4102. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.; Fartyal, D.; Agarwal, A.; Shukla, T.; James, D.; Kaul, T.; Negi, Y.K.; Arora, S.; Reddy, M.K. Abiotic stress tolerance in plants: Myriad roles of ascorbate peroxidase. Front. Plant Sci. 2017, 8, 581. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Xia, M.X.; Chen, J.; Yuan, R.; Deng, F.N.; Shen, F. Gene expression characteristics and regulation mechanisms of superoxide dismutase and its physiological roles in plants under stress. Biochemistry 2016, 81, 465–480. [Google Scholar] [CrossRef] [PubMed]
- Page, A.I.; Miller, R.H.; Keeney, D.R. Methods of Soil Analysis: Part. 2—Chemical and Microbiological Properties, 2nd ed.; American Society of Agronomy: Madison, WI, USA, 1982. [Google Scholar]
- Klute, A. Methods of Soil Analysis: Part. 1—Physical and Mineralogical Methods, 2nd ed.; American Society of Agronomy: Madison, WI, USA, 1986. [Google Scholar]
- Richards, L.F. Diagnosis and Improvement of Saline and Alkaline Soils (NO. 60); Soil and Water Conservative Research Branch, Agricultural Research Service, US Department of Agriculture: Washington, DC, USA, 1954.
- Phu, N.D.; Thy, L.H.P.; Lam, T.D.; Yen, V.H.; Lan, N.T.N. Extraction of jasmine essential oil by hydro-distillation method and applications on formulation of natural facial cleansers. IOP Conf. Ser. Mater. Sci. Eng. 2019, 542, 1–6. [Google Scholar]
- Reuveni, M.; Agapov, V.; Reuveni, M. Controlling powdery mildew caused by Sphaerotheca fuliginea in cucumber by foliar sprays of phosphate and potassium salts. Crop Prot. 1996, 15, 49–53. [Google Scholar] [CrossRef]
- Kumar, A.K.; Ramachandra, S.S.; Narsu, L. Pharmacognostic and phytochemical investigations of roots of Hibiscus micranthus Linn. Res. J. Pharm. Biol. Chem. Sci. 2010, 1, 324–337. [Google Scholar]
- Sahi, I.Y.; Khalid, A.N. In vitro biological control of Fusarium oxysporum causing wilt in Capsicum annuum. Mycopathol. 2007, 5, 85–88. [Google Scholar]
- Dhingra, O.D.; Sinclair, J.B. Basic Plant Pathology Methods, 2nd ed.; CRC-Press, Inc.: Boca Raton, FL, USA, 1995. [Google Scholar] [CrossRef]
- Dhingra, O.D.; Sinclair, J.B. Basic Plant Pathology Methods; CRC Press Inc.: Boca Raton, FL, USA, 1985; p. 355. [Google Scholar]
- Gilman, J.C. A Manual of Soil Fungi, 2nd ed.; Iowa State Univ. Press: Ames, IA, USA, 1957; p. 450. [Google Scholar]
- Reid, D.A.; Hayward, A.C.; Waterston, J.M. CMI descriptions of pathogenic fungi and bacteria. Kew Bull. 1965, 19, 414. [Google Scholar] [CrossRef]
