
Preharvest UV-B Treatment Improves Strawberry Quality and Extends Shelf Life


Food Quality Laboratory, USDA-ARS, Beltsville, MD 20705, USA
*
Author to whom correspondence should be addressed.
Horticulturae 2023, 9(2), 211; https://doi.org/10.3390/horticulturae9020211
Submission received: 29 December 2022
/
Revised: 30 January 2023
/
Accepted: 31 January 2023
/
Published: 5 February 2023
(This article belongs to the Collection New Advances in Fruit Quality: Pre-harvest Techniques and Postharvest Management)
Abstract
:Postharvest ultraviolet B (UV-B) radiation has been used to control pathogen incidence on fresh produce, but little attention has been paid to preharvest UV-B effect on strawberry fruit quality and storage. In this study, strawberry (Fragaria × ananassa Duch. cv Albion) plants grown in greenhouse were irradiated with UV-B lamps with an intensity of 0.7362 J/s m2 for 1 and 2 h every day from 5–7 pm after flowering. Fruits were harvested at red stage for quality evaluation. The results indicated that the preharvest UV-B-treated fruits had lighter color (increased L*) than the control. Total soluble solids (TSS), total phenolics content (TPS) and total anthocyanin content (TAC) of UV-B-treated fruits were higher than for the control fruits. There were no significant differences in any of the quality traits between UV-B one-hour and two-hour treatments. We further evaluated the fruit quality when stored at 5 °C on day 7 and day 14. Compared to untreated control, UV-B-treated fruits retarded decrease in L*, TSS, TPS, TAC and firmness, and reduced fruit decay. Examination of expression of genes related to UV-B signaling indicated that HY5 was the major component of UV-B signaling during the green and white stages. However, anthocyanin genes were highly responsive to UV-B treatment during the red stage. Our results suggest that utilization of a low dose of UV-B radiation during the growth stage can improve strawberry fruit quality, and extend shelf life. This research facilitates the utilization of preharvest UV-B treatment for improving fruit quality in controlled environment agriculture.
1. Introduction
Ultraviolet radiation (UV) is a form of electromagnetic radiation with a light spectrum from 200 to 400 nm. It usually includes three classes, UV-A, UV-B and UV-C, based on the wavelength. UV-C is short-wave UV, below 280 nm, that can kill pathogens and trigger the biosynthesis of secondary metabolites in plants [1,2]. However, UV-C often causes damage to plants, too [3]. On the other hand, UV-B (280–315 nm) radiation affects many aspects of plant growth and development, such as control of plant height and yield, and acts at the molecular level on signal transduction and gene expression [1,4]. The mechanism of UV-B perception and signal transduction involves UV Resistance locus 8 (UVR8) and the bZIP transcription factor Elongated Hypocotyl 5 (HY5) [5]. UVR8 itself is a constitutively expressed protein that functions as a UV-B photoreceptor regulating HY5 expression [5].
Postharvest UV-B treatments have been used to control postharvest diseases, extend shelf life, and improve nutritional benefits of many vegetables and fruits [6,7]. Exposure to low-dose UV-B radiation has been shown to reduce broccoli weight loss, delay yellowing, and improve chlorophyll retention [8]. In grapes, a higher level of resveratrol was identified in the skins of UV-B-treated grapes stored for 7 d under cold conditions [9]. UV-B radiation can stimulate the expression of genes involved in anthocyanin biosynthesis in blueberry fruit [10]. In tomato, postharvest UV-B irradiation maintained fruit sensory qualities and enhanced antioxidant capacity during storage [11]. The content of cyanidin 3-glucoside, quercetin 3-glucuronide and kaempferol 3-glucoside in strawberry increased with solar UV-B treatment [12]. The beneficial effect of UV-B on anthocyanin accumulation also has been demonstrated in various fruits, including apples [13], grapes [9], pears [14], and blueberries [15]. Expression of several anthocyanin biosynthetic genes, such as flavonol synthase (FLS) and hydroxycinnamoyl CoA: shikimate hydroxycinnamoyl transferase (HCT), was upregulated by UV-B [16,17,18,19]. However, limited research has been performed on the effects of preharvest UV-B application.
Strawberry is one of the most popular horticultural crops due to its characteristic flavor and nutraceutical value. Strawberry fruits are rich in natural antioxidants, such as anthocyanins [20], which are beneficial for human health [21]. Most strawberries grow in open fields. However, as the cost of growing fruit in open fields increases due to water shortages and challenges with pests, as well as increasing consumer demand regarding fruit quality, the strawberry production in controlled environment is increasing, especially in urban areas [22]. The controlled environment agriculture provides opportunities to optimize the fruit development including light control to obtain the desired produce. In this study, we utilized UV-B irradiation to treat strawberry fruits during the growth stage in a greenhouse and investigated its impact on fruit quality and shelf life.
2. Materials and Methods
2.1. Plant and Fruit Materials
Strawberry plants (Fragaria × ananassa Duch. L. cv. Albion) were grown in a greenhouse at 26 °C with 14 h light (from 6 am to 8 pm) every day. The average light intensity over the plant growth period was about 140 μmol/s m2, and no UV-B irradiation was detected inside the greenhouse. Fruits were harvested at three developmental stages based on size and color of receptacles and achenes: green stage, young fruit with green achenes and receptacle; white stage, white receptacles and tanning achenes; red stage, full size fruit with fully red receptacles.
2.2. UV-B Treatments
Strawberry plants were irradiated with UV-B lamps (280–320 nm, Q-Panel 313, Cleveland, OH, USA). The lamps were ~30 cm above the plants with an intensity of 0.7362 J/s m2 for 1 and 2 h every day from 5–6 or 5–7 pm after flowering. The experiment was completely randomized with 3 treatments: (1) no UV-B radiation (control); (2) UV-B for 1 h; and (3) UV-B for 2 h. Fruits at the red stage were harvested and used for postharvest quality trait analyses.
2.3. Analysis of Color, Texture, Water Loss and Decacy Incidence
Fruit surface color was determined with a Chroma meter CR-400 (Konica Minolta Sensing, Inc., Tokyo, Japan) as described [23]. The chromaticity coordinates are: L* for lightness (black = 0, white = 100), a* for red/green color (redness > 0, greenness < 0) and b* for yellow/blue color (yellowness > 0, blueness < 0). Chroma (C) was calculated using the equation . A total of 24 fruits from each treatment were measured.
Firmness was assessed as the maximum penetration force (N) reached during tissue breakage using a TA-XTplus Texture Analyzer (Texture Technologies Corp, New York City, NY, USA) with a 10-mm-diameter cylinder probe [23]. The penetration depth and the cross-head speed were 3 mm and 1 mm s−1, respectively. Fruits were sliced into halves, and each half was measured in the central zone. Eighteen fruits of each treatment were used for fruit firmness evaluation.