- Barnett, H.L.; Hunter, B.B. Illustrated Genera of Imperfect Fungi, 4th ed.; Macmillan Publishing Co.: New York, NY, USA, 1986. [Google Scholar]
- Adjou, E.S.; Kouton, S.; Dahouenon-Ahoussi, E.; Sohounhloue, C.K.; Soumanou, M.M. Antifungal activity of Ocimum canum essential oil against toxinogenic fungi isolated from peanut seeds in post-harvest in Benin. Int. Res. J. Biol. Sci. 2012, 1, 20–26. [Google Scholar]
- Pinto, C.M.F.; Maffia, L.A.; Casali, V.W.D.; Cardoso, A.A. In vitro effect of plant leaf extracts on mycelial growth and sclerotial germination of Sclerotium cepivorum. J. Phytopathol. 1998, 146, 421–425. [Google Scholar] [CrossRef]
- Whitehead, M.D. Sorghum, a medium suitable for the increase of inoculum for studies of soil-borne and certain other fungi. Phytopathology 1957, 47, 450. [Google Scholar]
- Chavdarov, P. Evaluation of lentil germplasm for disease resistance to fusarium wilt (Fusarium oxysporum f. sp. lentis). Cent. Eur Agric. 2006, 7, 121–126. [Google Scholar]
- Alirezalu, A.; Salehi, P.; Ahmadi, N.; Sonboli, A.; Aceto, S.; Maleki, H.; Ayyari, M. Flavonoids profile and antioxidant activity in flowers and leaves of hawthorn species (Crataegus spp.) from different regions of Iran. Int. J. Food Prop. 2018, 21, 452–470. [Google Scholar] [CrossRef]
- Georgé, S.; Brat, P.; Alter, P.; Amiot, M.J. Rapid determination of polyphenols and vitamin C in plant-derived products. J. Agric. and Food Chemistry 2005, 53, 1370–1373. [Google Scholar] [CrossRef]
- Gurnani, N.; Gupta, M.; Mehta, D.; Mehta, B.K. Chemical composition, total phenolic and flavonoid contents, and in vitro antimicrobial and antioxidant activities of crude extracts from red chili seeds (Capsicum frutescens L.). J. Taibah Univ. Sci. 2016, 10, 462–470. [Google Scholar] [CrossRef]
- Wagner, G.J. Content and vacuole/extra vacuole distribution of neutral sugars, free amino acids, and anthocyanin in protoplasts. Plant Physiol. 1979, 64, 88–93. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–380. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Sohrabi, Y.; Heidari, G.; Weisany, W.; Golezani, K.G.; Mohammadi, K. Changes of antioxidative enzymes, lipid peroxidation and chlorophyll content in chickpea types colonized by different Glomus species under drought stress. Symbiosis 2012, 56, 5–18. [Google Scholar] [CrossRef]