Water loss and decay incidence were calculated on the fruit weight changes and fruit decay ratios during storage. Twelve fruits were put in a clear plastic egg carton. A total of 120 fruits (10 cartons) were used for each treatment. The fruits were kept at a temperature of 5 °C and 75% relative humidity.
2.4. Determination of Quality Attributes
Total phenolic content (TPC) was evaluated following a method described previously [24], with modifications. Briefly, 100 mg of lyophilized fruit powder was added into 5 mL of 80% methanol, and centrifuged at 6650× g for 10 min. Then, 250 μL aliquots of the supernatants were mixed with 25 μL of 0.1% Fast Blue BB solution and 25 μL of 5% NaOH. The absorbance of samples was measured at 420 nm using a high-performance microplate spectrophotometer (Agilent, Santa Clara, CA, USA). Results are expressed as milligrams of gallic acid equivalent (mg GAE) per gram of dry weight. The measurements were performed in triplicate.
Total anthocyanin contents (TAC) were assessed using a pH differential method [25]. Absorbance at 510 nm and at 700 nm in buffer at pH 1.0 and pH 4.5 was calculated using A = (A510 − A700) pH1.0 − (A510 − A700) pH4.5 with a molar extinction coefficient for pelargonidin-3-glucoside of 15,600. Results are expressed as milligrams of pelargonidin-3 glucoside equivalent (mg PGE) per gram of dry weight. The measurements were performed in triplicate.
Total soluble solids (TSS) were estimated as described [23] with an Atago PR-101 digital refractometer (Atago Co., Ltd., Tokyo, Japan) at 20 °C. The measurements were performed in triplicate.
2.5. RNA Extraction and RT-qPCR
Total RNA was extracted from frozen tissue using the RNeasy Plant Mini Kit (Qiagen, Germantown, MD, USA). Reverse transcription reaction and quantitative real-time PCR (RT-qPCR) was essentially performed as described [26]. One μg of total RNA was used to synthesize cDNA with iScript kit (Bio-RAD, Hercules, CA, USA), and PCR analysis of cDNA was conducted on a CFX96 Real-Time System (Bio-RAD, Hercules, CA, USA). The gene-specific primers were designed with Primer3 software (http://bioinfo.ut.ee/primer3-0.4.0/primer3/ (accessed on 1 January 2023) from their coding regions, where they share low homology between the gene paralogues. The primer pairs used in this study are listed in Table 1. All the reactions were run in triplicate from three independent experiments. Relative quantification of specific genes was determined using the cycle threshold 2ΔΔCt method. Relative expression levels were normalized based on the housekeeping gene FaACTIN (JN616288).
2.6. Statistical Analysis
SPSS (SPSS for Windows, SPSS Inc., Chicago, IL, USA) was used for statistical analysis. Two-way ANOVA was performed, and Tukey–Kramer post hoc test at a 95% confidence level was used to evaluate the effect of postharvest storage and the UV-B irradiation at a significance level of 0.05 (p < 0.05).
3. Results
3.1. Effect on Fruit Appearance
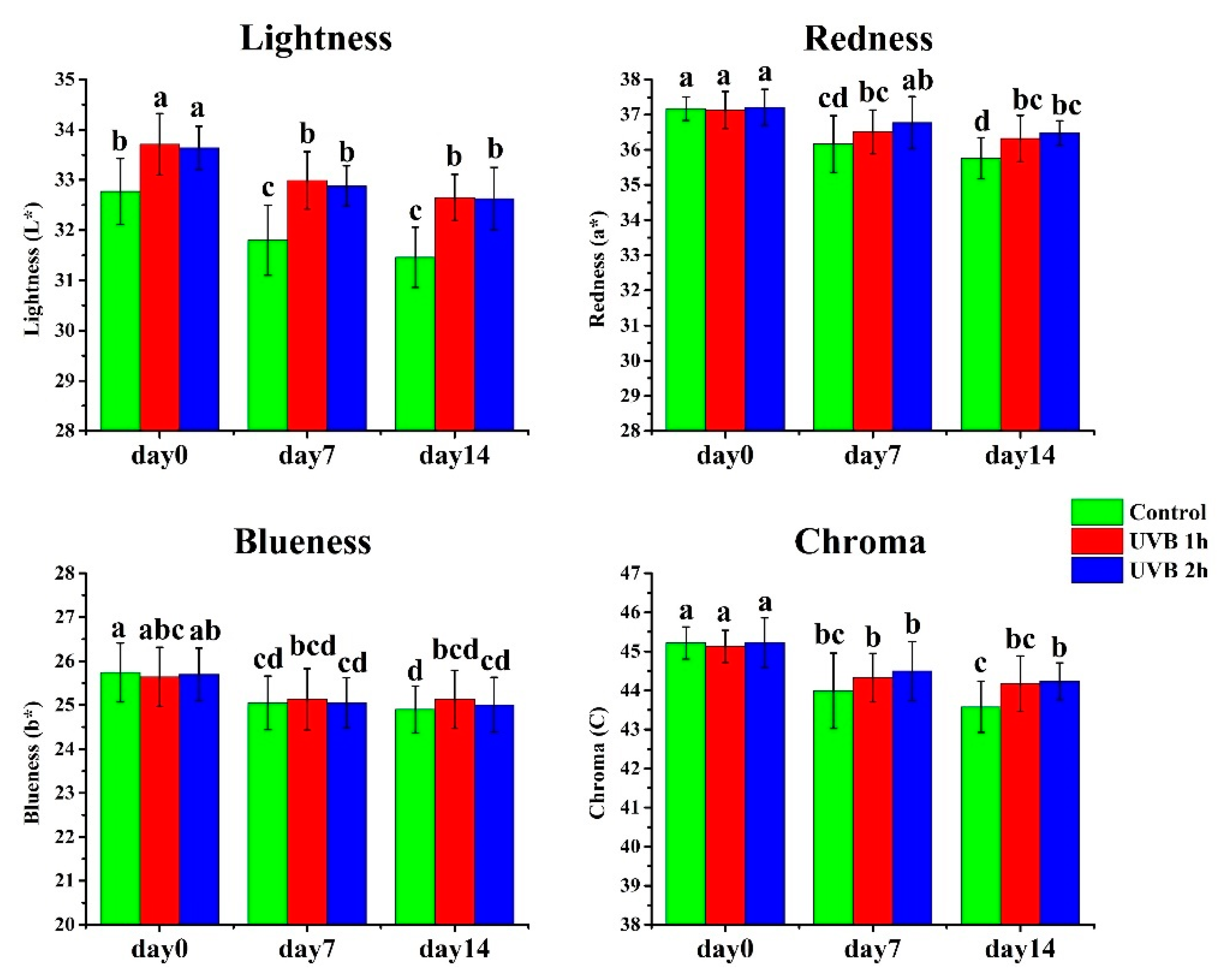
Both UV-B 1 h and UV-B 2 h treated plants exhibited similar growth to control (no treatment). Ripe fruits (red stage) were collected and used for all analyses. The individual fruit weight ranged from 12 to 14 g, and the average single fruit weight for UV-B 1 h, UV-B 2 h and control were 12.9, 13.0 and 12.9 g, respectively. As shown in Figure 1, there were no clear differences in fruit size between UV-B treatments and control, either. As for fruit appearance, at day 0 (harvest day), there were no significant differences in fruit redness (a*), blueness (b*) and chroma (C) (Figure 1 and Figure 2). However, the lightness (L*) value of UV-B-treated fruits was significantly higher than the no treatment control, suggesting that the UV-B-treated fruits maintained a fresher appearance, as darkening/dulling is an indication of senescence in strawberry fruits.
During the storage period, L*, a*, b* and C of strawberry fruit gradually decreased (Figure 2). However, fruits receiving UV-B treatment for one or two hours had higher L* and a* than the control. There was no significant difference between one- and two-hour UV-B treatments. Blueness (b*) and chroma (C) also gradually decreased during storage. However, UV-B treatments did not show a significant influence on them. Therefore, UV-B-treated fruits had greater lightness and a redder surface color than the no-treatment control.
3.2. Effect on Fruit Quality Traits
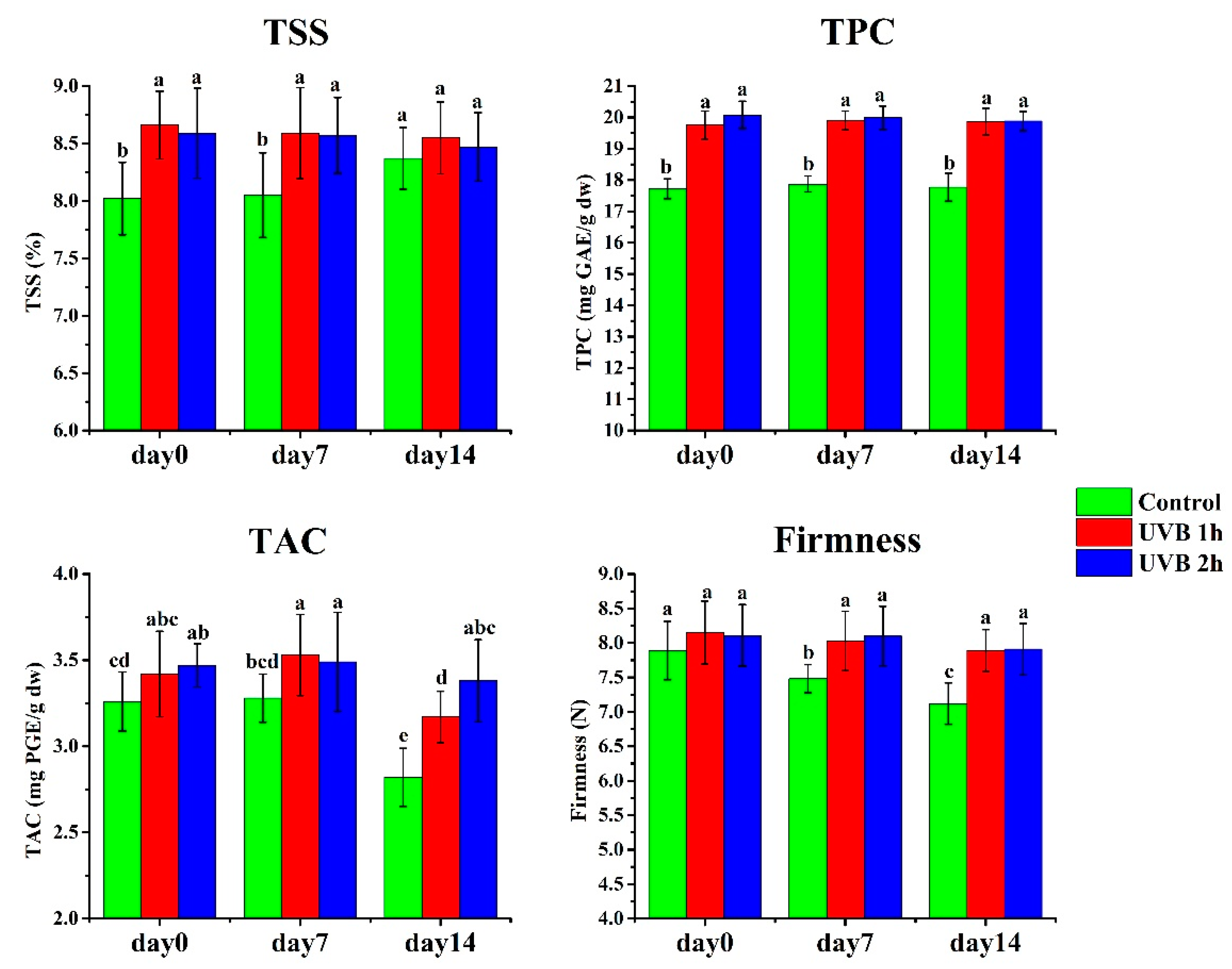
The total soluble solids (TSS), total phenolic content (TPC) and total anthocyanin content (TAC) in UV-B-treated fruits were significantly higher than in untreated fruits on harvest day (day 0) (Figure 3). However, there were no significant differences in fruit firmness between UV-B-treated and untreated fruits on harvest day (day 0).
During the 14-day storage period, TSS of fruits receiving 1 h and 2 h UV-B treatments was significantly higher than the control fruits on day 7, while no significant differences in TSS were observed on day 14. In comparison, TPC and TAC in 1 h and 2 h UV-B-treated fruits were significantly higher than the control fruits on both days 7 and 14. As for fruit firmness, there were no significant differences on day 0, but it steadily declined during storage. However, UV-B-treated fruits were firmer than the untreated fruits all the time. UV-B-treated fruits had higher TSS, TPC and TAC, and maintained greater firmness than control fruits during storage.