- Caverzan, A.; Passaia, G.; Rosa, S.B.; Ribeiro, C.W.; Lazzarotto, F.; Margis-Pinheiro, M. Plant responses to stresses: Role of ascorbate peroxidase in the antioxidant protection. Genet. Mol. Biol. 2012, 35, 1011–1019. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Species of Fungi | Occurrence of Fungal Isolates | Total Isolates | Frequency (%) | ||
|---|---|---|---|---|---|
| Beni-Suef District | Ihnasia District | Somosta District | |||
| F. nygamai | 3 | 0 | 2 | 5 | 4.6 |
| F. oxysporum | 8 | 10 | 7 | 25 | 22.9 |
| F. solani | 9 | 5 | 6 | 20 | 18.3 |
| F. verticillioides | 3 | 4 | 2 | 9 | 8.3 |
| P. splendens | 0 | 4 | 7 | 11 | 10.1 |
| R. solani | 5 | 9 | 8 | 22 | 20.2 |
| S. sclerotiorum | 4 | 8 | 5 | 17 | 15.6 |
| Total | 32 | 40 | 37 | 109 | 100 |
| Tested Fungi | Damping-Off % | Root Rot/ Wilt % | Total Infection % | * Survived Plants % | |
|---|---|---|---|---|---|
| Pre- Emergence % | Post- Emergence % | ||||
| F. oxysporum | 13.3 ± 1.02 c | 20.0 ± 1.24 b | 26.7 ± 1.89 b | 60.0 | 40.0 |
| F. solani | 20.0 ± 1.04 b | 13.3 ± 1.37 c | 20.0 ± 1.40 c | 53.3 | 46.7 |
| P. splendens | 20.0 ± 1.06 b | 6.7 ± 0.66 d | 0.0 ± 0.0 d | 26.7 | 73.3 |
| R. solani | 13.3 ± 0.97 c | 26.7 ± 1.17 a | 33.3 ± 1.04 a | 73.3 | 26.7 |
| S. sclerotiorum | 26.7 ± 1.31 a | 13.3 ± 1.04 c | 26.7 ± 2.55 b | 66.7 | 33.3 |
| Control | 0.0 ± 0.0 d | 0.0 ± 0.0 e | 0.0 ± 0.0 d | 0.0 | 100 |
| Essential Oil Application | Conc. (mL/L) | * Mycelium Growth Inhibition % | |||
|---|---|---|---|---|---|
| R. solani | S. sclerotiorum | F. solani | F. oxysporum | ||
| Cinnamomum zeylanicum | 0.25 | 56.3 ± 1.87 g | 60.2 ± 1.74 g | 59.1 ± 1.67 g | 58.2 ± 1.61 f |
| 0.5 | 62.4 ± 1.44 e | 65.9 ± 1.94 e | 65.4 ± 1.96 ef | 63.7 ± 1.69 e | |
| 1.0 | 78.2 ± 2.02 b | 81.1 ± 1.90 b | 80.9 ± 1.39 b | 79.4 ± 1.35 b | |
| Mean | 65.6 | 69.1 | 68.5 | 67.1 | |
| Mentha piperita | 0.25 | 45.4 ± 1.96 i | 50.2 ± 1.74 i | 52.6 ± 1.64 i | 45.7 ± 1.94 h |
| 0.5 | 58.3 ± 2.01 f | 63.0 ± 1.26 f | 64.3 ± 1.63 f | 60.6 ± 1.98 f | |
| 1.0 | 68.7 ± 2.09 d | 70.0 ± 2.62 d | 72.4 ±1.41 d | 69.1 ± 1.67 d | |
| Mean | 57.5 | 61.0 | 63.1 | 58.5 | |
| Syzygium aromaticum | 0.25 | 61.9 ± 2.39 e | 65.2 ± 1.91 ef | 67.0 ± 1.16 e | 63.5 ± 1.68 e |
| 0.5 | 74.3 ± 1.75 c | 76.5 ± 2.25 c | 76.7 ± 1.35 c | 75.0 ± 1.05 c | |
| 1.0 | 85.4 ± 1.99 a | 100 ± 0.0 a | 100 ± 0.0 a | 90.2 ± 2.59 a | |
| Mean | 73.8 | 80.6 | 81.2 | 76.2 | |
| Thymus vulgaris | 0.25 | 31.1 ± 1.83 k | 34.6 ± 1.97 k | 32.4 ± 2.29 k | 30.6 ± 2.42 i |
| 0.5 | 43.0 ± 1.61 j | 46.9 ± 2.04 j | 45.9 ± 2.38 j | 44.1 ± 2.59 h | |
| 1.0 | 55.2 ± 2.02 h | 57.2 ± 1.95 h | 56.3 ± 1.48 h | 53.2 ± 1.85 g | |
| Mean | 43.1 | 46.2 | 44.9 | 42.6 | |
| Plant Extract Application | Conc. (%) | * Mycelium Growth Inhibition % | |||
|---|---|---|---|---|---|
| R. solani | S. sclerotiorum | F. solani | F. oxysporum | ||
| Cymbopogon citratus | 5 | 45.2 ± 1.47 g | 47.6 ± 2.09 f | 51.9 ± 2.68 f | 49.3 ± 2.48 f |
| 10 | 50.9 ± 2.03 e | 53.7 ± 1.35 e | 55.0 ± 1.73 ef | 56.5 ± 1.50 d | |
| 20 | 64.8 ± 1.45 b | 67.2 ± 1.73 b | 69.1 ± 2.61 b | 68.2 ± 1.61 b | |
| Mean | 53.6 | 56.2 | 58.6 | 58.0 | |
| Lantana camara | 5 | 29.6 ± 1.87 j | 33.2 ± 1.76 h | 33.5 ± 1.29 h | 29.3 ± 1.62 i |
| 10 | 42.0 ± 1.70 h | 45.4 ± 2.02 g | 47.6 ± 2.02 g | 43.5 ± 1.68 g | |
| 20 | 55.2 ± 2.44 d | 58.0 ± 2.48 d | 60.6 ± 1.28 cd | 56.3 ± 1.51 d | |
| Mean | 42.3 | 45.5 | 47.2 | 43.0 | |
| Ocimum basilicum | 5 | 23.7 ± 1.35 k | 0.0 ± 0.0 i | 21.3 ± 2.80 i | 0.0 ± 0.0 j |
| 10 | 39.1 ± 1.67 i | 44.3 ± 1.66 g | 45.9 ± 2.40 g | 41.9 ± 2.30 h | |
| 20 | 47.6 ± 1.78 f | 49.6 ± 2.40 f | 53.9 ± 1.01 f | 50.9 ± 2.48 f | |
| Mean | 36.8 | 31.3 | 40.4 | 30.9 | |
| Zingiber officinales | 5 | 51.3 ± 1.85 e | 55.2 ± 2.52 e | 57.6 ± 2.10 de | 53.7 ± 2.09 e |
| 10 | 58.2 ± 1.69 c | 61.3 ± 1.91 c | 61.9 ± 1.78 c | 60.0 ± 1.82 c | |
| 20 | 72.0 ± 1.70 a | 76.5 ± 2.11 a | 77.4 ± 1.90 a | 73.5 ± 1.81 a | |
| Mean | 60.5 | 64.3 | 65.6 | 62.4 | |
| Treatments | Application Method | * Disease Reduction Rate % | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| R. solani | S. sclerotiorum | F. solani | F. oxysporum | |||||||
| SS | FS | DO% | RR% | DO% | RR% | DO% | RR% | DO% | W% | |
| Conc. (mL/L) | Essential Oils (EOs) | |||||||||
| A | 1.0 | 0.5 | 32.1 ± 0.95 f | 30.8 ± 1.53 f | 41.3 ± 1.83 f | 39.9 ± 1.87 f | 45.6 ± 1.80 f | 44.0 ± 1.09 g | 36.0 ± 0.90 g | 36.1 ± 0.82 g |
| 1.5 | 0.75 | 51.1 ± 1.87 c | 51.9 ± 2.30 c | 60.4 ± 2.25 d | 61.6 ± 2.12 d | 65.0 ± 2.36 d | 66.2 ± 1.53 d | 55.4 ± 2.01 d | 57.2 ± 1.66 d | |
| 3.0 | 1.5 | 65.4 ± 1.94 b | 67.7 ± 2.14 b | 74.8 ± 1.44 b | 67.8 ± 1.64 c | 79.5 ± 1.52 b | 82.9 ± 1.42 b | 69.9 ± 1.45 b | 73.0 ± 1.13 b | |
| B | 1.0 | 0.5 | 13.1 ±1.09 h | 9.8 ± 0.57 h | 22.1 ± 0.83 h | 18.3 ± 0.60 h | 26.3 ± 1.05 h | 21.8 ± 1.20 i | 16.7 ± 0.92 i | 15.1 ± 0.18 i |
| 1.5 | 0.75 | 32.1 ± 0.88 f | 30.8 ± 2.19 f | 41.3 ± 1.38 f | 39.9 ± 1.46 f | 45.6 ± 0.80 f | 44.0 ± 1.82 g | 36.0 ± 0.94 g | 36.1 ± 1.36 g | |
| 3.0 | 1.5 | 51.1 ± 1.95 c | 51.9 ± 2.03 c | 60.4 ± 1.38 d | 61.6 ± 1.59 d | 65.0 ± 1.40 d | 66.2 ± 0.95 d | 55.4 ± 1.22 d | 57.2 ± 1.09 d | |