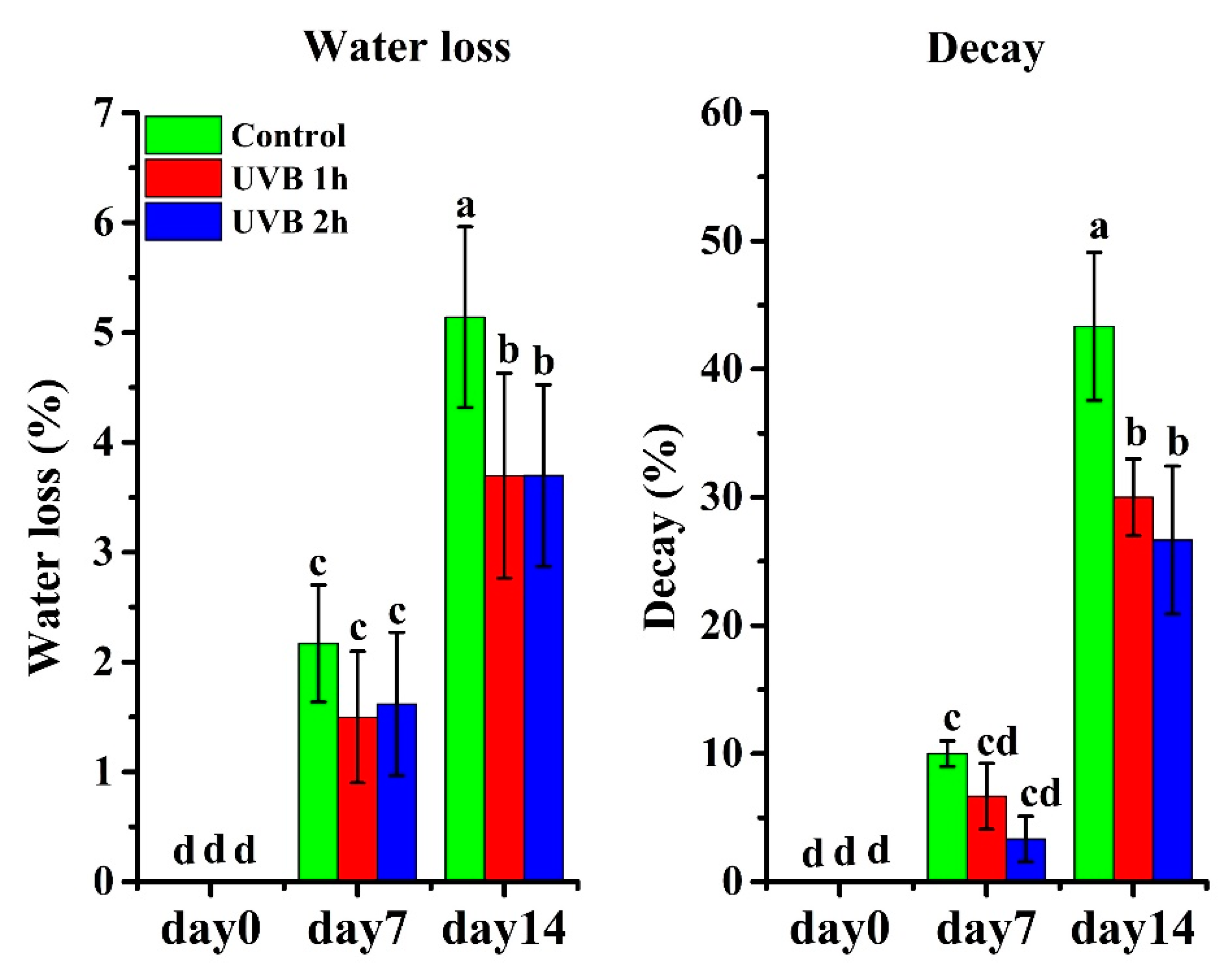
In addition, we observed the occurrence of fruit decay (mainly Botrytis) in control fruit on day 5, with a decay ratio of 10% on day 7, reaching 43.3% on day 14 (Figure 1 and Figure 4). In contrast, the UV-B-treated fruits, especially those treated with 2 h UV-B daily, exhibited a lower decay ratio than the controls. On day 7, one- and two-hour UV-B-treated fruits had decay ratios of 7% and 3%, respectively. On day 14, the decay ratio for one- and two-hour UV-B-treated fruits were 33% and 25%. These results indicate that preharvest UV-B treatments can dramatically reduce fruit decay incidence during storage and extend shelf life.
3.3. Expression of UV-B Responsive Genes in Fruits during Growth Stage in Response to UV-B Treatment
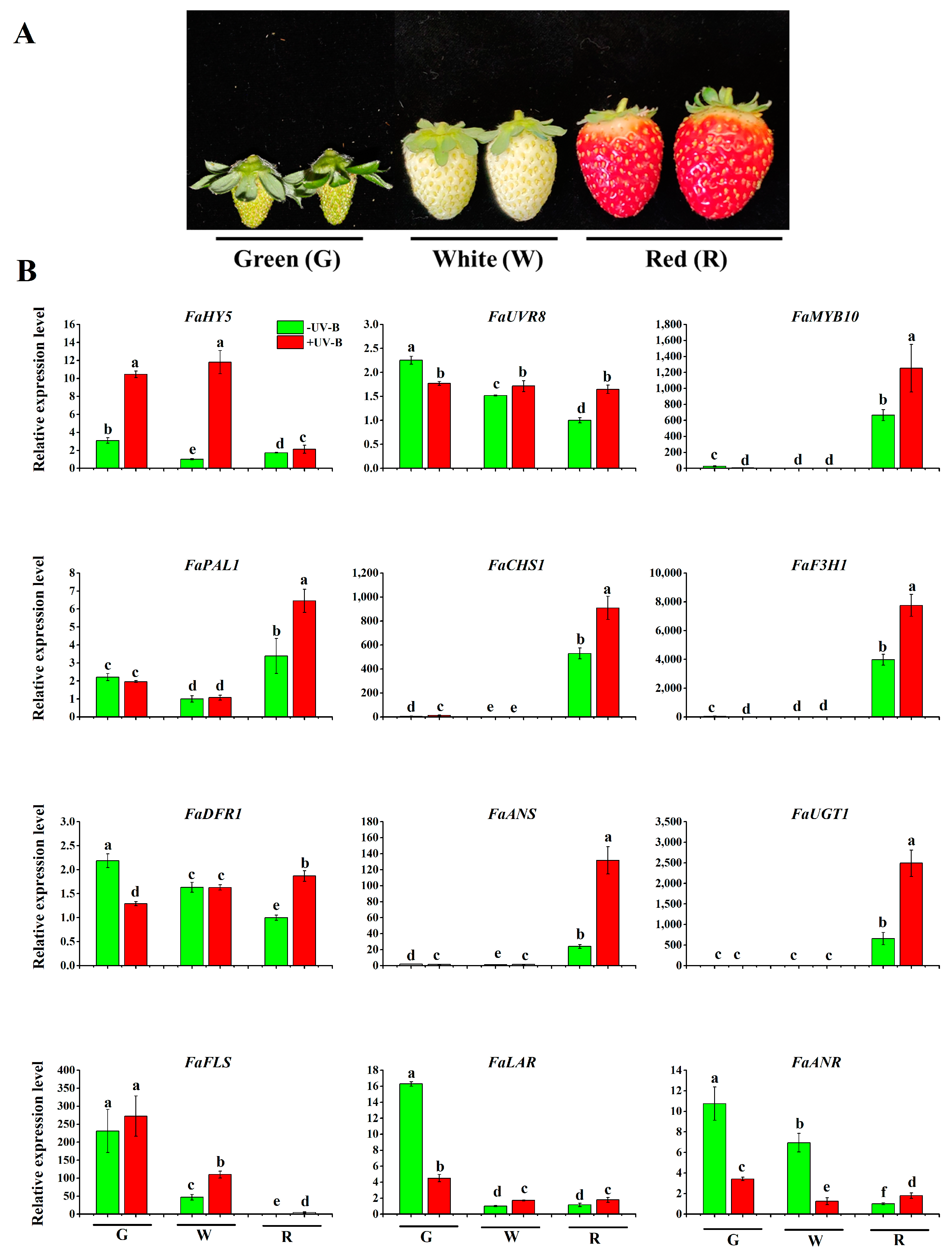
To investigate how fruits respond to UV-B treatments during fruit development, we treated fruits at green, white and red stages with UV-B and examined the expression changes in the genes involved in the UV-B pathway. As shown in Figure 5, the expression of the FaHY5 gene was significantly induced by the UV-B treatment in green- and white-stage berries, while the expression level in red berries was the lowest, and had no obvious response to UV-B treatment. We also examined the expression pattern of FaUVR8. In all berry stages, FaUVR8 was constitutively expressed and showed no response to UV-B treatment. The results were consistent with previous findings in grape, tomato and Arabidopsis showing that UVR8 does not respond to UV-B and FaHY5 shows a significant UV-B regulation [5,19,27].
We further analyzed the expression changes in genes involved in the phenylpropanoid pathway, as well as flavonoids, anthocyanins and proanthocyanidins biosynthesis, because these genes determine fruit color and phenolic content (Figure 5). MYB10 is a key transcription factor in regulating flavonoid and anthocyanin biosynthesis. It is highly expressed in the red stage, and showed a positive response to UV-B treatment. In the general phenylpropanoid pathway, expression of PAL1 was detected in all stages. However, it had the highest expression in the red stage, and UV-B further boosted its expression. As for CHS1 and F3H1, they were dominantly expressed in the red stage. UV-B treatment further stimulated its expression level. In the flavonoid pathway, there are three expression patterns. First, FaLAR and FaANR, which were involved in proanthocyanidin biosynthesis, were highly expressed in the green stage, and hardly detected in the red stage. UV-B treatment reduced their expression. Second, FLS for flavonoid biosynthesis followed a similar expression pattern to that of LAR and ANR. However, UV-B stimulated its expression. Last, ANS and UGT1 for anthocyanin biosynthesis exhibited the highest expression in the red stage, while almost no expression occurred during the green and white stages. UV-B treatment boosted their expression levels by 2–4 folds. These results demonstrate that UV-B can increase anthocyanin biosynthesis in the red stage and flavonoid biosynthesis in the green and white stages.
4. Discussion
Flesh fruits, such as strawberry, are perishable, and their postharvest quality and shelf life are greatly affected by preharvest practice. In this study, we found that the long-term preharvest application of low doses of UV-B improved the fruit quality and shelf life of strawberry fruits.
Plants can perceive UV-B using the UV-B photoreceptor UVR8, which is linked to a specific molecular signaling pathway and leads to UV-B acclimation [5]. In general, the basic leucine zipper (bZIP) transcription factor HY5 plays a significant role in relaying UV-B signaling [28]. The interaction between UVR8 and HY5 is confirmed to play a well-characterized and central role in the UV-B signaling pathway according to Arabidopsis studies [5]. In this study, we observed that FaHY5 was induced significantly in green and white stages. However, the expression of FaHY5 was relatively low in the red stage. This phenomenon has been reported in other crops [29,30,31]. For example, Loyola et al. (2016) reported that VvHY5 was induced in response to low and high UV-B irradiance in green-stage berry skins [30]. VvHYH (HY5 homologue) expression increased in response to UV-B at ripening, and more strikingly by the low radiant exposure treatment before ripening. Li et al. (2021) [31] reported that preharvest long-term UV-B radiation can increase HY5 in the green stage, but not in the mature stage, in blueberry fruit. HY5 may play a key role in responding to UV-B in promoting strawberry fruit development before the red stage.
In the red stage, strawberry fruits undergo color change and the accumulation of anthocyanins, with anthocyanin biosynthesis genes being highly expressed. Those anthocyanin genes are all upregulated by UV-B treatment, while most of them have low expression in the green and white stages. Therefore, HY5 may not be the major component for high anthocyanin accumulation induced by UV-B in strawberry fruits. Li et al. (2021) [31] suggested that the abscisic acid (ABA) pathway could be involved in UV-B perception and anthocyanin accumulation in blueberry fruit, since UV-B irradiation boosted ABA biosynthesis [31,32]. In strawberry, exogenous ABA application can promote MYB expression, accelerate fruit ripening, and increase anthocyanin content [33,34]. Kadomura-Ishikawa et al. (2015) suggested that light and ABA independently regulated FaMYB10. In this study, FaMYB10 and anthocyanin structural genes were upregulated by UV-B treatments. It is still unclear whether UV-B irradiation can increase ABA biosynthesis in strawberry [33].
The abundant phenolic and anthocyanin compounds are regarded as important beneficial antioxidants in strawberries [20]. Postharvest UV-B displayed positive effect on total phenolic and anthocyanin in grapes [9], apples [13] and pears [14]. In this study, the UV-B-irradiated strawberry fruits were found to be redder during the growth stage than control, and UV-B treatment was correlated with high anthocyanin content, too. In addition, preharvest UV-B treatment also enhanced the TCC or sugar content of strawberry fruit, possibly by increasing the sugar transport to fruits. Li et al. (2021) reported that preharvest long-term exposure to UV-B radiation of blueberry plants promoted fruit growth, sugar and anthocyanin contents [31]. In peach, UV-B irradiation enhanced anthocyanin content and sugar accumulation in fruit by increased sorbitol transport to fruit from leaves [35]. UV-B also increased the flavonoid pathway genes and flavonol content of grape berries in a controlled environment [36]. Thus, it could be a general phenomenon that preharvest UV-B promotes the accumulation of sugar and phenolics in fruit. Furthermore, we also observed that preharvest UV-B irradiation has a significant maintenance effect on TAC, TPC and firmness of fruits during storage, as compared to untreated fruits. These events combined, especially high antioxidants, likely contributed to the lower occurrence of fruit decay in strawberry for UV-B-treated fruits. Our study is the first to examine changes in postharvest quality and shelf life in strawberry fruits in response to UV-B treatment during the growth stage.