| C | 1.0 | 0.5 | 46.4 ± 1.15 d | 46.6 ± 1.74 d | 50.8 ± 1.33 e | 50.8 ± 1.47 e | 55.3 ± 1.08 e | 55.1 ± 1.36 f | 50.5 ± 1.13 e | 51.9 ± 2.43 e |
| 1.5 | 0.75 | 65.4 ± 1.41 b | 67.7 ± 1.41 b | 70.0 ± 1.97 c | 72.4 ± 1.44 b | 74.6 ± 1.33 c | 77.3 ± 1.94 c | 65.0 ± 1.43 c | 67.7 ± 1.33 c | |
| 3.0 | 1.5 | 75.0 ± 2.07 a | 78.2 ± 1.84 a | 79.6 ± 1.44 a | 83.2 ± 1.59 a | 84.3 ± 1.10 a | 88.5 ± 1.34 a | 79.5 ± 1.93 a | 83.5 ± 0.89 a | |
| D | 1.0 | 0.5 | 5.5 ± 0.42 i | 8.3 ± 0.36 h | 14.5 ± 0.84 i | 9.5 ± 0.43 i | 18.6 ± 0.66 i | 16.7 ± 0.84 j | 9.0 ± 0.58 j | 6.5 ± 0.27 j |
| 1.5 | 0.75 | 25.6 ± 1.72 g | 22.3 ± 2.57 g | 33.7 ± 2.06 g | 31.1 ± 1.65 g | 37.9 ± 1.30 g | 34.9 ± 0.92 h | 28.4 ± 1.48 h | 27.6 ± 1.04 h | |
| 3.0 | 1.5 | 43.6 ± 1.60 e | 43.4 ± 1.03 e | 52.8 ± 1.28 e | 50.8 ± 1.49 e | 57.3 ± 1.58 e | 58.1 ± 1.70 e | 47.7 ± 1.82 f | 48.6 ± 1.30 f | |
| Conc. (%) | Plant Extracts (PEs) | |||||||||
| E | 10 | 5 | 22.3 ± 1.24 g | 20.6 ± 1.18 g | 26.7 ± 2.79 g | 24.1 ± 1.36 g | 30.9 ± 1.89 f | 27.9 ± 1.81 g | 21.4 ± 2.01 i | 20.6 ± 1.43 i |
| 20 | 10 | 41.4 ± 2.12 d | 41.6 ± 1.55 d | 45.8 ± 1.80 c | 45.8 ± 2.95cd | 45.4 ± 0.70 d | 44.6 ± 1.04 e | 40.7 ± 0.74 e | 41.6 ± 1.62 e | |
| 40 | 20 | 55.7 ± 0.84 b | 57.4 ± 1.66 b | 55.4 ± 1.03 b | 56.6 ± 2.86 b | 60.0 ± 1.20 b | 61.2 ± 1.18 c | 55.2 ± 0.71 b | 57.4 ± 1.67 b | |
| F | 10 | 5 | 9.1 ± 0.24 i | 5.8 ± 0.29 i | 13.3 ± 0.34 i | 8.9 ± 0.31 h | 12.6 ± 0.60 g | 6.7 ± 0.15 h | 7.9 ± 0.28 k | 5.8 ± 0.17 k |
| 20 | 10 | 28.1 ± 1.0 f | 26.8 ± 1.30 f | 32.4 ± 2.29 f | 30.5 ± 0.97 f | 31.9 ± 1.90 f | 28.9 ± 1.30 g | 27.2 ± 2.62 h | 26.8 ± 1.24 h | |
| 40 | 20 | 42.4 ± 1.10 d | 42.6 ± 0.93 d | 46.8 ± 1.17 c | 46.8 ± 1.44 c | 51.3 ± 0.97 c | 51.1 ± 1.34 d | 46.5 ± 0.85 d | 47.9 ± 1.68 d | |
| G | 10 | 5 | 0.0 ± 0.0 j | 0.0 ± 0.0 j | 0.0 ± 0.0 j | 0.0 ± 0.0 i | 2.9 ± 0.22 h | 0.0 ± 0.0 i | 0.0 ± 0.0 l | 0.0 ± 0.0 l |
| 20 | 10 | 13.8 ± 0.99 h | 11.1 ± 0.44 h | 22.9 ± 1.21 h | 19.7 ± 0.60 g | 12.6 ± 0.61 g | 6.7 ± 0.31 h | 17.5 ± 0.87 j | 16.3 ± 0.97 j | |
| 40 | 20 | 32.9 ± 1.97 e | 32.1 ± 0.99 e | 42.0 ± 1.08 d | 41.3 ± 1.68 d | 41.6 ± 1.95 e | 40.0 ± 1.61 f | 36.9 ± 1.08 f | 37.4 ± 1.23 f | |
| H | 10 | 5 | 32.6 ± 2.23 e | 32.4 ± 1.51 e | 37.0 ± 1.72 e | 36.3 ± 0.68 e | 41.4 ± 1.22 e | 40.6 ± 1.40 f | 31.9 ± 3.05 g | 32.4 ± 2.20 g |
| 20 | 10 | 51.7 ± 2.02 c | 53.4 ± 1.46 c | 56.2 ± 1.29 b | 58.0 ± 1.86 b | 60.8 ± 1.01 b | 68.3 ± 1.75 b | 51.2 ± 2.87 c | 53.4 ± 2.06 c | |
| 40 | 20 | 61.2 ± 1.20 a | 64.0 ± 1.72 a | 65.8 ± 1.51 a | 68.8 ± 9.52 a | 70.5 ± 2.07 a | 73.9 ± 2.10 a | 60.9 ± 0.74 a | 64.0 ± 1.56 a | |