It should be noted that UV-C (200–280 nm) is often used for preharvest and postharvest treatment of fruits and vegetables, too [1,2,6]. However, UV-C has some extremely inhibitory and damaging effects on plant cells, although it is more effective for postharvest treatment to reduce fungal infections compared to UV-B treatment [3,6]. For example, UV-C inhibited quality formation in strawberry fruit at large green stage during plant growth [34]. Instead, UV-B causes much less damage to plants. We did not observe any inhibitory effect of UV-B treatment on strawberry plant growth and fruit development.
In summary, long-term preharvest irradiation with UV-B (0.7362 J/s m2) had a pronounced effect on improving strawberry fruit color, phenolic and anthocyanin content, and maintaining fruit firmness during postharvest storage, thus extending shelf life. No significant differences were observed between 1 h and 2 h UV-B treatment. Our investigation revealed that that UV-B signal was mainly modulated through the upregulation of HY5 associated with UV-B signal transduction pathway on the early stage of strawberry fruit development. On the red stage, UV-B radiation significantly stimulated the anthocyanin genes independent of HY5. Although most strawberries grow in open fields, and it is costly to add UV-B treatment in fields, the production of strawberries in controlled environments grows rapidly, particularly in urban areas [22]. This makes it feasible to manipulate plant growth conditions, such as through UV light management. Thus, this study provides an effective approach for utilizing preharvest UV-B radiation to improve strawberry fruit appearance and nutritional quality and extend shelf life in controlled-environment agriculture.
Author Contributions
Conceptualization, T.Y.; methodology, X.Z. and F.T.; writing—original draft preparation, X.Z.; writing—review and editing, T.Y. and F.T.; visualization, X.Z., F.T. and T.Y.; supervision, T.Y.; project administration, T.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by USDA-ARS National Program 306 (No. 8042-43000-016-00D) and USDA NE1836.
Data Availability Statement
Not applicable.
Acknowledgments
We thank Jonathan Alcazar for his technical support, and Ellen Turner and Irma Ortiz for editing the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Peng, H.; Pang, Y.; Liao, Q.; Wang, F.; Qian, C. The Effect of Preharvest UV Light Irradiation on Berries Quality: A Review. Horticulturae 2022, 8, 1171. [Google Scholar] [CrossRef]
- Loconsole, D.; Santamaria, P. UV Lighting in Horticulture: A Sustainable Tool for Improving Production Quality and Food Safety. Horticulturae 2021, 7, 9. [Google Scholar] [CrossRef]
- Yao, Y.; Danna, C.H.; Zemp, F.J.; Titov, V.; Ciftci, O.N.; Przybylski, R.; Ausubel, F.M.; Kovalchuk, I. UV-C–irradiated Arabidopsis and tobacco emit volatiles that trigger genomic instability in neighboring plants. Plant Cell 2011, 23, 3842–3852. [Google Scholar] [CrossRef] [PubMed]
- Paul, N.D.; Gwynn-Jones, D. Ecological roles of solar UV radiation: Towards an integrated approach. Trends Ecol. Evol. 2003, 18, 48–55. [Google Scholar] [CrossRef]
- Tilbrook, K.; Arongaus, A.B.; Binkert, M.; Heijde, M.; Yin, R.; Ulm, R. The UVR8 UV-B Photoreceptor: Perception, Signaling and Response. Arab. Book 2013, 11, e0164. [Google Scholar] [CrossRef]
- Urban, L.; Charles, F.; de Miranda, M.R.A.; Aarrouf, J. Understanding the physiological effects of UV-C light and exploiting its agronomic potential before and after harvest. Plant Physiol. Biochem. 2016, 105, 1–11. [Google Scholar] [CrossRef]
- Ruiz, V.E.; Cerioni, L.; Zampini, I.C.; Cuello, S.; Isla, M.I.; Hilal, M.; Rapisarda, V.A. UV-B radiation on lemons enhances antifungal activity of flavedo extracts against Penicillium digitatum. Lwt-Food Sci. Technol. 2017, 85, 96–103. [Google Scholar] [CrossRef]
- Darre, M.; Valerga, L.; Araque, L.C.O.; Lemoine, M.L.; Demkura, P.V.; Vicente, A.R.; Concellon, A. Role of UV-B irradiation dose and intensity on color retention and antioxidant elicitation in broccoli florets (Brassica oleracea var. Italica). Postharvest Biol. Technol. 2017, 128, 76–82. [Google Scholar] [CrossRef]
- Li, X.D.; Wu, B.H.; Wang, L.J.; Zheng, X.B.; Yan, S.T.; Li, S.H. Changes in trans-resveratrol and other phenolic compounds in grape skin and seeds under low temperature storage after post-harvest UV-irradiation. J. Hortic. Sci. Biotech. 2009, 84, 113–118. [Google Scholar] [CrossRef]
- Nguyen, C.T.T.; Lim, S.; Lee, J.G.; Lee, E.J. VcBBX, VcMYB21, and VcR2R3MYB transcription factors are involved in UV B-induced anthocyanin biosynthesis in the peel of harvested blueberry fruit. J. Agric. Food Chem. 2017, 65, 2066–2073. [Google Scholar] [CrossRef]
- Liu, C.H.; Han, X.X.; Cai, L.Y.; Lu, X.Y.; Ying, T.J.; Jiang, Z.H. Postharvest UV-B irradiation maintains sensory qualities and enhances antioxidant capacity in tomato fruit during storage. Postharvest Biol. Technol. 2011, 59, 232–237. [Google Scholar] [CrossRef]
- Josuttis, M.; Dietrich, H.; Treutter, D.; Will, F.; Linnemannstons, L.; Kruger, E. Solar UVB response of bioactives in strawberry (Fragaria x ananassa Duch. L.): A comparison of protected and open-field cultivation. J. Agric. Food Chem. 2010, 58, 12692–12702. [Google Scholar] [CrossRef] [PubMed]
- Marais, E.; Jacobs, G.; Holcroft, D.M. Postharvest irradiation enhances anthocyanin synthesis in apples but not in pears. Hortscience 2001, 36, 738–740. [Google Scholar] [CrossRef]
- Zhang, D.; Yu, B.; Bai, J.H.; Qian, M.J.; Shu, Q.; Su, J.; Teng, Y.W. Effects of high temperatures on UV-B/visible irradiation induced postharvest anthocyanin accumulation in ‘Yunhongli No. 1’ (Pyrus pyrifolia Nakai) pears. Sci. Hortic. 2012, 134, 53–59. [Google Scholar] [CrossRef]