| Conc. (g/L) | Fungicides | |||||||||
| I | 0.25 | 0.125 | 79.2 ± 2.03 d | 74.1 ± 1.22 e | 83.2 ± 2.24 d | 78.3 ± 1.61 d | 88.4 ± 0.74 b | 84.8 ± 3.24 c | 77.1 ± 2.60 d | 73.0 ± 1.89 e |
| 0.50 | 0.25 | 93.5 ± 2.30 b | 90.3 ± 2.42 c | 95.4 ± 2.43 b | 93.1 ± 1.99 b | 100 ± 0.0 a | 96.1 ± 2.57 b | 94.0 ± 2.02 b | 88.6 ± 2.09 c | |
| 1.00 | 0.50 | 97.8 ± 2.36 a | 94.6 ± 1.86 b | 98.2 ± 3.15ab | 95.9 ± 1.17 b | 100 ± 0.0 a | 100 ± 0.0 a | 97.4 ± 2.42ab | 91.4 ± 1.87 c | |
| J | 0.25 | 0.125 | 85.7 ± 1.79 c | 83.6 ± 1.84 d | 90.4 ± 1.92 c | 94.6 ± 2.0 b | 100 ± 0.0 a | 100 ± 0.0 a | 85.5 ± 1.68 c | 85.2 ± 1.67 d |
| 0.50 | 0.25 | 100 ± 0.0 a | 94.6 ± 1.02 b | 100 ± 0.0 a | 89.2 ± 2.56 c | 100 ± 0.0 a | 100 ± 0.0 a | 100 ± 0.0 a | 94.7 ± 2.38 b | |
| 1.00 | 0.50 | 100 ± 0.0 a | 100 ± 0.0 a | 100 ± 0.0 a | 100 ± 0.0 a | 100 ± 0.0 a | 100 ± 0.0 a | 100 ± 0.0 a | 100 ± 0.0 a | |
| Treatments | Application Method/Conc. | Damping-Off % | Root Rot % | Wilt % | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Pre-Emergence % | Post-Emergence % | |||||||||
| SS | FS | DI % | * R % | DI % | * R % | DI % | * R % | DI % | * R % | |
| C. zeylanicum EO | 3 mL/L | 1.5 mL/L | 18.6 ± 1.40 f | 69.2 | 21.4 ± 1.46 d | 61.3 | 11.6 ± 1.38 e | 73.2 | 19.7 ± 1.32 f | 68.4 |
| M. piperita EO | 3 mL/L | 1.5 mL/L | 29.7 ± 1.89 c | 50.6 | 25.7 ± 1.90 c | 53.5 | 17.1 ± 1.28 c | 60.4 | 30.1 ± 1.78 c | 51.7 |
| S. aromaticum EO | 3 mL/L | 1.5 mL/L | 14.9 ± 1.52 g | 75.2 | 15.8 ± 1.38 e | 71.4 | 8.3 ± 1.13 f | 80.7 | 16.4 ±1.23 g | 73.7 |
| C. citratus PE | 40 % | 20 % | 26.4 ± 1.39 d | 56.1 | 22.6 ± 2.01 d | 59.0 | 15 ± 1.26 cd | 65.3 | 26.1 ± 0.94 d | 58.2 |
| L. camara PE | 40 % | 20 % | 35.1 ± 1.80 b | 41.7 | 30.2 ± 1.53 b | 45.2 | 20.3 ± 1.02 b | 52.8 | 34.1 ± 0.98 b | 45.3 |
| Z. officinales PE | 40 % | 20 % | 22.1 ± 0.95 e | 63.4 | 20.4 ± 0.79 d | 63.0 | 12.9 ± 1.04 de | 70.1 | 22.9 ± 1.34 e | 63.3 |
| Carbendazim | 1.0 g/L | 0.5 g/L | 2.5 ± 0.42 h | 95.8 | 3.2 ± 0.21 f | 94.2 | 4.6 ± 0.61 g | 89.4 | 5.3 ± 0.76 h | 91.6 |
| Control | – | – | 60.2 ± 2.04 a | – | 55.1 ± 2.11 a | – | 43.1 ± 1.83 a | – | 62.3 ± 2.44 a | – |
| Common/Scientific Name | Family/Part Used | Target Pathogens | References |
|---|---|---|---|
| Cinnamon (Cinnamomum zeylanicum) | Lauraceae/ bark | Fusarium spp., Aspergillus spp., Botrytis cinerea, Colletotrichum gloerpoides, C. musae, Phytophthora palmivora, Sclerotinia sclerotiorum, Lasidiploidia theobromae, Alternaria citrii, Rhizoctonia solani, and Penicillium expansum | [30,31,32] |
| Lemongrass (Cymbopogon citratus) | Gramineae/ leaves | Aspergillus spp., F. oxysporum, and F. solani | [18,19,20] |
| Lantana (Lantana camara) | Verbenaceae/ leaves | Fusarium spp., and Rhizoctonia solani | [21,22] |
| Peppermint (Mentha piperita) | Lamiaceae/ leaves | Aspergillus spp., Fusarium spp., Rhizoctomia saloni, Macrophomina phaseolina, Sclerotinia minör, S. sclerotiorum, Alternaria alternaria, Colletotrichum musae, and C. gloesporioides | [19,23,33] |
| Basil (Ocimum basilicum) | Lamiaceae/ leaves | Sclerotinia sclerotiorum, Stromatinia cepivora, Fusarium oxysporum, Verticillium dahlia, Mucor racemosus, Penicillium spp., Rhizoctomia saloni, and Macrophomina phaseolina, | [23,24,25] |
| Clove (Syzygium aromaticum) | Myrtaceae/ flower buds | Aspergillus spp., Fusarium oxysporum, Penicillium citrinum, Rhizopus nigricans, Rhizoctonia solani, and Botryotinia fuckeliana | [25,34,35] |
| Thyme (Thymus vulgaris) | Lamiaceae/ Leaves and flowers | Aspergillus spp., Fusarium spp., Penicillium expansum, P. brevicompactum, Botrytis cinerea, B. fuckeliana, and Rhizoctonia solani | [24,35] |
| Ginger (Zingiber officinales) | Zingiberaceae Rhizomes | Aspergillus spp., Penicillium citrinum, P. chrysogenum, F. verticillioides, F. oxysporum, Rhizoctonia solani, and Sclerotinia sclerotiorum | [23,25,26] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, H.F.A.; Seleiman, M.F.; Mohamed, I.A.A.; Taha, R.S.; Wasonga, D.O.; Battaglia, M.L. Activity of Essential Oils and Plant Extracts as Biofungicides for Suppression of Soil-Borne Fungi Associated with Root Rot and Wilt of Marigold (Calendula officinalis L.). Horticulturae 2023, 9, 222. https://doi.org/10.3390/horticulturae9020222
Ahmed HFA, Seleiman MF, Mohamed IAA, Taha RS, Wasonga DO, Battaglia ML. Activity of Essential Oils and Plant Extracts as Biofungicides for Suppression of Soil-Borne Fungi Associated with Root Rot and Wilt of Marigold (Calendula officinalis L.). Horticulturae. 2023; 9(2):222. https://doi.org/10.3390/horticulturae9020222
Chicago/Turabian StyleAhmed, Hamada F. A., Mahmoud F. Seleiman, Ibrahim A. A. Mohamed, Ragab S. Taha, Daniel O. Wasonga, and Martin L. Battaglia. 2023. "Activity of Essential Oils and Plant Extracts as Biofungicides for Suppression of Soil-Borne Fungi Associated with Root Rot and Wilt of Marigold (Calendula officinalis L.)" Horticulturae 9, no. 2: 222. https://doi.org/10.3390/horticulturae9020222