- Wang, C.Y.; Chen, C.T.; Wang, S.Y. Changes of flavonoid content and antioxidant capacity in blueberries after illumination with UV-C. Food Chem. 2009, 117, 426–431. [Google Scholar] [CrossRef]
- Ubi, B.E.; Honda, C.; Bessho, H.; Kondo, S.; Wada, M.; Kobayashi, S.; Moriguchi, T. Expression analysis of anthocyanin biosynthetic genes in apple skin: Effect of UV-B and temperature. Plant Sci. 2006, 170, 571–578. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, G.; Zhang, W. UV-B responsive microRNA genes in Arabidopsis thaliana. Mol. Syst. Biol. 2007, 3, 103. [Google Scholar] [CrossRef]
- Guo, J.; Han, W.; Wang, M.H. Ultraviolet and environmental stresses involved in the induction and regulation of anthocyanin biosynthesis: A review. Afr. J. Biotechnol. 2008, 7, 4966–4972. [Google Scholar]
- Catola, S.; Castagna, A.; Santin, M.; Calvenzani, V.; Petroni, K.; Mazzucato, A.; Ranieri, A. The dominant allele Aft induces a shift from flavonol to anthocyanin production in response to UV-B radiation in tomato fruit. Planta 2017, 246, 263–275. [Google Scholar] [CrossRef]
- Erkan, M.; Wang, S.Y.; Wang, C.Y. Effect of UV treatment on antioxidant capacity, antioxidant enzyme activity and decay in strawberry fruit. Postharvest Biol. Technol. 2008, 48, 163–171. [Google Scholar] [CrossRef]
- Ross, J.A.; Kasum, C.M. Dietary flavonoids: Bioavailability, metabolic effects, and safety. Annu. Rev. Nutr. 2002, 22, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Samtani, J.B.; Rom, C.R.; Friedrich, H.; Fennimore, S.A.; Finn, C.E.; Petran, A.; Wallace, R.W.; Pritts, M.P.; Fernandez, G.; Chase, C.A.; et al. The Status and Future of the Strawberry Industry in the United States. Horttechnology 2019, 29, 11–24. [Google Scholar] [CrossRef]
- Dong, W.; Lu, Y.; Yang, T.; Trouth, F.; Lewers, K.S.; Daughtry, C.S.; Cheng, Z.M. Effect of genotype and plastic film type on strawberry fruit quality and post-harvest shelf life. Int. J. Fruit Sci. 2020, 20, 750–767. [Google Scholar] [CrossRef]
- Lester, G.E.; Lewers, K.S.; Medina, M.B.; Saftner, R.A. Comparative analysis of strawberry total phenolics via Fast Blue BB vs. Folin-Ciocalteu: Assay interference by ascorbic acid. J. Food Compos. Anal. 2012, 27, 102–107. [Google Scholar] [CrossRef]
- Sanchez-Moreno, C.; Cao, G.H.; Ou, B.X.; Prior, R.L. Anthocyanin and proanthocyanidin content in selected white and red wines. Oxygen radical absorbance capacity comparison with nontraditional wines obtained from highbush blueberry. J. Agric. Food Chem. 2003, 51, 4889–4896. [Google Scholar] [CrossRef]
- Xu, W.; Peng, H.; Yang, T.; Whitaker, B.; Huang, L.; Sun, J.; Chen, P. Effect of calcium on strawberry fruit flavonoid pathway gene expression and anthocyanin accumulation. Plant Physiol. Biochem. 2014, 82, 289–298. [Google Scholar] [CrossRef]
- Liu, L.L.; Gregan, S.; Winefield, C.; Jordan, B. From UVR8 to flavonol synthase: UV-B-induced gene expression in Sauvignon blanc grape berry. Plant Cell Environ. 2015, 38, 905–919. [Google Scholar] [CrossRef]
- An, J.P.; Qu, F.J.; Yao, J.F.; Wang, X.N.; You, C.X.; Wang, X.F.; Hao, Y.J. The bZIP transcription factor MdHY5 regulates anthocyanin accumulation and nitrate assimilation in apple. Hortic. Res. 2017, 4, 17056. [Google Scholar] [CrossRef]
- Carbonell-Bejerano, P.; Diago, M.P.; Martinez-Abaigar, J.; Martinez-Zapater, J.M.; Tardaguila, J.; Nunez-Olivera, E. Solar ultraviolet radiation is necessary to enhance grapevine fruit ripening transcriptional and phenolic responses. BMC Plant Biol. 2014, 14, 183. [Google Scholar] [CrossRef]
- Loyola, R.; Herrera, D.; Mas, A.; Wong, D.C.J.; Holl, J.; Cavallini, E.; Amato, A.; Azuma, A.; Ziegler, T.; Aquea, F.; et al. The photomorphogenic factors UV-B RECEPTOR 1, ELONGATED HYPOCOTYL 5, and HY5 HOMOLOGUE are part of the UV-B signalling pathway in grapevine and mediate flavonol accumulation in response to the environment. J. Exp. Bot. 2016, 67, 5429–5445. [Google Scholar] [CrossRef]
- Li, T.S.; Yamane, H.; Tao, R. Preharvest long-term exposure to UV-B radiation promotes fruit ripening and modifies stage-specific anthocyanin metabolism in highbush blueberry. Hortic. Res. 2021, 8, 67. [Google Scholar] [CrossRef] [PubMed]
- Tossi, V.; Lamattina, L.; Cassia, R. An increase in the concentration of abscisic acid is critical for nitric oxide-mediated plant adaptive responses to UV-B irradiation. New Phytol. 2009, 181, 871–879. [Google Scholar] [CrossRef] [PubMed]
- Kadomura-Ishikawa, Y.; Miyawaki, K.; Takahashi, A.; Masuda, T.; Noji, S. Light and abscisic acid independently regulated FaMYB10 in Fragaria x ananassa fruit. Planta 2015, 241, 953–965. [Google Scholar] [CrossRef]
- Li, D.D.; Luo, Z.S.; Mou, W.S.; Wang, Y.S.; Ying, T.J.; Mao, L.C. ABA and UV-C effects on quality, antioxidant capacity and anthocyanin contents of strawberry fruit (Fragaria ananassa Duch.). Postharvest Biol. Technol. 2014, 90, 56–62. [Google Scholar] [CrossRef]
- Wang, X.; Fu, X.; Chen, M.; Huan, L.; Liu, W.; Qi, Y.; Gao, Y.; Xiao, W.; Chen, X.; Li, L.; et al. Ultraviolet B irradiation influences the fruit quality and sucrose metabolism of peach (Prunus persica L.). Environ. Exp. Bot. 2018, 153, 286–301. [Google Scholar] [CrossRef]
- Liu, L.; Gregan, S.M.; Winefield, C.; Jordan, B. Comparisons of controlled environment and vineyard experiments in Sauvignon blanc grapes reveal similar UV-B signal transduction pathways for flavonol biosynthesis. Plant Sci. 2018, 276, 44–53. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effect of preharvest UV-B treatment on fruit appearance and shelf life. Red fruits were harvested from plants treated with different durations of UV-B (0.7362 J/s m2) treatment and control (no treatment). Fruits were stored at 5 °C and 75% relative humidity. Photos were taken on day 0 (harvest day) and days 7 and 14 after harvest.
Figure 1.
Effect of preharvest UV-B treatment on fruit appearance and shelf life. Red fruits were harvested from plants treated with different durations of UV-B (0.7362 J/s m2) treatment and control (no treatment). Fruits were stored at 5 °C and 75% relative humidity. Photos were taken on day 0 (harvest day) and days 7 and 14 after harvest.

Figure 2.
Effects of UV-B treatment on surface color of strawberry fruits. The data are based on ten biological replicates. * p < 0.05.
Figure 2.
Effects of UV-B treatment on surface color of strawberry fruits. The data are based on ten biological replicates. * p < 0.05.

Figure 3.
Effects of UV-B treatment on strawberry fruit quality. TSS, total soluble solids (Brix%); TPC, total phenolic content (mg GAE/g dry weight)), and TAC, total anthocyanin content (mg PGE/g dry weight). The data are based on ten biological replicates.
Figure 3.
Effects of UV-B treatment on strawberry fruit quality. TSS, total soluble solids (Brix%); TPC, total phenolic content (mg GAE/g dry weight)), and TAC, total anthocyanin content (mg PGE/g dry weight). The data are based on ten biological replicates.

Figure 4.
Effects of UV-B treatment on water loss and decay incidence of strawberry fruits. The data are based on ten biological replicates.
Figure 4.
Effects of UV-B treatment on water loss and decay incidence of strawberry fruits. The data are based on ten biological replicates.

Figure 5.
Expression patterns of UV-B signaling pathway and anthocyanin biosynthetic genes in strawberry fruits after UV-B 1 h treatment. (A) Fruit images at different stages. (B) Comparison of gene expression levels. G: fruit at the green stage; W: fruit at the white stage; R: fruit at the ripe stage; RNA levels were quantified and normalized to the level of FaACTIN. Values are means ± SD of three replicates. The data are based on three biological replicates.
Figure 5.
Expression patterns of UV-B signaling pathway and anthocyanin biosynthetic genes in strawberry fruits after UV-B 1 h treatment. (A) Fruit images at different stages. (B) Comparison of gene expression levels. G: fruit at the green stage; W: fruit at the white stage; R: fruit at the ripe stage; RNA levels were quantified and normalized to the level of FaACTIN. Values are means ± SD of three replicates. The data are based on three biological replicates.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
List of primers used for gene expression analysis.
| Gene Name | Gene ID | Forward Sequence (5′-3′) | Reverse Sequence (3′-5′) |
|---|---|---|---|
| FaACTIN | JN616288 | TCACCACCACTGCTGAACGGGA | TGCAAGCTTCTCCTTCATGTCACGG |
| FaHY5 | KP984791 | TGTTGCAAGACCAAGCCACGAGC | AGACCTCTCACTGCTGGAAGGCA |
| FaUVR8 | KU647690 | AGGGAAGATTTGGGTGTCACCGTCA | GCCGTATGCGTTTCCCGTCGGTTT |
| FaMYB10 | EU155162.1 | GACGGCTTCATACGCAAAGA | TCTGTGGTGGTTCTGTTGGT |
| FaPAL1 | KX450226 | GCCAAGGAGGAGGACTACTACATGACC | TCCGCCGCCAAGTTCCAGTTCA |
| FaCHS1 | AY997297 | ACGGCCCAAACTATCCTTCCCGA | GCCCAACTTCACGAAGATGCCCGT |
| FaF3H1 | AY691919 | CGGTGCAGGATTGGCGCGAGA | CTGCTGTGGTTTGAGTTCACCAC |
| FaDFR1 | KC894047 | GCCGGAAAGTAAAGATGACTGG | GCTTCATTTCCAGTGAGTGGTG |
| FaANS | AY695817 | TCGAGAGTTTGGCCAGCAGCGGGAT | TGAGGCCCTTCGGTGCTCTTCTCGT |
| FaUGT1 | LC312710 | GGAATCATCTTCGGAAACTTGG | TGCACTTGCTGGTGGTTCTAGT |
| FaFLS | DQ087252 | CCCGGCGGAGTACATTAGGTCGGA | ACGGTGGTGATTCCCGGCTGCT |
| FaLAR | DQ087253.1 | GCAATATCACGGCTACTTGTGC | GAAAATGCAGCTTCCTTGCTTT |
| FaANR | DQ664192.1 | CCGATGAAAATGATTGGTCTGA | GTCTGGAGTGAGAGAAGCACCA |
HY5: ELONGATED HYPOCOTYL 5; UVR8: ultraviolet-B receptor 8; PAL1: Phenylalanine ammonia lyase 1; CHS1: Chalcone synthase 1; F3H1: flavanone 3-hydroxylase 1; DFR1: Dihydroflavonol 4-reductase 1; ANS: Anthocyanidin synthase; UGT1: UDP-glucose glucosyltransferase 1; FLS: Flavonol synthase; LAR: leucoanthocyanidin reductase; ANR1: Anthocyanidin reductase.
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhu, X.; Trouth, F.; Yang, T. Preharvest UV-B Treatment Improves Strawberry Quality and Extends Shelf Life. Horticulturae 2023, 9, 211. https://doi.org/10.3390/horticulturae9020211
AMA Style
Zhu X, Trouth F, Yang T. Preharvest UV-B Treatment Improves Strawberry Quality and Extends Shelf Life. Horticulturae. 2023; 9(2):211. https://doi.org/10.3390/horticulturae9020211
Chicago/Turabian StyleZhu, Xudong, Frances Trouth, and Tianbao Yang. 2023. "Preharvest UV-B Treatment Improves Strawberry Quality and Extends Shelf Life" Horticulturae 9, no. 2: 211. https://doi.org/10.3390/horticulturae9020211
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.